
YTHDF2 as a Mediator in BDNF-Induced Proliferation of Porcine Follicular Granulosa Cells

Abstract
:1. Introduction
2. Results
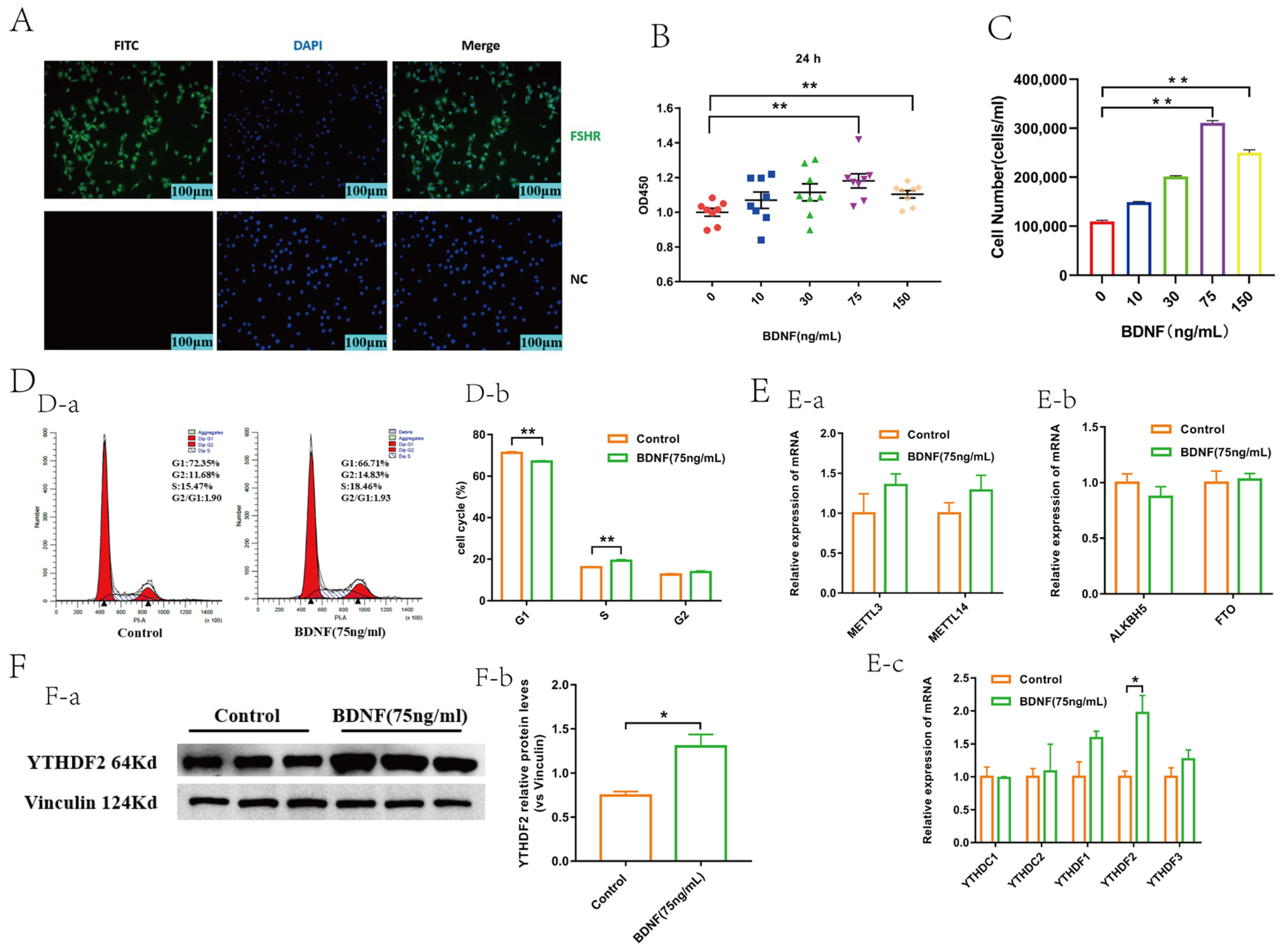
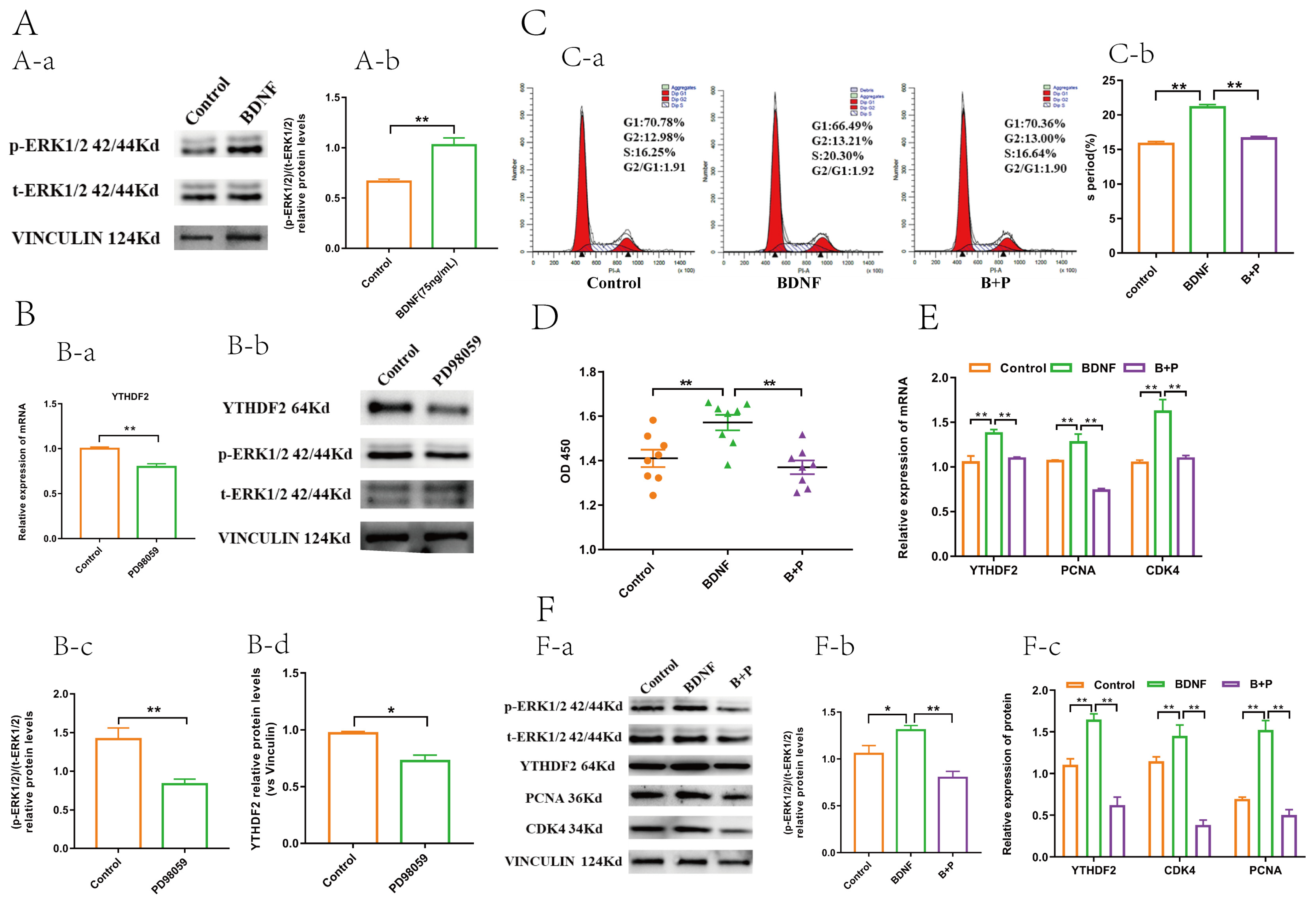
2.1. BDNF Promoted the Proliferation and Upregulated the Expression of YTHDF2 in GCs
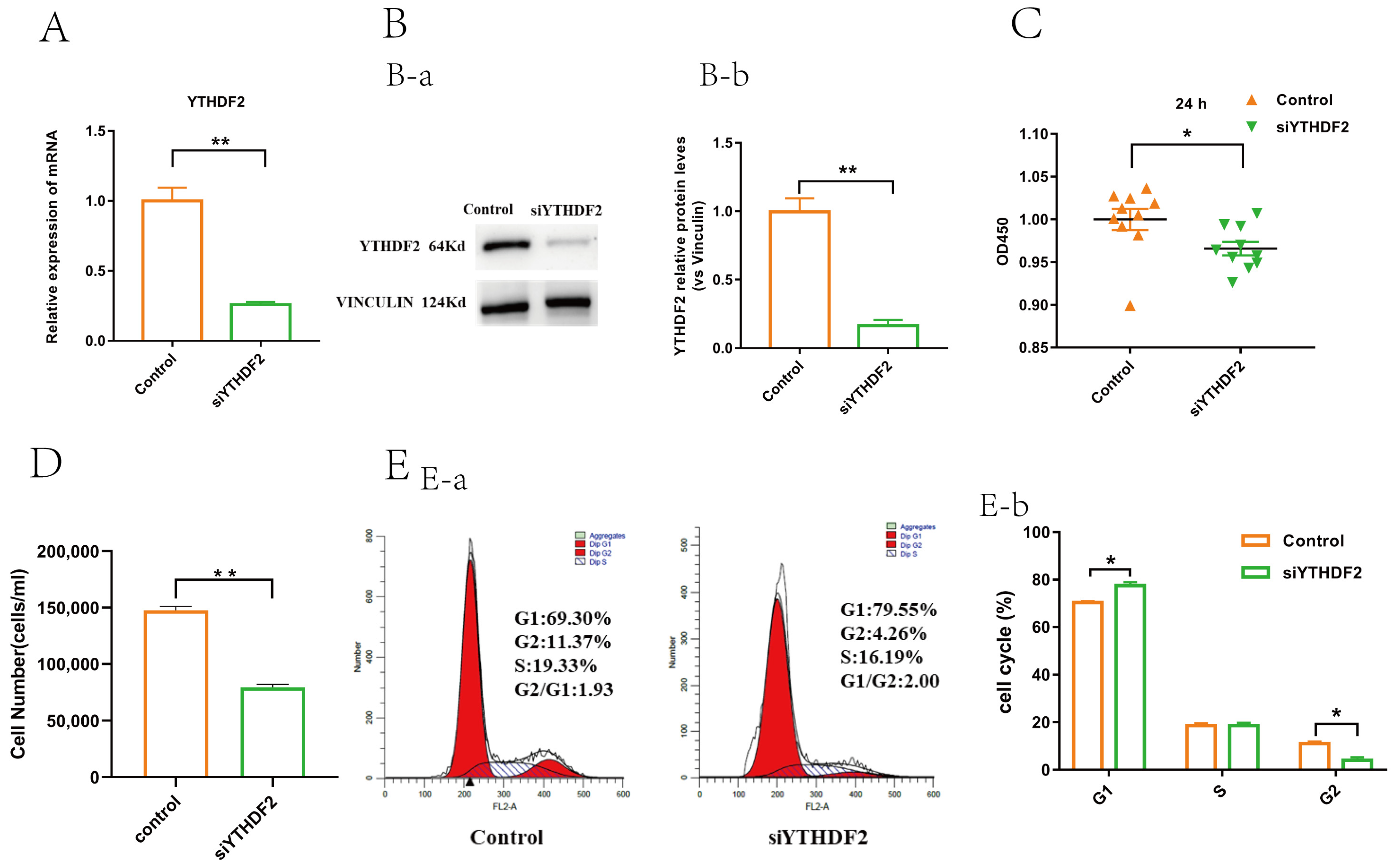
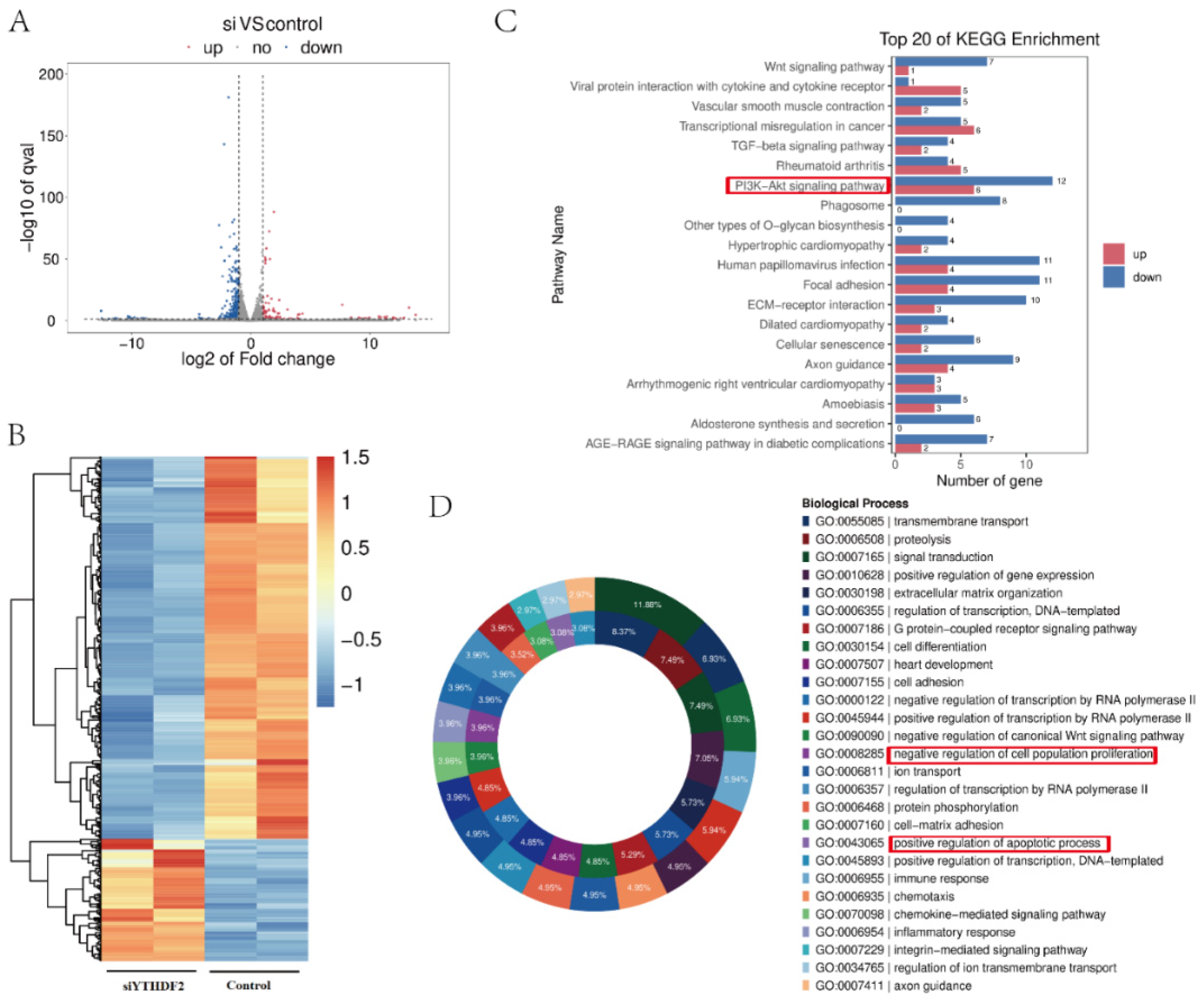
2.2. YTHDF2 Knockdown Impacts Proliferation and Gene Expression in GCs
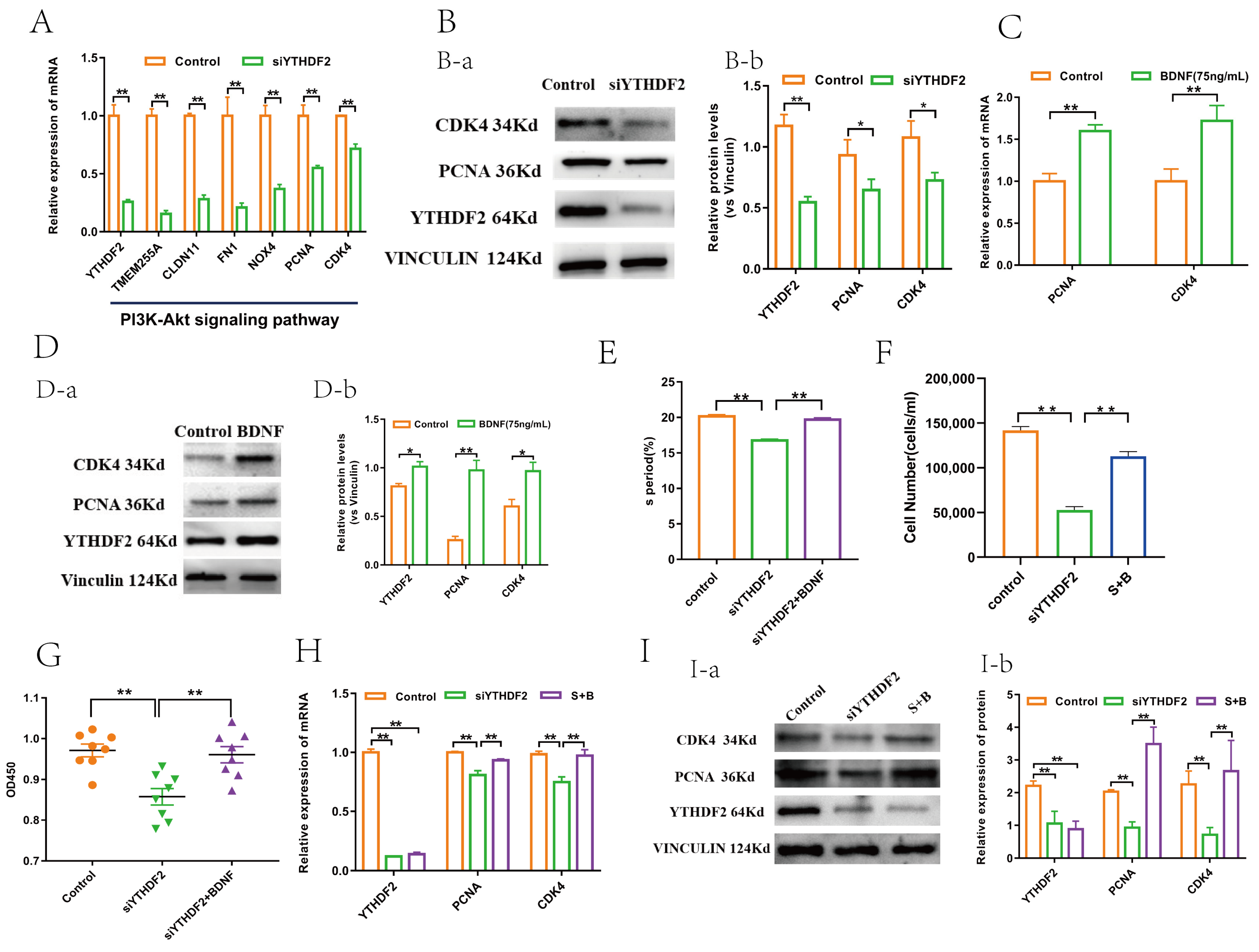
2.3. YTHDF2 Facilitates GC Proliferation by Upregulating CDK4 and PCNA
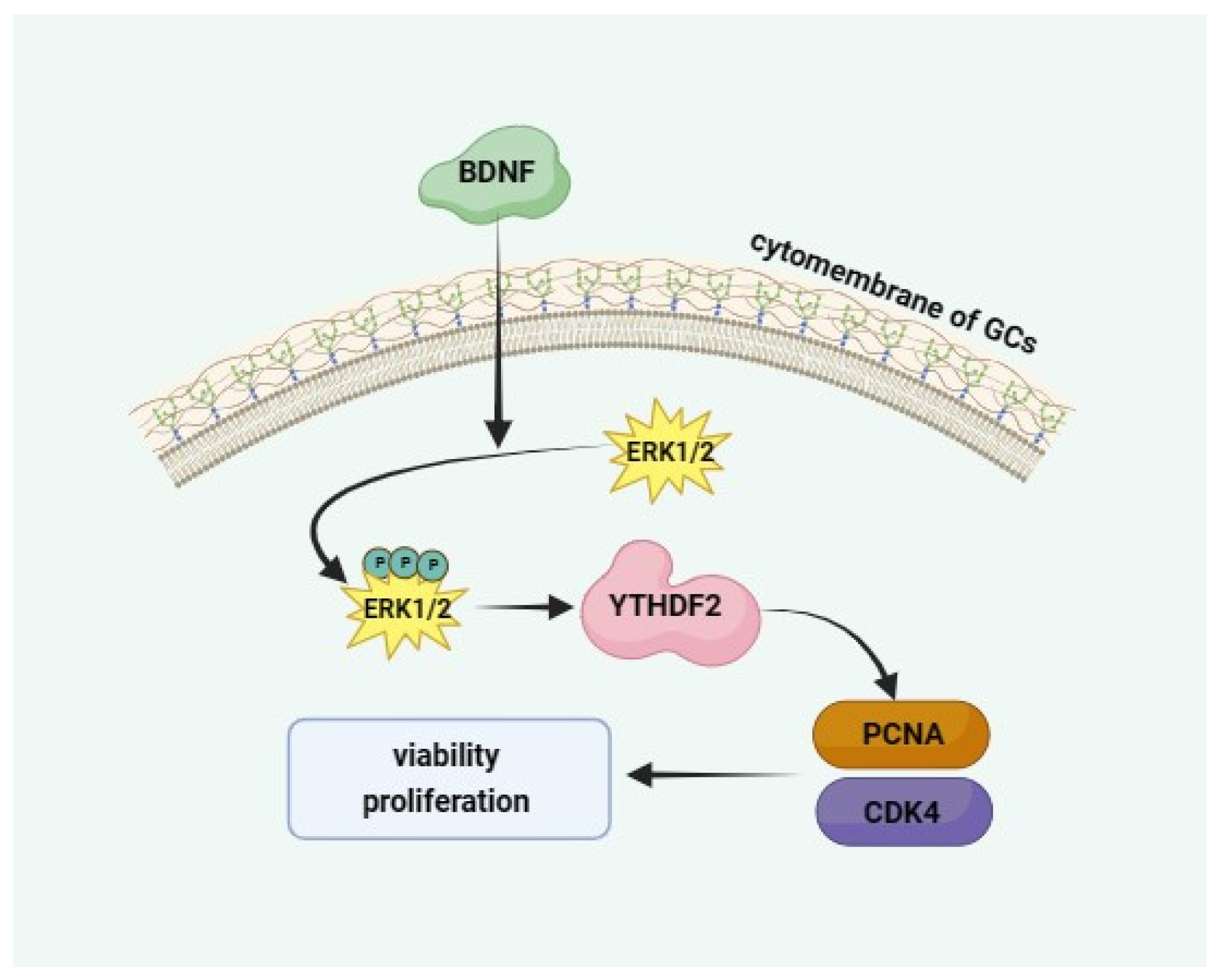
2.4. BDNF Regulates YTHDF2 through the ERK1/2 Signalling Pathway
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Treatments
4.2. Cellular Immunofluorescence
4.3. Cell Viability Assay and Cell Counting Assay
4.4. Flow Cytometry
4.5. Transfection with Small Interfering RNA
4.6. RNA Extraction, RT-PCR, and RT-qPCR Analysis
4.7. Western Blot Analysis
4.8. Sequencing and Data Processing
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| GCs | Granulosa cells |
| BDNF | Brain-derived neurotrophic factor |
| YTHDF2 | YTH domain-containing family member 2 |
| CDK4 | Cyclin-dependent Kinase 4 |
| PCNA | Proliferating Cell Nuclear Antigen |
| ERK1/2 | Extracellular regulated protein kinases 1/2 |
| siYTHDF2 | siRNA targeting YTHDF2 |
| FSH | Follicle-stimulating hormone |
| hCG | Human chorionic gonadotropin |
| NGF | Nerve growth factor |
| NT-3 | Neurotrophin 3 |
| NT-4/5 | Neurotrophin 4/5 |
| TrkB | Tyrosine kinase receptor protein B |
| m6A | N6-methyladenosine |
References
- Chow, R.; Wessels, J.M.; Foster, W.G. Brain-derived neurotrophic factor (BDNF) expression and function in the mammalian reproductive Tract. Hum. Reprod. Update 2020, 26, 545–564. [Google Scholar] [CrossRef]
- Dissen, G.A.; Romero, C.; Hirshfield, A.N.; Ojeda, S.R. Nerve Growth Factor Is Required for Early Follicular Development in the Mammalian Ovary. Endocrinology 2001, 142, 2078–2086. [Google Scholar] [CrossRef]
- Oron, G.; Ao, A.; Friedman, O.; Fisch, B.; Zhang, X.Y.; Ben-Haroush, A.; Peled, Y.; Abir, R. Expression of neurotrophin 3 and its tropomyosin-related kinase receptor C in human preantral follicles. Fertil. Steril. 2011, 95, 2056–2062. [Google Scholar] [CrossRef]
- Nilsson, E.; Dole, G.; Skinner, M.K. Neurotrophin NT3 promotes ovarian primordial to primary follicle transition. Reproduction 2009, 138, 697–707. [Google Scholar] [CrossRef]
- Farhi, J.; Fisch, B.; Garor, R.; Peled, Y.; Pinkas, H.; Abir, R. Neurotrophin 4 enhances in vitro follicular assembly in human fetal ovaries. Fertil. Steril. 2011, 95, 1267–1271. [Google Scholar] [CrossRef]
- Deinhardt, K.; Chao, M.V. Shaping neurons: Long and short range effects of mature and proBDNF signalling upon neuronal structure. Neuropharmacology 2014, 76, 603–609. [Google Scholar] [CrossRef]
- Li, C.; Chen, C.; Chen, L.; Chen, S.; Li, H.; Zhao, Y.; Rao, J.; Zhou, X. BDNF-induced expansion of cumulus-oocyte complexes in pigs was mediated by microRNA-205. Theriogenology 2016, 85, 1476–1482. [Google Scholar] [CrossRef]
- Zhao, P.; Qiao, J.; Huang, S.; Zhang, Y.; Liu, S.; Yan, L.-Y.; Hsueh, A.J.; Duan, E.-K. Gonadotrophin-induced paracrine regulation of human oocyte maturation by BDNF and GDNF secreted by granulosa cells. Hum. Reprod. 2011, 26, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wang, F.; Liu, Z.; Zhao, Y.; Jiang, Y.; Chen, L.; Li, C.; Zhou, X. Brain-derived neurotrophic factor promotes proliferation and progesterone synthesis in bovine granulosa cells. J. Cell. Physiol. 2019, 234, 8776–8787. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Du, F.; Liu, X.; Ruan, Q.; Wu, Z.; Lei, C.; Deng, Y.; Luo, C.; Jiang, J.; Shi, D.; et al. Brain-derived neurotrophic factor (BDNF) is expressed in buffalo (Bubalus bubalis) ovarian follicles and promotes oocyte maturation and early embryonic development. Theriogenology 2019, 130, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Fang, Y.; Xu, Y.; Sun, H. Role of m6A modification in female infertility and reproductive system diseases. Int. J. Biol. Sci. 2022, 18, 3592–3604. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Wu, R.; Ming, L. The role of m6A RNA methylation in cancer. Biomed. Pharmacother. 2019, 112, 108613. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Ruan, Z.; Feng, Y.; Wang, Y.; Zhang, J.; Lu, C.; Shi, D.; Lu, F. METTL3-mediated m6A methylation regulates granulosa cells autophagy during follicular atresia in pig ovaries. Theriogenology 2023, 201, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Mu, H.; Zhang, T.; Yang, Y.; Zhang, D.; Gao, J.; Li, J.; Yue, L.; Gao, D.; Shi, B.; Han, Y.; et al. METTL3-mediated mRNA N6-methyladenosine is required for oocyte and follicle development in mice. Cell Death Dis. 2021, 12, 989. [Google Scholar] [CrossRef]
- Jin, D.; Guo, J.W.; Wu, Y.; Yang, L.J.; Wang, X.H.; Du, J.; Dai, J.J.; Chen, W.W.; Gong, K.K.; Miao, S.; et al. m6A demethylase ALKBH5 inhibits tumor growth and metastasis by reducing YTHDFs-mediated YAP expression and inhibiting miR-107/LATS2-mediated YAP activity in NSCLC. Mol. Cancer 2020, 19, 40. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Han, X.; Li, W.; Wang, N.; Yao, L.; Zhao, Y.; Zhang, L. N6-methyladenosine Demethylase FTO Induces the Dysfunctions of Ovarian Granulosa Cells by Upregulating Flotillin 2. Reprod. Sci. 2022, 29, 1305–1315. [Google Scholar] [CrossRef] [PubMed]
- Mapperley, C.; van de Lagemaat, L.N.; Lawson, H.; Tavosanis, A.; Paris, J.; Campos, J.; Wotherspoon, D.; Durko, J.; Sarapuu, A.; Choe, J.; et al. The mRNA m6A reader YTHDF2 suppresses proinflammatory pathways and sustains hematopoietic stem cell function. J. Exp. Med. 2021, 218, e20200829. [Google Scholar] [CrossRef]
- Kasowitz, S.D.; Ma, J.; Anderson, S.J.; Leu, N.A.; Xu, Y.; Gregory, B.D.; Schultz, R.M.; Wang, P.J. Nuclear m6A reader YTHDC1 regulates alternative polyadenylation and splicing during mouse oocyte development. PLoS Genet. 2018, 14, e1007412. [Google Scholar] [CrossRef]
- Zeng, M.; Dai, X.; Liang, Z.; Sun, R.; Huang, S.; Luo, L.; Li, Z. Critical roles of mRNA m6A modification and YTHDC2 expression for meiotic initiation and progression in female germ cells. Gene 2020, 753, 144810. [Google Scholar] [CrossRef]
- Zhang, S.; Deng, W.; Liu, Q.; Wang, P.; Yang, W.; Ni, W. Altered m6A modification is involved in up-regulated expression of FOXO3 in luteinized granulosa cells of non-obese polycystic ovary syndrome patients. J. Cell. Mol. Med. 2020, 24, 11874–11882. [Google Scholar] [CrossRef]
- Ivanova, I.; Much, C.; Di Giacomo, M.; Azzi, C.; Morgan, M.; Moreira, P.N.; Monahan, J.; Carrieri, C.; Enright, A.J.; O’carroll, D. The RNA m 6 A Reader YTHDF2 Is Essential for the Post-transcriptional Regulation of the Maternal Transcriptome and Oocyte Competence. Mol. Cell 2017, 67, 1059–1067.e4. [Google Scholar] [CrossRef]
- Wang, J.-Y.; Lu, A.-Q. The biological function of m6A reader YTHDF2 and its role in human disease. Cancer Cell Int. 2021, 21, 109. [Google Scholar] [CrossRef]
- Childs, A.J.; Bayne, R.A.; Murray, A.A.; Da Silva, S.J.M.; Collins, C.S.; Spears, N.; Anderson, R.A. Differential expression and regulation by activin of the neurotrophins BDNF and NT4 during human and mouse ovarian development. Dev. Dyn. 2010, 239, 1211–1219. [Google Scholar] [CrossRef]
- Peng, J.; An, X.; Fang, F.; Gao, K.; Xin, H.; Han, P.; Bao, L.; Ma, H.; Cao, B. MicroRNA-10b suppresses goat granulosa cell proliferation by targeting brain-derived neurotropic factor. Domest. Anim. Endocrinol. 2016, 54, 60–67. [Google Scholar] [CrossRef]
- Gao, S.; Chen, S.; Chen, L.; Zhao, Y.; Sun, L.; Cao, M.; Huang, Y.; Niu, Q.; Wang, F.; Yuan, C.; et al. Brain-derived neurotrophic factor: A steroidogenic regulator of Leydig cells. J. Cell. Physiol. 2019, 234, 14058–14067. [Google Scholar] [CrossRef]
- Cao, Z.; Zhang, D.; Wang, Y.; Tong, X.; Avalos, L.F.C.; Khan, I.M.; Gao, D.; Xu, T.; Zhang, L.; Knott, J.G.; et al. Identification and functional annotation of m6A methylation modification in granulosa cells during antral follicle development in pigs. Anim. Reprod. Sci. 2020, 219, 106510. [Google Scholar] [CrossRef] [PubMed]
- Fei, Q.; Zou, Z.; Roundtree, I.A.; Sun, H.-L.; He, C. YTHDF2 promotes mitotic entry and is regulated by cell cycle mediators. PLoS Biol. 2020, 18, e3000664. [Google Scholar] [CrossRef]
- Hua, Z.; Wei, R.; Guo, M.; Lin, Z.; Yu, X.; Li, X.; Gu, C.; Yang, Y. YTHDF2 promotes multiple myeloma cell proliferation via STAT5A/MAP2K2/p-ERK axis. Oncogene 2022, 41, 1482–1491. [Google Scholar] [CrossRef]
- Zhong, L.; Liao, D.; Zhang, M.; Zeng, C.; Li, X.; Zhang, R.; Ma, H.; Kang, T. YTHDF2 suppresses cell proliferation and growth via destabilizing the EGFR mRNA in hepatocellular carcinoma. Cancer Lett. 2019, 442, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Lim, W.; Bae, H.; Bazer, F.W.; Song, G. Brain-derived neurotrophic factor improves proliferation of endometrial epithelial cells by inhibition of endoplasmic reticulum stress during early pregnancy. J. Cell. Physiol. 2017, 232, 3641–3651. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, I.; Thompson, W.E.; Welch, C.; Thomas, K.; Matthews, R. Prohibitin (PHB) inhibits apoptosis in rat granulosa cells (GCs) through the extracellular signal-regulated kinase 1/2 (ERK1/2) and the Bcl family of proteins. Apoptosis 2013, 18, 1513–1525. [Google Scholar] [CrossRef] [PubMed]
- Fang, R.; Chen, X.; Zhang, S.; Shi, H.; Ye, Y.; Shi, H.; Zou, Z.; Li, P.; Guo, Q.; Ma, L.; et al. EGFR/SRC/ERK-stabilized YTHDF2 promotes cholesterol dysregulation and invasive growth of glioblastoma. Nat. Commun. 2021, 12, 177. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequences | Products Sizes, bp | Melting Temperature, °C |
|---|---|---|---|
| GAPDH | F-5′ TCGGAGTGAACGGATTTGGC3′ | 147 | 83.6 |
| R-5′ TGCCGTGGGTGGAATCATAC3′ | |||
| YTHDF2 | F-5′ CAGGCAAGGCCCAATAATGC3′ | 186 | 86.1 |
| R-5′ TCTCCGTTGCTCAGTTGTCC3′ | |||
| CDK4 | F-5′ ATGGGACCGTGTACAAAGCA3′ | 165 | 81.2 |
| R-5′ CATCCATCAGCCGGACAACA3′ | |||
| PCNA | F-5′ ATGCAGACACCTTGGCACTA3′ | 153 | 79.9 |
| R-5′ ACGTGCAAATTCACCAGAAGG3′ | |||
| TMEM255A | F-5′ TCCTCAAGTGGCCTCCTACA3′ | 190 | 88.2 |
| R-5′ TGGAGAGTATCGGGGTGGAG3′ | |||
| CLDN11 | F-5′ GTGACCTGCGGCTACACTAT3′ | 182 | 91.1 |
| R-5′ TCGGCAAGCCTGAACATAGC3′ | |||
| FN1 | F-5′ GCACCATCCAACTTGCGTTT3′ | 179 | 88.1 |
| R-5′ TGTACTCGGTTGCTGGTTCC3′ | |||
| NOX4 | F-5′ TCCTGGCTTACCTTCGAGGA3′ | 146 | 85.4 |
| R-5′ TTCACGGAGAAGTTGAGGGC3′ |
| Antibodies Name | Diluted Multiples | Accession Number | Reagent Company |
|---|---|---|---|
| Rabbit anti-vinculin polyclonal antibody | 1:750 | BS62273 | Bioworld |
| Rabbit anti-YTHDF2 monoclonal antibody | 1:750 | ab220163 | Abcam |
| Rabbit anti-CDK4 polyclonal antibody | 1:750 | ab95255 | Abcam |
| Mouse anti-PCNA monoclonal antibody | 1:750 | MB66871 | Bioworld |
| Rabbit anti-ERK1/2 polyclonal antibody | 1:750 | BS90472 | Bioworld |
| Rabbit anti-phospho-ERK1/2 polyclonal antibody | 1:750 | BS4621P | Bioworld |
| Goat anti-rabbit IgG antibody | 1:8000 | BS13278 | Bioworld |
| Goat anti-mouse IgG antibody | 1:8000 | BS12478 | Bioworld |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, K.; Zhou, X.; Li, C.; Shen, C.; He, G.; Chen, T.; Cao, M.; Chen, X.; Zhang, B.; Chen, L. YTHDF2 as a Mediator in BDNF-Induced Proliferation of Porcine Follicular Granulosa Cells. Int. J. Mol. Sci. 2024, 25, 2343. https://doi.org/10.3390/ijms25042343
Liu K, Zhou X, Li C, Shen C, He G, Chen T, Cao M, Chen X, Zhang B, Chen L. YTHDF2 as a Mediator in BDNF-Induced Proliferation of Porcine Follicular Granulosa Cells. International Journal of Molecular Sciences. 2024; 25(4):2343. https://doi.org/10.3390/ijms25042343
Chicago/Turabian StyleLiu, Kening, Xu Zhou, Chunjin Li, Caomeihui Shen, Guitian He, Tong Chen, Maosheng Cao, Xue Chen, Boqi Zhang, and Lu Chen. 2024. "YTHDF2 as a Mediator in BDNF-Induced Proliferation of Porcine Follicular Granulosa Cells" International Journal of Molecular Sciences 25, no. 4: 2343. https://doi.org/10.3390/ijms25042343
APA StyleLiu, K., Zhou, X., Li, C., Shen, C., He, G., Chen, T., Cao, M., Chen, X., Zhang, B., & Chen, L. (2024). YTHDF2 as a Mediator in BDNF-Induced Proliferation of Porcine Follicular Granulosa Cells. International Journal of Molecular Sciences, 25(4), 2343. https://doi.org/10.3390/ijms25042343