

A Russian Doll of Resistance: Nested Gains and Losses of Venom Immunity in Varanid Lizards
 , , , , and
, , , , and 
Abstract
:1. Introduction
1.1. Venom Evolution and Predator–Prey Dynamics
1.2. Mechanisms of Resistance to Snake Venom α-Neurotoxins
1.3. Varanid Lizard Diversity and Dietary Patterns
1.4. Study Aims and Approach
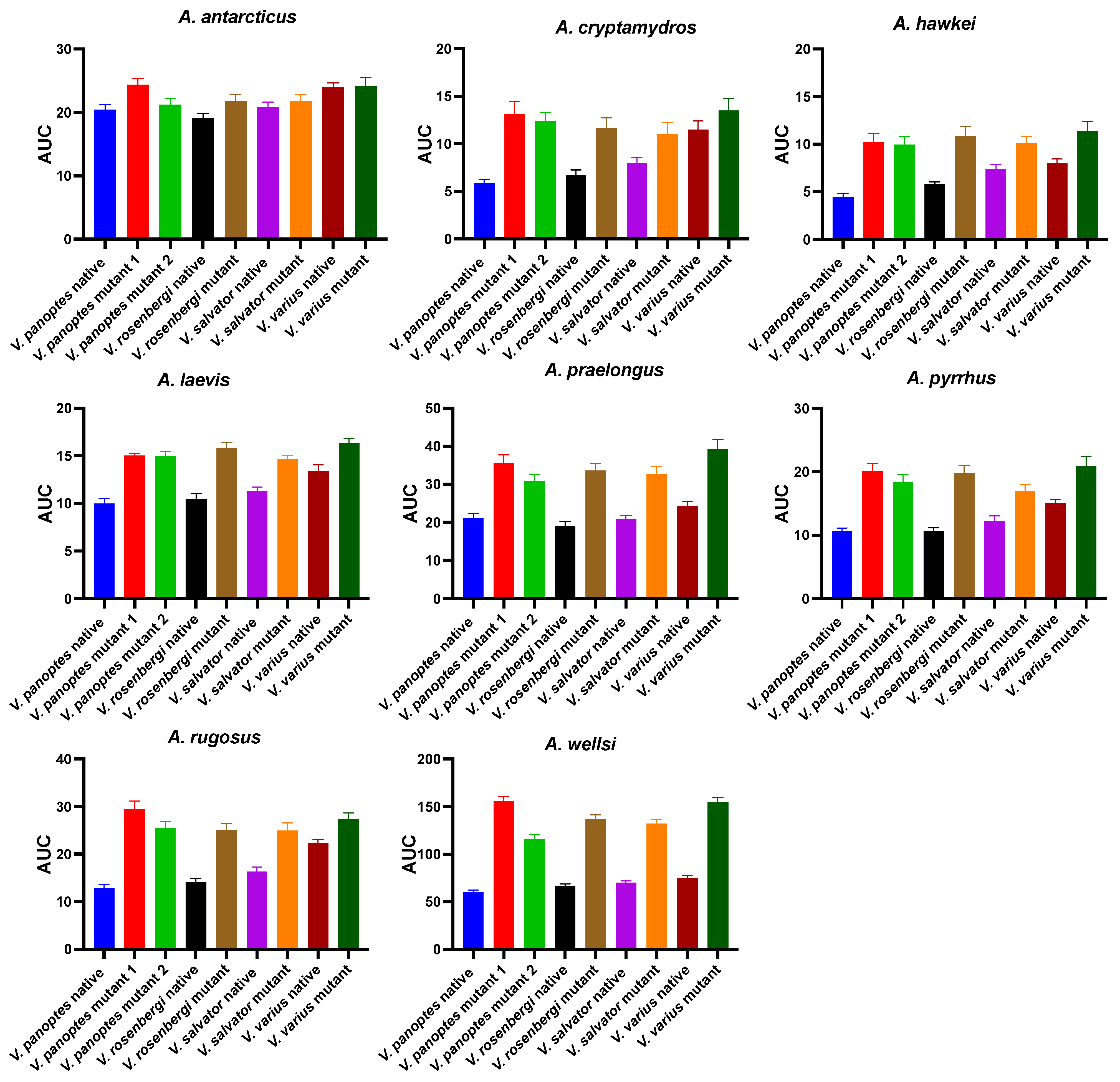
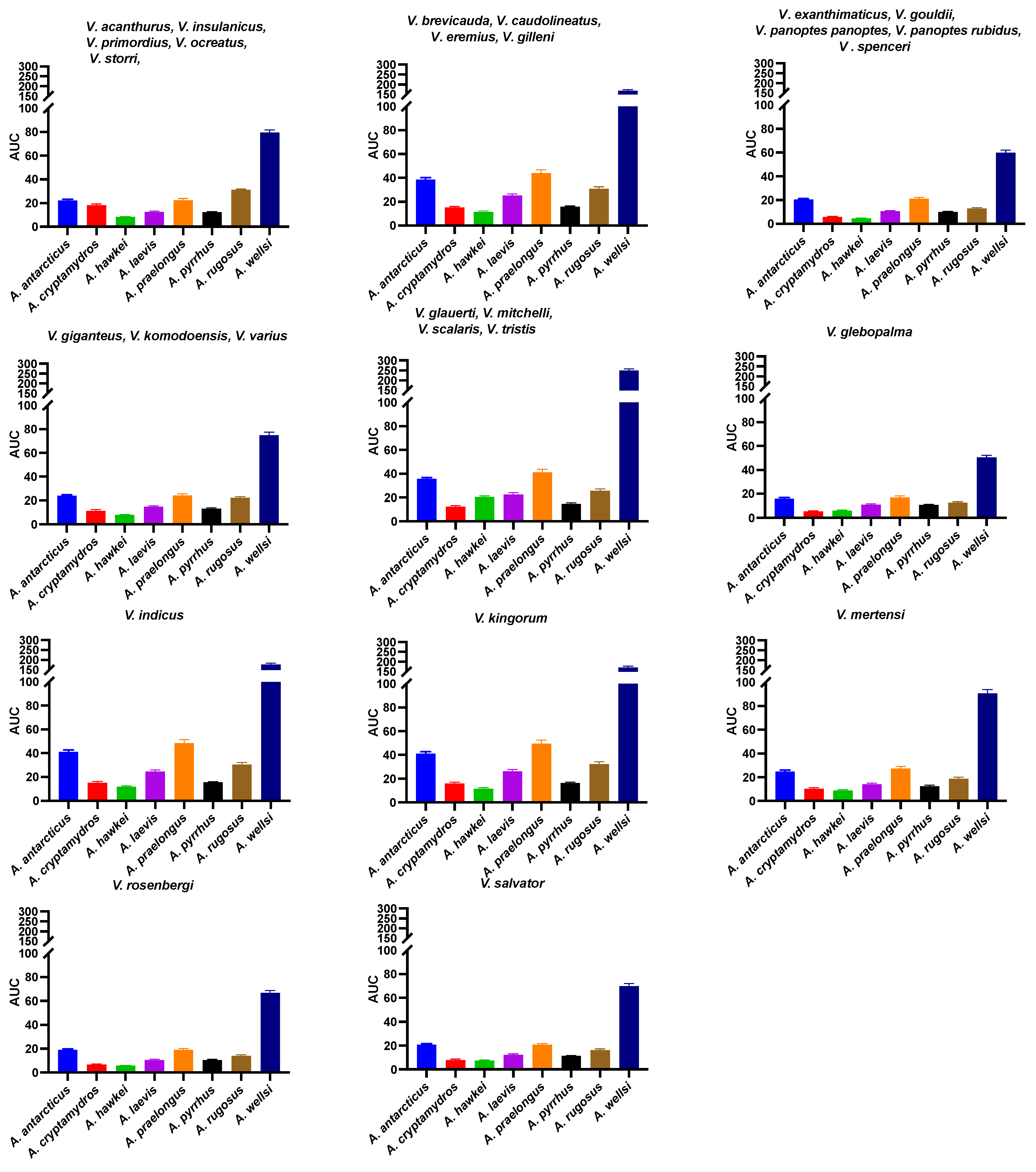
2. Results
2.1. Orthosteric Site Characterization
2.2. Functional Validation of Resistance Motifs
2.3. Sequence Analysis and Hypothesis Testing Using Biolayer Interferometry
2.3.1. Orthosteric Site Position 187
2.3.2. Orthosteric Site Position 189
2.3.3. Orthosteric Site Position 191
2.3.4. Orthosteric Site Position 194
2.3.5. Orthosteric Site Position 195
2.3.6. Orthosteric Site Position 196
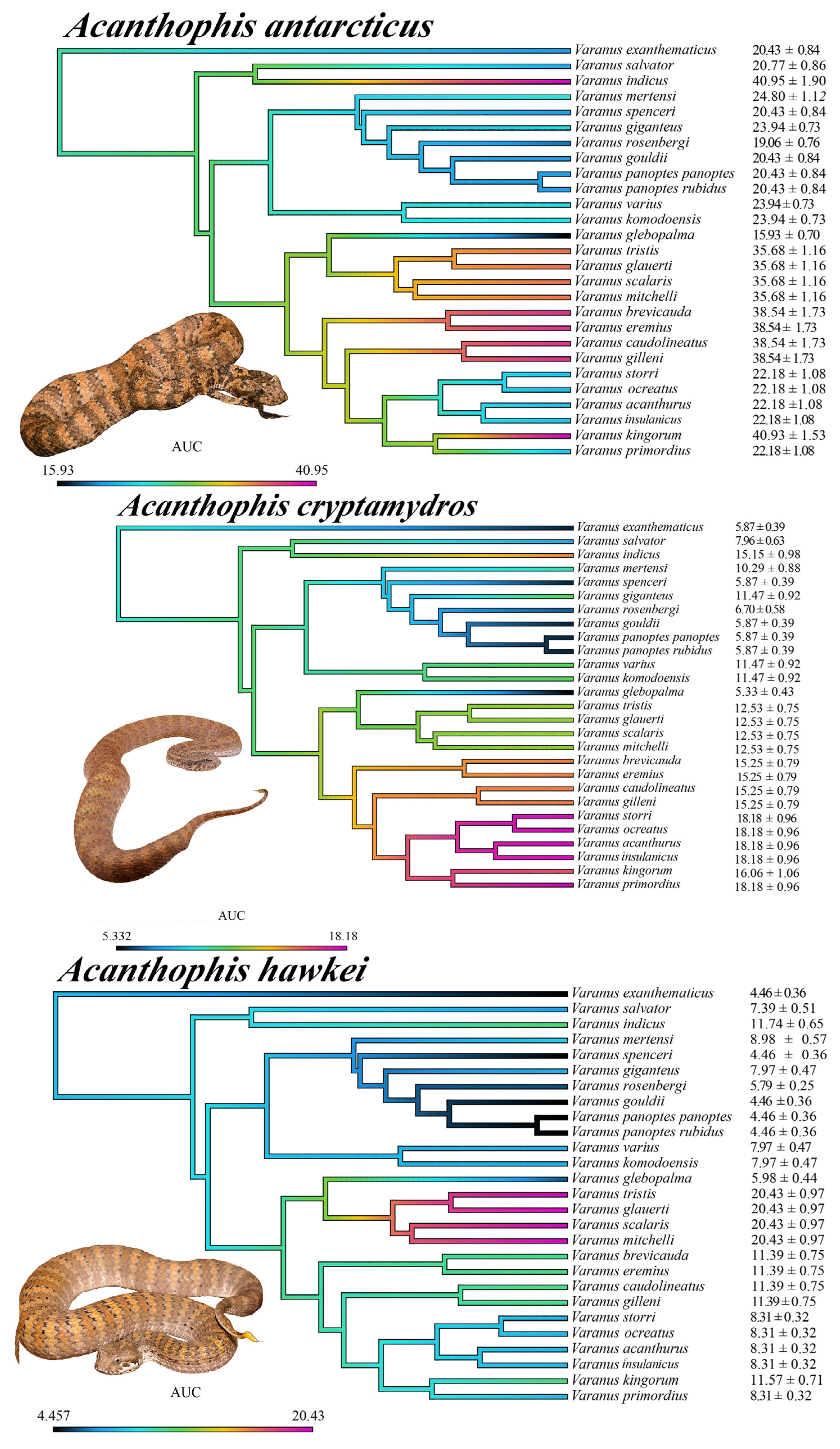
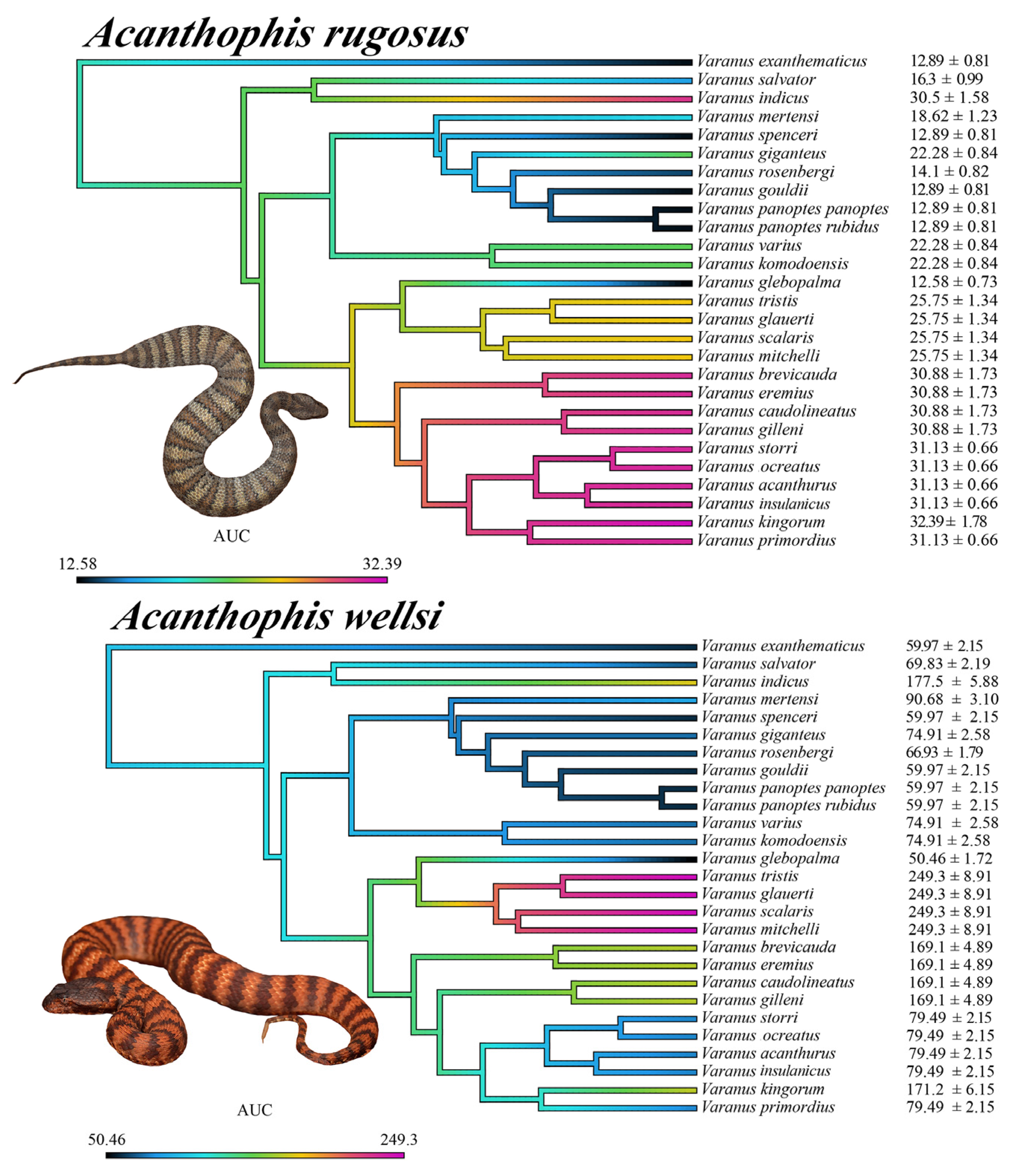
2.3.7. Patterns of Venom Resistance in Varanid Orthosteric Sites and Reciprocal Patterns of Selective Actions by Acanthophis Venoms on Varanid Orthosteric Sites
2.4. Selection Pressure and Adaptive Evolution
- Orthosteric site position 187:
- ○
- Two origins of deviation from the ancestral tryptophan were found:
- ▪
- V. rosenbergi, where it is replaced by a serine.
- ▪
- V. indicus has glutamic acid at this position and V. salvator has glutamine. Considering the phylogenetic relatedness of these species (Figure 3), the most parsimonious explanation is a mutation to either glutamic acid or glutamine occurred in their last common ancestor, and that a subsequent mutation occurred to convert the derived amino acid into the other form. Further sequencing will be necessary, however, to reconstruct the molecular evolutionary history. Regardless, the change in venom affinity was shown to be statistically significant (Table 1).
- Orthosteric site position 189:
- Orthosteric site position 191:
- ○
- The ancestral state was found to be represented by the achiral proteinogenic amino glycine.
- ○
- Consequently, the negatively charged aspartic acid derived state, convergently evolved on three occasions (Figure 3):
- ▪
- Last common ancestor of the V. brevicauda/V. primordius clade.
- ▪
- V. giganteus.
- ○
- Last common ancestor of the V. komodoensis/V. varius clade.
- Orthosteric site position 194:
- ○
- A derivation from the ancestral proline occurred in the last common ancestor of the V. glebopalma/V. primordius clade.
- ○
- Subsequently a secondary reversal occurred in V. glebopalma resulting in the reversal to the resistant state. The change in venom affinity was shown to be statistically significant (Table 1).
- Orthosteric site position 195:
- ○
- Derivation from the ancestral asparagine (R) to aspartic acid (D) convergently occurred on three occasions (Figure 3):
- ▪
- V. mertensi
- ○
- Last common ancestor of the V. glebopalma/V. primordius clade.
- ▪
- Subsequent reversal in V. glebopalma, resulting in the reversal to the resistant state. The change in venom affinity was shown to be statistically significant (Table 1).
- ▪
- Subsequently in the V. brevicauda/V. gilleni clade there was a further derivation to glutamic acid (E).
- Orthosteric site position 196
- ○
- A derivation from the ancestral threonine (T) to isoleucine (I) occurred V. kingorum and is associated with the loss of venom resistance in this species. The change in venom affinity was shown to be statistically significant (Table 1).
3. Discussion
4. Materials and Methods
4.1. DNA Extraction from Varanid Tissue Samples
4.2. Amplification of Orthosteric Site Sequence of α-1 Subunit of nAChR
- Var_Loc_F1 TAAGTAACTACATGGAGAGTGG,
- Var_Loc_R1 TGTGGGCAGATAAAAGACTAAACC
- whilst the primer annealing temperature was set to 55 °C.
4.3. Sequencing of nAChR
4.4. Venom Stock Collection and Preparation
4.5. Mimotope Design and Preparation
4.6. Biolayer Interferometry Assay (BLI)
4.7. Data Acquisition, Processing, and Statistical Analysis
4.8. Evolutionary Analysis
4.9. Phylogenetic Tree Heat Maps
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; et al. The toxicogenomic multiverse: Convergent recruitment of proteins into animal venoms. Annu. Rev. Genom. Hum. Genet. 2009, 10, 483–511. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekara, U.; Harris, R.J.; Fry, B.G. The Target Selects the Toxin: Specific Amino Acids in Snake-Prey Nicotinic Acetylcholine Receptors That Are Selectively Bound by King Cobra Venoms. Toxins 2022, 14, 528. [Google Scholar] [CrossRef] [PubMed]
- van Thiel, J.; Khan, M.A.; Wouters, R.M.; Harris, R.J.; Casewell, N.R.; Fry, B.G.; Kini, R.M.; Mackessy, S.P.; Vonk, F.J.; Wüster, W. Convergent evolution of toxin resistance in animals. Biol. Rev. 2022, 97, 1823–1843. [Google Scholar] [CrossRef] [PubMed]
- Barchan, D.; Kachalsky, S.; Neumann, D.; Vogel, Z.; Ovadia, M.; Kochva, E.; Fuchs, S. How the mongoose can fight the snake—The binding-site of the mongoose acetylcholine-receptor. Proc. Natl. Acad. Sci. USA 1992, 89, 7717–7721. [Google Scholar] [CrossRef] [PubMed]
- Barchan, D.; Ovadia, M.; Kochva, E.; Fuchs, S. The binding-site of the nicotinic acetylcholine-receptor in animal species resistant to alpha-bungarotoxin. Biochemistry 1995, 34, 9172–9176. [Google Scholar] [CrossRef]
- Geffeney, S.L.; Fujimoto, E.; Brodie, E.D., 3rd; Brodie, E.D., Jr.; Ruben, P.C. Evolutionary diversification of TTX-resistant sodium channels in a predator-prey interaction. Nature 2005, 434, 759–763. [Google Scholar] [CrossRef]
- Jansa, S.A.; Voss, R.S. Adaptive evolution of the venom-targeted vWF protein in opossums that eat pitvipers. PLoS ONE 2011, 6, e20997. [Google Scholar] [CrossRef]
- Tarvin, R.D.; Borghese, C.M.; Sachs, W.; Santos, J.C.; Lu, Y.; O’Connell, L.A.; Cannatella, D.C.; Harris, R.A.; Zakon, H.H. Interacting amino acid replacements allow poison frogs to evolve epibatidine resistance. Science 2017, 357, 1261–1266. [Google Scholar] [CrossRef]
- Karageorgi, M.; Groen, S.C.; Sumbul, F.; Pelaez, J.N.; Verster, K.I.; Aguilar, J.M.; Hastings, A.P.; Bernstein, S.L.; Matsunaga, T.; Astourian, M.; et al. Genome editing retraces the evolution of toxin resistance in the monarch butterfly. Nature 2019, 574, 409–412. [Google Scholar] [CrossRef]
- Solé, R. Revisiting Leigh Van Valen’s “A New Evolutionary Law” (1973). Biol. Theory 2022, 17, 120–125. [Google Scholar] [CrossRef]
- Holding, M.L.; Biardi, J.E.; Gibbs, H.L. Coevolution of venom function and venom resistance in a rattlesnake predator and its squirrel prey. Proc. Biol. Sci. R. Soc. 2016, 283, 2841. [Google Scholar] [CrossRef] [PubMed]
- Holding, M.L.; Drabeck, D.H.; Jansa, S.A.; Gibbs, H.L. Venom Resistance as a Model for Understanding the Molecular Basis of Complex Coevolutionary Adaptations. Integr. Comp. Biol. 2016, 56, 1032–1043. [Google Scholar] [CrossRef] [PubMed]
- Biardi, J.; Coss, R.; Smith, D. California ground squirrel (Spermophilus beecheyi) blood sera inhibits crotalid venom proteolytic activity. Toxicon 2000, 38, 713–721. [Google Scholar] [CrossRef]
- Barlow, A.; Pook, C.E.; Harrison, R.A.; Wuster, W. Coevolution of diet and prey-specific venom activity supports the role of selection in snake venom evolution. Proc. Biol. Sci. R. Soc. 2009, 276, 2443–2449. [Google Scholar] [CrossRef] [PubMed]
- Aird, S.D.; Aggarwal, S.; Villar-Briones, A.; Tin, M.M.; Terada, K.; Mikheyev, A.S. Snake venoms are integrated systems, but abundant venom proteins evolve more rapidly. BMC Genom. 2015, 16, 647. [Google Scholar] [CrossRef] [PubMed]
- Kachalsky, S.G.; Jensen, B.S.; Barchan, D.; Fuchs, S. Two subsites in the binding domain of the acetylcholine receptor: An aromatic subsite and a proline subsite. Proc. Natl. Acad. Sci. USA 1995, 92, 10801–10805. [Google Scholar] [CrossRef] [PubMed]
- Asher, O.; Jensen, B.S.; Lupu-Meiri, M.; Oron, Y.; Fuchs, S. The mongoose acetylcholine receptor α-subunit: Analysis of glycosylation and α-bungarotoxin binding. FEBS Lett. 1998, 426, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Takacs, Z.; Wilhelmsen, K.C.; Sorota, S. Snake α-neurotoxin binding site on the Egyptian Cobra (Naja haje) nicotinic acetylcholine receptor is conserved. Mol. Biol. Evol. 2001, 18, 1800–1809. [Google Scholar] [CrossRef]
- Takacs, Z.; Wilhelmsen, K.C.; Sorota, S. Cobra (Naja spp.) nicotinic acetylcholine receptor exhibits resistance to Erabu sea snake (Laticauda semifasciata) short-chain alpha-neurotoxin. J. Mol. Evol. 2004, 58, 516–526. [Google Scholar] [CrossRef]
- Dellisanti, C.D.; Yao, Y.; Stroud, J.C.; Wang, Z.Z.; Chen, L. Structural determinants for alpha-neurotoxin sensitivity in muscle nAChR and their implications for the gating mechanism. Channels 2007, 1, 234–237. [Google Scholar] [CrossRef]
- Rahman, M.M.; Teng, J.; Worrell, B.T.; Noviello, C.M.; Lee, M.; Karlin, A.; Stowell, M.H.B.; Hibbs, R.E. Structure of the Native Muscle-type Nicotinic Receptor and Inhibition by Snake Venom Toxins. Neuron 2020, 106, 952–962. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.J.; Fry, B.G. Electrostatic resistance to alpha-neurotoxins conferred by charge reversal mutations in nicotinic acetylcholine receptors. Proc. Biol. Sci. R. Soc. 2021, 288, 20202703. [Google Scholar] [CrossRef]
- Harris, R.J.; Nekaris, K.A.; Fry, B.G. Monkeying around with venom: An increased resistance to alpha-neurotoxins supports an evolutionary arms race between Afro-Asian primates and sympatric cobras. BMC Biol. 2021, 19, 253. [Google Scholar] [CrossRef] [PubMed]
- Jones, L.; Harris, R.J.; Fry, B.G. Not Goanna Get Me: Mutations in the Savannah Monitor Lizard (Varanus exanthematicus) Nicotinic Acetylcholine Receptor Confer Reduced Susceptibility to Sympatric Cobra Venoms. Neurotox. Res. 2021, 39, 1116–1122. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Dashevsky, D.; Kerkkamp, H.; Kordis, D.; de Bakker, M.A.G.; Wouters, R.; van Thiel, J.; den Brouw, B.O.; Vonk, F.; Kini, R.M.; et al. Widespread Evolution of Molecular Resistance to Snake Venom alpha-Neurotoxins in Vertebrates. Toxins 2020, 12, 638. [Google Scholar] [CrossRef] [PubMed]
- Drabeck, D.H.; Holt, J.; McGaugh, S.E. Widespread convergent evolution of alpha-neurotoxin resistance in African mammals. Biol. Lett. 2022, 18, 20220361. [Google Scholar] [CrossRef] [PubMed]
- Mancuso, M.; Zaman, S.; Maddock, S.T.; Kamei, R.G.; Salazar-Valenzuela, D.; Wilkinson, M.; Roelants, K.; Fry, B.G. Resistance Is Not Futile: Widespread Convergent Evolution of Resistance to Alpha-Neurotoxic Snake Venoms in Caecilians (Amphibia: Gymnophiona). Int. J. Mol. Sci. 2023, 24, 11353. [Google Scholar] [CrossRef]
- Drabeck, D.H.; Dean, A.M.; Jansa, S.A. Why the honey badger don’t care: Convergent evolution of venom-targeted nicotinic acetylcholine receptors in mammals that survive venomous snake bites. Toxicon 2015, 99, 68–72. [Google Scholar] [CrossRef]
- Brennan, I.G.; Lemmon, A.R.; Lemmon, E.M.; Portik, D.M.; Weijola, V.; Welton, L.; Donnellan, S.C.; Keogh, J.S. Phylogenomics of monitor lizards and the role of competition in dictating body size disparity. Syst. Biol. 2021, 70, 120–132. [Google Scholar] [CrossRef]
- Pianka, E.R. Comparative ecology of Varanus in the Great Victoria Desert. Aust. J. Ecol. 1994, 19, 395–408. [Google Scholar] [CrossRef]
- Shine, R. Food habits, habitats and reproductive biology of four sympatric species of varanid lizards in tropical Australia. Herpetologica 1986, 42, 346–360. [Google Scholar]
- Mayes, P.; Thompson, G.; Withers, P. Diet and foraging behaviour of the semi-aquatic Varanus mertensi (Reptilia: Varanidae). ECU Publ. 2005, 32, 67–74. [Google Scholar]
- Cogger, H. Reptiles and Amphibians of Australia, 7th ed.; CSIRO Publishing: Melbourne, VIC, Australia, 2014. [Google Scholar]
- Sutherland, S.K.; Tibbals, J. Australian Animal Toxins: The Creatures, Their Toxins and Care of the Poisoned Patient, 2nd ed.; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Gotti, C.; Mazzola, G.; Longhi, R.; Fornasari, D.; Clementi, F. The binding site for α-bungarotoxin resides in the sequence 188–201 of the α-subunit of acetylcholine receptor: Structure, conformation and binding characteristics of peptide [Lys] 188–201. Neurosci. Lett. 1987, 82, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Testai, F.D.; Venera, G.D.; Peña, C.; de Jiménez Bonino, M.J.B. Histidine 186 of the nicotinic acetylcholine receptor α subunit requires the presence of the 192–193 disulfide bridge to interact with α-bungarotoxin. Neurochem. Int. 2000, 36, 27–33. [Google Scholar] [CrossRef]
- Sine, S.M.; Quiram, P.; Papanikolaou, F.; Kreienkamp, H.J.; Taylor, P. Conserved tyrosines in the alpha subunit of the nicotinic acetylcholine receptor stabilize quaternary ammonium groups of agonists and curariform antagonists. J. Biol. Chem. 1994, 269, 8808–8816. [Google Scholar] [CrossRef] [PubMed]
- Thompson, G.; Clemente, C.; Withers, P.; Norman, J. Is body shape of varanid lizards linked with retreat choice. ECU Publ. 2008, 56, 351–362. [Google Scholar] [CrossRef]
- Godwin, C.; Doody, J.; Rhind, D.; Clulow, S.; Soennichsen, K.; Murray, C.; Bartek, B.; Severin, A.; Severin, L. Varanus gouldii (Gould’s monitor). Diet and cannibalism. Herpetol. Rev. 2020, 51, 137–138. [Google Scholar]
- Pavon-Vazquez, C.J.; Esquerre, D.; Fitch, A.J.; Maryan, B.; Doughty, P.; Donnellan, S.C.; Keogh, J.S. Between a rock and a dry place: Phylogenomics, biogeography, and systematics of ridge-tailed monitors (Squamata: Varanidae: Varanus acanthurus complex). Mol. Phylogenet Evol. 2022, 173, 107516. [Google Scholar] [CrossRef]
- Harris, R.J.; Zdenek, C.N.; Harrich, D.; Frank, N.; Fry, B.G. An Appetite for Destruction: Detecting Prey-Selective Binding of alpha-Neurotoxins in the Venom of Afro-Asian Elapids. Toxins 2020, 12, 205. [Google Scholar] [CrossRef]
- Zdenek, C.N.; Harris, R.J.; Kuruppu, S.; Youngman, N.J.; Dobson, J.S.; Debono, J.; Khan, M.; Smith, I.; Yarski, M.; Harrich, D.; et al. A Taxon-Specific and High-Throughput Method for Measuring Ligand Binding to Nicotinic Acetylcholine Receptors. Toxins 2019, 11, 600. [Google Scholar] [CrossRef]
- Bracci, L.; Lozzi, L.; Lelli, B.; Pini, A.; Neri, P. Mimotopes of the nicotinic receptor binding site selected by a combinatorial peptide library. Biochemistry 2001, 40, 6611–6619. [Google Scholar] [CrossRef] [PubMed]
- Fry, B.G.; Wickramaratna, J.C.; Hodgson, W.C.; Alewood, P.F.; Kini, R.M.; Ho, H.; Wuster, W. Electrospray liquid chromatography/mass spectrometry fingerprinting of Acanthophis (death adder) venoms: Taxonomic and toxinological implications. Rapid Commun. Mass. Spectrom. 2002, 16, 600–608. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Yang, Z. PAMLX: A graphical user interface for PAML. Mol. Biol. Evol. 2013, 30, 2723–2724. [Google Scholar] [CrossRef] [PubMed]
- Maddison, W.P.; Maddison, D.R. Mesqite: A Modular System for Evolutionary Analysis. Version 3.80. 2023. Available online: http://www.mesquiteproject.org (accessed on 1 August 2023).
- Smith, J.G.; Christian, K.; Green, B. Physiological Ecology of the Mangrove-Dwelling Varanid Varanus indicus. Physiol. Biochem. Zool. 2008, 81, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Dryden, G. The food and feeding habits of Varanus indicus on Guam. Micronesica 1965, 2, 73–76. [Google Scholar]
- Dobson, J.S.; Harris, R.J.; Zdenek, C.N.; Huynh, T.; Hodgson, W.C.; Bosmans, F.; Fourmy, R.; Violette, A.; Fry, B.G. The Dragon’s Paralysing Spell: Evidence of Sodium and Calcium Ion Channel Binding Neurotoxins in Helodermatid and Varanid Lizard Venoms. Toxins 2021, 13, 549. [Google Scholar] [CrossRef] [PubMed]
- Dobson, J.S.; Zdenek, C.N.; Hay, C.; Violette, A.; Fourmy, R.; Cochran, C.; Fry, B.G. Varanid Lizard Venoms Disrupt the Clotting Ability of Human Fibrinogen through Destructive Cleavage. Toxins 2019, 11, 255. [Google Scholar] [CrossRef] [PubMed]
- Koludarov, I.; Jackson, T.N.; op den Brouw, B.; Dobson, J.; Dashevsky, D.; Arbuckle, K.; Clemente, C.J.; Stockdale, E.J.; Cochran, C.; Debono, J. Enter the dragon: The dynamic and multifunctional evolution of Anguimorpha lizard venoms. Toxins 2017, 9, 242. [Google Scholar] [CrossRef]
- Fry, B.G.; Wroe, S.; Teeuwisse, W.; van Osch, M.J.; Moreno, K.; Ingle, J.; McHenry, C.; Ferrara, T.; Clausen, P.; Scheib, H.; et al. A central role for venom in predation by Varanus komodoensis (Komodo Dragon) and the extinct giant Varanus (Megalania) priscus. Proc. Natl. Acad. Sci. USA 2009, 106, 8969–8974. [Google Scholar] [CrossRef]
- Harris, R.J.; Youngman, N.J.; Chan, W.; Bosmans, F.; Cheney, K.L.; Fry, B.G. Getting stoned: Characterisation of the coagulotoxic and neurotoxic effects of reef stonefish (Synanceia verrucosa) venom. Toxicol. Lett. 2021, 346, 16–22. [Google Scholar] [CrossRef]
- Harris, R.J.; Youngman, N.J.; Zdenek, C.N.; Huynh, T.M.; Nouwens, A.; Hodgson, W.C.; Harrich, D.; Dunstan, N.; Portes-Junior, J.A.; Fry, B.G. Assessing the Binding of Venoms from Aquatic Elapids to the Nicotinic Acetylcholine Receptor Orthosteric Site of Different Prey Models. Int. J. Mol. Sci. 2020, 21, 97377. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.J.; Zdenek, C.N.; Debono, J.; Harrich, D.; Fry, B.G. Evolutionary Interpretations of Nicotinic Acetylcholine Receptor Targeting Venom Effects by a Clade of Asian Viperidae Snakes. Neurotox. Res. 2020, 38, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.J.; Zdenek, C.N.; Nouwens, A.; Sweeney, C.; Dunstan, N.; Fry, B.G. A symmetry or asymmetry: Functional and compositional comparison of venom from the left and right glands of the Indochinese spitting cobra (Naja siamensis). Toxicon X 2020, 7, 100050. [Google Scholar] [CrossRef] [PubMed]
- Pianka, E.R. Evolution of Body Size: Varanid Lizards as a Model System. Am. Nat. 1995, 146, 398–414. [Google Scholar] [CrossRef]
- Kumar, S.; Suleski, M.; Craig, J.M.; Kasprowicz, A.E.; Sanderford, M.; Li, M.; Stecher, G.; Hedges, S.B. TimeTree 5: An expanded resource for species divergence times. Mol. Biol. Evol. 2022, 39, msac174. [Google Scholar] [CrossRef] [PubMed]
- Bourke, L.A.; Zdenek, C.N.; Tanaka-Azevedo, A.M.; Silveira, G.P.M.; Sant’Anna, S.S.; Grego, K.F.; Rodrigues, C.F.B.; Fry, B.G. Clinical and Evolutionary Implications of Dynamic Coagulotoxicity Divergences in Bothrops (Lancehead Pit Viper) Venoms. Toxins 2022, 14, 50297. [Google Scholar] [CrossRef] [PubMed]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [PubMed]
- Revell, L.J. phytools: An R package for phylogenetic comparative biology (and other things). Methods Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| V. kingorum vs. Ancestral Clade 1 p = | V. indicus vs. V. salvator p = | V. glebopalma vs. Ancestral Clade 2 p = | V. komodoensis/V. varius and V. giganteus 3 vs. Ancestral Clade 4 p = | |
|---|---|---|---|---|
| A. antarcticus | 0.0001 | 0.0007 | <0.0001 | 0.0058 |
| A. cryptomydros | 0.0641 | 0.0010 | 0.0005 | 0.0036 |
| A. hawkei | 0.0071 | 0.0010 | 0.0003 | 0.0007 |
| A. laevis | 0.0007 | 0.0002 | 0.0020 | 0.0031 |
| A. praelongus | 0.0009 | 0.0017 | 0.0012 | 0.0301 |
| A. rugosus | 0.3472 | 0.0005 | 0.0006 | 0.0260 |
| A. pyrrhus | 0.0010 | 0.0002 | 0.0008 | 0.0010 |
| A. wellsi | 0.0005 | 0.0003 | 0.0004 | 0.0017 |
| ω Values | |||
|---|---|---|---|
| Selection Model | Clade/Branch Set | Toxin-Binding (Orthosteric) Site | Flanking Regions |
| One-ratio model | All varanids (27 spp.) | 1.0231 | 0.1791 |
| Branch model | “Small” varanids (SVL < 500 m) | 0.8237 | 0.0001 |
| “Large” varanids (SVL > 500 m) | 1.4107 | 0.3245 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chandrasekara, U.; Mancuso, M.; Seneci, L.; Bourke, L.; Trembath, D.F.; Sumner, J.; Zdenek, C.N.; Fry, B.G. A Russian Doll of Resistance: Nested Gains and Losses of Venom Immunity in Varanid Lizards. Int. J. Mol. Sci. 2024, 25, 2628. https://doi.org/10.3390/ijms25052628
Chandrasekara U, Mancuso M, Seneci L, Bourke L, Trembath DF, Sumner J, Zdenek CN, Fry BG. A Russian Doll of Resistance: Nested Gains and Losses of Venom Immunity in Varanid Lizards. International Journal of Molecular Sciences. 2024; 25(5):2628. https://doi.org/10.3390/ijms25052628
Chicago/Turabian StyleChandrasekara, Uthpala, Marco Mancuso, Lorenzo Seneci, Lachlan Bourke, Dane F. Trembath, Joanna Sumner, Christina N. Zdenek, and Bryan G. Fry. 2024. "A Russian Doll of Resistance: Nested Gains and Losses of Venom Immunity in Varanid Lizards" International Journal of Molecular Sciences 25, no. 5: 2628. https://doi.org/10.3390/ijms25052628
APA StyleChandrasekara, U., Mancuso, M., Seneci, L., Bourke, L., Trembath, D. F., Sumner, J., Zdenek, C. N., & Fry, B. G. (2024). A Russian Doll of Resistance: Nested Gains and Losses of Venom Immunity in Varanid Lizards. International Journal of Molecular Sciences, 25(5), 2628. https://doi.org/10.3390/ijms25052628