
Selection and Validation of qRT-PCR Internal Reference Genes to Study Flower Color Formation in Camellia impressinervis

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
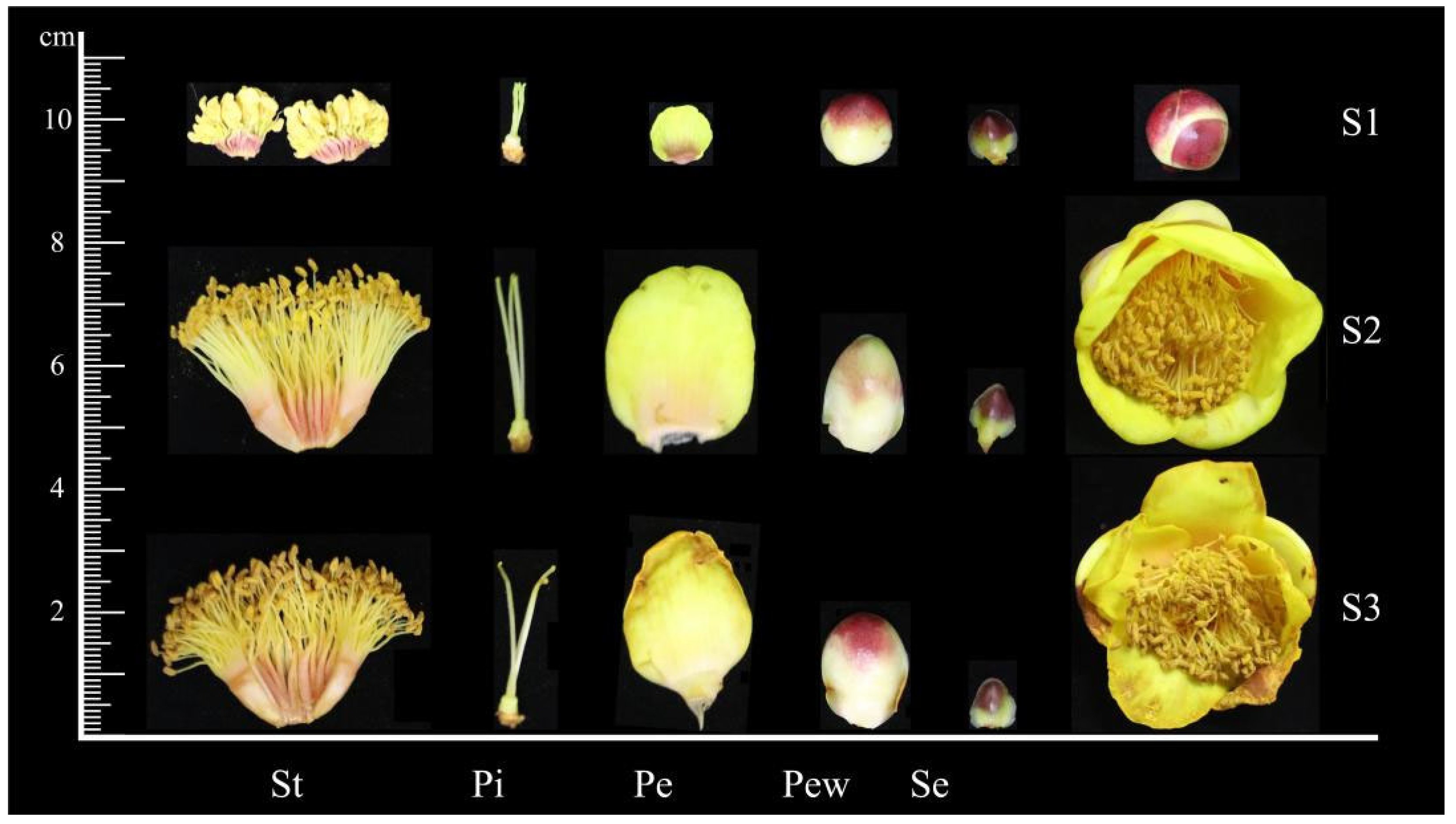
2.1. Petal Pigment Categories
2.2. Petal Pigment Content
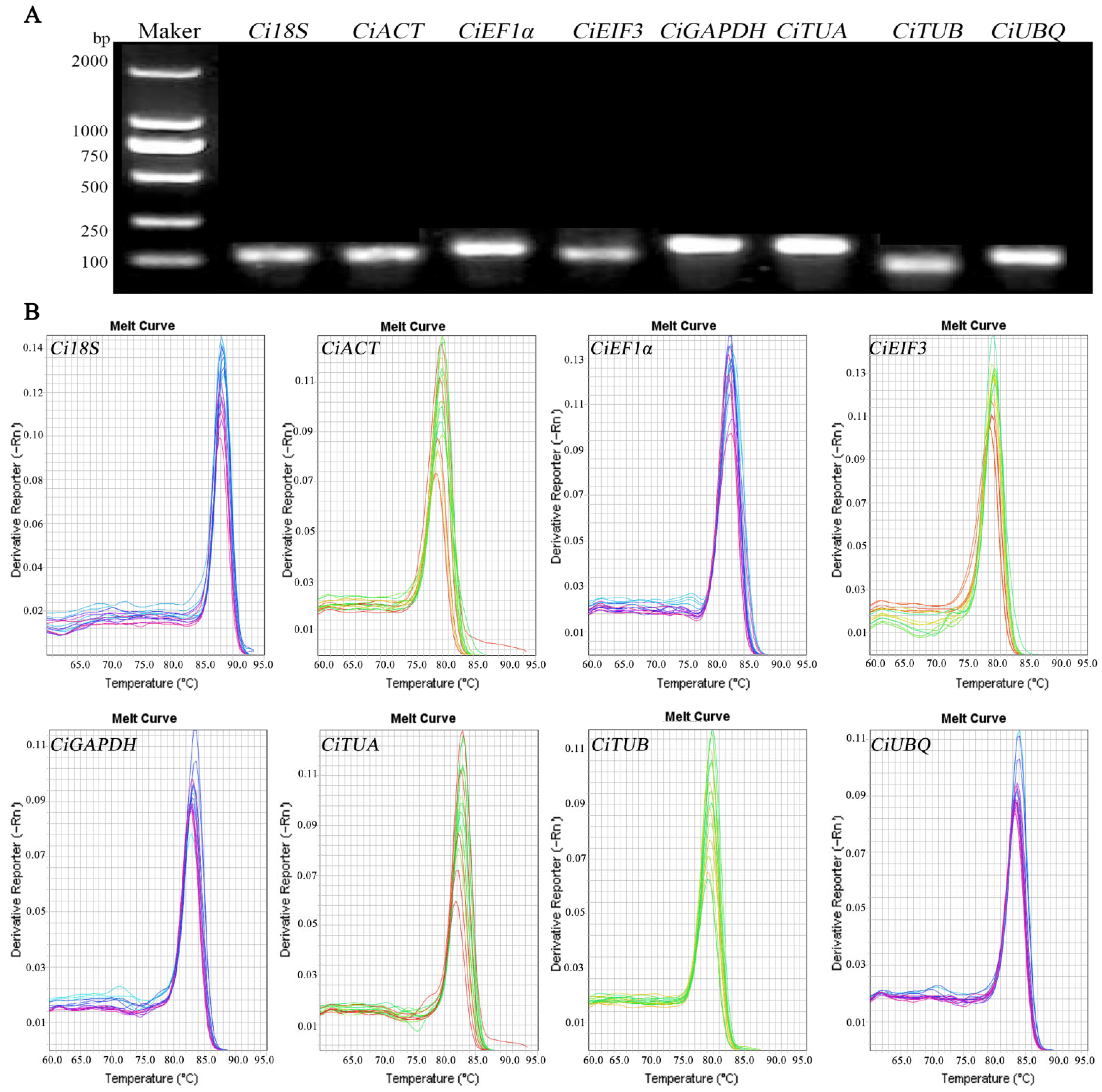
2.3. Primer Specificity Analysis
2.4. Analysis of Candidate Gene Expression Abundance
2.5. geNorm Analysis
2.6. NormFinder Analysis
2.7. BestKeeper Analysis
2.8. Comprehensive Analysis of Internal Parameter Stability
2.9. Expression Analysis of Flower Color Related Genes
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Identification of the Petal Pigment Categories
4.3. Determination of Flavonoids and Carotenoids Content
4.4. RNA Extraction and cDNA Synthesis
4.5. Primer Design for Candidate Genes and Flower Color Related Genes
4.6. Candidate Reference Gene qRT-PCR Analysis
4.7. Stability Evaluation of Candidate Genes
4.8. Verification of Stability of Candidate Internal Reference Genes
4.9. Data Processing
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Trinh, T.D. Yellow Camellias: A Review of Chemical Constituents and Biological Activities. Dalat Univ. J. Sci. 2022, 12, 117–144. [Google Scholar] [CrossRef]
- Dai, L.; Li, J.; Liang, X.; Li, L.; Feng, Y.; Liu, H.; Wei, W.; Ning, S.; Zhang, L. Flowers of Camellia nitidissima cause growth inhibition, cell-cycle dysregulation and apoptosis in a human esophageal squamous cell carcinoma cell line. Mol. Med. Rep. 2016, 14, 1117–1122. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Hao, E.; Zhang, F.; Du, Z.; Xie, J.; Chen, F.; Yu, C.; Hou, X.; Deng, J. Identifying Active Compounds and Mechanism of Camellia nitidissima Chi on Anti-Colon Cancer by Network Pharmacology and Experimental Validation. Evid.-Based Compl. Alt. 2021, 2021, 7169211. [Google Scholar] [CrossRef]
- Scogin, R. Floral Pigments of the Yellow Camellia, Camellia chrysantha (Theaceae). Aliso 1986, 11, 387–392. [Google Scholar] [CrossRef]
- Vignolini, S.; Moyroud, E.; Glover, B.J.; Steiner, U. Analysing photonic structures in plants. J. R. Soc. Interface 2013, 10, 20130394. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Li, X.; Sai, X.; Wang, L.; Li, S.; Xu, Y. Camellia nitidissima C.W. Chi: A review of botany, chemistry, and pharmacology. Phytochem. Rev. 2018, 17, 327–349. [Google Scholar] [CrossRef]
- Rudall, P.J. Colourful cones: How did flower colour first evolve? J. Exp. Bot. 2020, 71, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Grotewold, E. The genetics and biochemistry of floral pigments. Annu. Rev. Plant Biol. 2006, 57, 761–780. [Google Scholar] [CrossRef]
- Dao, T.T.; Linthorst, H.J.; Verpoorte, R. Chalcone synthase and its functions in plant resistance. Phytochem. Rev. 2011, 10, 397–412. [Google Scholar] [CrossRef]
- Feng, F.; Li, M.; Ma, F.; Cheng, L. Phenylpropanoid metabolites and expression of key genes involved in anthocyanin biosynthesis in the shaded peel of apple fruit in response to sun exposure. Plant Physiol. Biochem. 2013, 69, 54–61. [Google Scholar] [CrossRef]
- Kayani, S.; Shen, Q.; Rahman, S.; Fu, X.; Li, Y.; Wang, C.; Hassani, D.; Tang, K. Transcriptional regulation of flavonoid biosynthesis in Artemisia annua by AaYABBY5. Hortic. Res. 2021, 8, 257. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Yamamura, S.; Nishihara, M.; Koiwa, H.; Sandmann, G. cDNAs for the synthesis of cyclic carotenoids in petals of Gentiana lutea and their regulation during flower development. Biochim. Biophys. Acta (BBA) Gene Struct. Expr. 2003, 1625, 305–308. [Google Scholar] [CrossRef]
- Obrero, Á.; González-Verdejo, C.I.; Die, J.V.; Gómez, P.; Del Río-Celestino, M.; Román, B. Carotenogenic Gene Expression and Carotenoid Accumulation in Three Varieties of Cucurbita pepo during Fruit Development. J. Agric. Food Chem. 2013, 61, 6393–6403. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Jiang, M.; Liang, S.; Zheng, H.; Chen, Q.; Wang, Y.; Lin, Y.; Liu, Z.; Wang, X.; Zhang, F.; et al. Functional differences of BaPDS1 and BaPDS2 genes in Chinese kale. R. Soc. Open Sci. 2019, 6, 190260. [Google Scholar] [CrossRef]
- Zhang, N.; Ma, X.; Li, R.; Xue, Y.; Sun, Y.; Nie, S.; Zhang, L. Transcriptome-based analysis of carotenoid accumulation-related gene expression in petals of Chinese cabbage (Brassica rapa L.). 3 Biotech 2019, 9, 274. [Google Scholar] [CrossRef]
- Solangi, Z.A.; Zhang, Y.; Li, K.; Du, D.; Yao, Y. Fine mapping and candidate gene analysis of the orange petal colour gene Bnpc2 in spring rapeseed (Brassica napus). Plant Breed. 2021, 140, 294–304. [Google Scholar] [CrossRef]
- Jiang, L.; Fan, Z.; Tong, R.; Zhou, X.; Li, J.; Yin, H. Functional Diversification of the Dihydroflavonol 4-Reductase from Camellia nitidissima Chi. in the Control of Polyphenol Biosynthesis. Genes 2020, 11, 1341. [Google Scholar] [CrossRef]
- Jiang, L.; Fan, Z.; Tong, R.; Yin, H.; Li, J.; Zhou, X. Flavonoid 3′-hydroxylase of Camellia nitidissima Chi. promotes the synthesis of polyphenols better than flavonoids. Mol. Biol. Rep. 2021, 48, 3903–3912. [Google Scholar] [CrossRef] [PubMed]
- Giulietti, A.; Overbergh, L.; Valckx, D.; Decallonne, B.; Bouillon, R.; Mathieu, C. An Overview of Real-Time Quantitative PCR: Applications to Quantify Cytokine Gene Expression. Methods 2001, 25, 386–401. [Google Scholar] [CrossRef] [PubMed]
- Nolan, T.; Hands, R.E.; Bustin, S.A. Quantification of mRNA using real-time RT-PCR. Nat. Protoc. 2006, 1, 1559–1582. [Google Scholar] [CrossRef]
- Kou, X.; Zhang, L.; Yang, S.; Li, G.; Ye, J. Selection and validation of reference genes for quantitative RT-PCR analysis in peach fruit under different experimental conditions. Sci. Hortic. 2017, 225, 195–203. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Robledo, D.; Hernández-Urcera, J.; Cal, R.M.; Pardo, B.G.; Sánchez, L.; Martínez, P.; Viñas, A. Analysis of qPCR reference gene stability determination methods and a practical approach for efficiency calculation on a turbot (Scophthalmus maximus) gonad dataset. BMC Genom. 2014, 15, 648. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Xu, H.; Cao, Y.; Yang, P.; Feng, Y.; Tang, Y.; Yuan, S.; Ming, J. Validation of Reference Genes for Quantitative Real-Time PCR during Bicolor Tepal Development in Asiatic Hybrid Lilies (Lilium spp.). Front. Plant Sci. 2017, 8, 669. [Google Scholar] [CrossRef]
- Long, X.; Wang, J.; Ouellet, T.; Rocheleau, H.; Wei, Y.; Pu, Z.; Jiang, Q.; Lan, X.; Zheng, Y. Genome-wide identification and evaluation of novel internal control genes for Q-PCR based transcript normalization in wheat. Plant Mol. Biol. 2010, 74, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Kozera, B.; Rapacz, M. Reference genes in real-time PCR. J. Appl. Genet. 2013, 54, 391–406. [Google Scholar] [CrossRef] [PubMed]
- Chapman, J.R.; Waldenström, J. With Reference to Reference Genes: A Systematic Review of Endogenous Controls in Gene Expression Studies. PLoS ONE 2015, 10, e141853. [Google Scholar] [CrossRef]
- Li, T.; Wang, J.; Lu, M.; Zhang, T.; Qu, X.; Wang, Z. Selection and Validation of Appropriate Reference Genes for qRT-PCR Analysis in Isatis indigotica Fort. Front. Plant Sci. 2017, 8, 1139. [Google Scholar] [CrossRef]
- Liu, Q.; Qi, X.; Yan, H.; Huang, L.; Nie, G.; Zhang, X. Reference gene selection for quantitative real-time reverse-transcriptase PCR in annual ryegrass (Lolium multiflorum) subjected to various abiotic stresses. Molecules 2018, 23, 172–184. [Google Scholar] [CrossRef]
- Gutierrez, L.; Mauriat, M.; Guénin, S.; Pelloux, J.; Lefebvre, J.F.; Louvet, R.; Rusterucci, C.; Moritz, T.; Guerineau, F.; Bellini, C.; et al. The lack of a systematic validation of reference genes: A serious pitfall undervalued in reverse transcription-polymerase chain reaction (RT-PCR) analysis in plants. Plant Biotechnol. J. 2008, 6, 609–618. [Google Scholar] [CrossRef]
- Hong, S.; Seo, P.J.; Yang, M.; Xiang, F.; Park, C. Exploring valid reference genes for gene expression studies in Brachypodium distachyonby real-time PCR. BMC Plant Biol. 2008, 8, 112. [Google Scholar] [CrossRef]
- Liang, W.; Zou, X.; Carballar-Lejarazú, R.; Wu, L.; Sun, W.; Yuan, X.; Wu, S.; Li, P.; Ding, H.; Ni, L.; et al. Selection and evaluation of reference genes for qRT-PCR analysis in Euscaphis konishii Hayata based on transcriptome data. Plant Methods 2018, 14, 42. [Google Scholar] [CrossRef]
- Gu, C.; Liu, L.; Deng, Y.; Zhu, X.; Lu, X.; Huang, S. Validation of reference genes for RT-qPCR normalization in Iris. lactea var. chinensis leaves under different experimental conditions. Sci. Hortic. 2014, 175, 144–149. [Google Scholar] [CrossRef]
- Gu, C.; Liu, L.; Xu, C.; Zhao, Y.; Zhu, X.; Huang, S. Reference gene selection for quantitative real-Time RT-PCR normalization in Iris. lactea var. chinensis roots under cadmium, lead, and salt stress conditions. Sci. World J. 2014, 2014, 532713. [Google Scholar] [CrossRef]
- Fei, X.; Shi, Q.; Yang, T.; Fei, Z.; Wei, A. Expression Stabilities of Ten Candidate Reference Genes for RT-qPCR in Zanthoxylum bungeanum Maxim. Molecules 2018, 23, 802. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Zhang, P.; Lin, W.; Zheng, X.; Cai, M.; Peng, C. Sequencing of anthocyanin synthesis-related enzyme genes and screening of reference genes in leaves of four dominant subtropical forest tree species. Gene 2019, 716, 144024. [Google Scholar] [CrossRef] [PubMed]
- Yoshihara, N.; Imayama, T.; Matsuo, Y.; Fukuchi-Mizutani, M.; Tanaka, Y.; Ino, I.; Yabuya, T. Characterization of cDNA clones encoding anthocyanin 3-p-coumaroyltransferase from Iris hollandica. Plant Sci. 2006, 171, 632–639. [Google Scholar] [CrossRef]
- Yoshihara, N.; Imayama, T.; Fukuchi-Mizutani, M.; Okuhara, H.; Tanaka, Y.; Ino, I.; Yabuya, T. cDNA cloning and characterization of UDP-glucose: Anthocyanidin 3-O-glucosyltransferase in Iris hollandica. Plant Sci. 2005, 169, 496–501. [Google Scholar] [CrossRef]
- Chen, C.; Wu, J.; Hua, Q.; Tel-Zur, N.; Xie, F.; Zhang, Z.; Chen, J.; Zhang, R.; Hu, G.; Zhao, J.; et al. Identification of reliable reference genes for quantitative real-time PCR normalization in pitaya. Plant Methods 2019, 15, 70. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Han, S.; Yin, W.; Xia, X.; Liu, C. Comparison of reliable reference genes following different hormone treatments by various algorithms for qRT-PCR analysis of Metasequoia. Int. J. Mol. Sci. 2019, 20, 34. [Google Scholar] [CrossRef] [PubMed]
- Gantasala, N.P.; Papolu, P.K.; Thakur, P.K.; Kamaraju, D.; Sreevathsa, R.; Rao, U. Selection and validation of reference genes for quantitative gene expression studies by real-time PCR in eggplant (Solanum melongena L). BMC Res. Notes 2013, 6, 312. [Google Scholar] [CrossRef]
- Hou, F.; Li, S.; Wang, J.; Kang, X.; Weng, Y.; Xing, G. Identification and validation of reference genes for quantitative real-time PCR studies in long yellow daylily, Hemerocallis citrina Borani. PLoS ONE 2017, 12, e174933. [Google Scholar] [CrossRef]
- Li, X.; Tang, D.; Shi, Y. Selection of reference genes for quantitative real-time PCR normalization in Narcissus pseudonarcissus in different cultivars and different organs. Heliyon 2018, 4, e686. [Google Scholar] [CrossRef]
- De Spiegelaere, W.; Dern-Wieloch, J.; Weigel, R.; Schumacher, V.; Schorle, H.; Nettersheim, D.; Bergmann, M.; Brehm, R.; Kliesch, S.; Vandekerckhove, L.; et al. Reference Gene Validation for RT-qPCR, a Note on Different Available Software Packages. PLoS ONE 2015, 10, e122515. [Google Scholar] [CrossRef]
- Desai, A.; Mitchison, T.J. Microtubule polymerization dynamics. Annu. Rev. Cell Dev. Biol. 1997, 13, 83–117. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Zhou, X.R.; Pang, B.P. Reference gene selection and evaluation for expression analysis using qRT-PCR in Galeruca daurica (Joannis). Bull. Entomol. Res. 2017, 107, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Yang, Z.; Sun, X.; Zhang, L.; Fang, Z. Selection of reference genes for quantitative real-time PCR during flower bud development in CMS7311 of heading Chinese cabbage (Brassica rapa L. ssp. pekinensis). Acta Physiol. Plant. 2014, 36, 809–814. [Google Scholar] [CrossRef]
- Chang, E.; Shi, S.; Liu, J.; Cheng, T.; Xue, L.; Yang, X.; Yang, W.; Lan, Q.; Jiang, Z. Selection of reference genes for quantitative gene expression studies in Platycladus orientalis (Cupressaceae) using real-time PCR. PLoS ONE 2012, 7, e33278. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Ohmiya, A. Seeing is believing: Engineering anthocyanin and carotenoid biosynthetic pathways. Curr. Opin. Biotech. 2008, 19, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Lou, Q.; Liu, Y.; Qi, Y.; Jiao, S.; Tian, F.; Jiang, L.; Wang, Y. Transcriptome sequencing and metabolite analysis reveals the role of delphinidin metabolism in flower colour in grape hyacinth. J. Exp. Bot. 2014, 65, 3157–3164. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.; Bian, X.; Jia, Z.; Guo, X.; Xie, Y. De novo sequencing and comprehensive analysis of the mutant transcriptome from purple sweet potato (Ipomoea batatas L.). Gene 2016, 575, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Sagawa, J.M.; Di Stilio, V.S.; Bradshaw, H.D. Bulk segregant analysis of an induced floral mutant identifies a MIXTA-Like R2R3MYB controlling nectar guide formation in Mimulus lewisii. Genetics 2013, 194, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Rocha, A.J.; Monteiro-Júnior, J.E.; Freire, J.E.C.; Sousa, A.J.S.; Fonteles, C.S.R. Real Time PCR: The use of reference genes and essential rules required to obtain normalisation data reliable to quantitative gene expression. J. Mol. Biol. Res. 2014, 5, 45. [Google Scholar] [CrossRef]
- Chen, D.; Pan, X.; Xiao, P.; Farwell, M.A.; Zhang, B. Evaluation and identification of reliable reference genes for pharmacogenomics, toxicogenomics, and small RNA expression analysis. J. Cell. Physiol. 2011, 226, 2469–2477. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Flavonoids Content (mg/g) | Carotenoids Content (mg/g) |
|---|---|---|
| S1_Pew | 7.746 ± 0.3217 a | 0.030 ± 0.0022 d |
| S2_Pew | 5.101 ± 0.6136 b | 0.110 ± 0.0069 c |
| S3_Pew | 3.592 ± 0.2200 c | 0.112 ± 0.0060 c |
| S1_Pe | 7.845 ± 0.3986 a | 0.035 ± 0.0029 d |
| S2_Pe | 5.661 ± 0.3800 b | 0.149 ± 0.0043 a |
| S3_Pe | 3.045 ± 0.5477 c | 0.136 ± 0.0119 b |
| Gene | K | E (%) | R2 | Product Length (bp) |
|---|---|---|---|---|
| Ci18S | −3.50 | 93.07 | 0.9979 | 102 |
| CiACT | −3.14 | 108.2 | 0.9924 | 93 |
| CiEF1α | −3.57 | 90.60 | 0.9978 | 119 |
| CiEIF3 | −3.51 | 92.71 | 0.9879 | 92 |
| CiGAPDH | −3.55 | 91.29 | 0.9925 | 140 |
| CiTUA | −3.09 | 110.68 | 0.9952 | 146 |
| CiTUB | −3.43 | 95.68 | 0.9924 | 81 |
| CiUBQ | −3.22 | 104.44 | 0.9911 | 117 |
| Sample | Ci18S | CiACT | CiEF1α | CiEIF3 | CiGAPDH | CiTUA | CiTUB | CiUBQ | |
|---|---|---|---|---|---|---|---|---|---|
| Se | average value | 17.24 | 23.3 | 22.8 | 26.39 | 23.51 | 27.23 | 25.73 | 26.26 |
| SD | 0.65 | 0.94 | 1.33 | 1.3 | 1.33 | 1.05 | 0.56 | 1.43 | |
| CV (%) | 3.76 | 4.04 | 5.84 | 4.92 | 5.64 | 3.87 | 2.17 | 5.44 | |
| Pew | average value | 16.66 | 19.71 | 20.05 | 24.35 | 20.59 | 25 | 22.03 | 23.57 |
| SD | 0.54 | 1.37 | 1.08 | 1.23 | 1.23 | 1.54 | 0.56 | 1.04 | |
| CV (%) | 3.25 | 6.96 | 5.4 | 5.06 | 5.95 | 6.14 | 2.55 | 4.43 | |
| Pe | average value | 16.22 | 19.3 | 19.25 | 21.3 | 15.38 | 23.05 | 19.35 | 19.78 |
| SD | 0.91 | 1.01 | 1.43 | 1.28 | 0.85 | 1.79 | 0.78 | 0.7 | |
| CV (%) | 5.6 | 5.24 | 7.44 | 6.03 | 5.53 | 7.76 | 4.04 | 3.53 | |
| St | average value | 16.33 | 20.61 | 19.39 | 23.7 | 19.24 | 21.09 | 23 | 22.27 |
| SD | 0.86 | 1.28 | 0.95 | 0.96 | 0.49 | 1.56 | 1.13 | 0.76 | |
| CV (%) | 5.29 | 6.19 | 4.88 | 4.05 | 2.53 | 7.41 | 4.91 | 3.43 | |
| Pi | average value | 16.96 | 21.28 | 20.37 | 25.04 | 21.3 | 25.46 | 22.74 | 23.11 |
| SD | 0.68 | 1.95 | 0.96 | 1.38 | 1.1 | 0.7 | 1.09 | 1.07 | |
| CV (%) | 3.98 | 9.16 | 4.73 | 5.52 | 5.16 | 2.75 | 4.79 | 4.62 |
| Rank | Se | Pew | Pe | St | Pi | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Gene | SV | Gene | SV | Gene | SV | Gene | SV | Gene | SV | |
| 1 | CiTUB | 0.16 | CiTUB | 0.23 | CiTUB | 0.12 | CiTUA | 0.16 | CiTUB | 0.11 |
| 2 | Ci18S | 0.25 | Ci18S | 0.26 | CiEIF3 | 0.19 | CiEIF3 | 0.19 | CiEF1α | 0.12 |
| 3 | CiGAPDH | 0.25 | CiEF1α | 0.27 | Ci18S | 0.21 | CiEF1α | 0.22 | CiEIF3 | 0.14 |
| 4 | CiEF1α | 0.30 | CiEIF3 | 0.29 | CiACT | 0.22 | CiUBQ | 0.25 | CiUBQ | 0.15 |
| 5 | CiACT | 0.30 | CiUBQ | 0.34 | CiGAPDH | 0.24 | CiGAPDH | 0.26 | CiACT | 0.23 |
| 6 | CiTUA | 0.34 | CiACT | 0.34 | CiUBQ | 0.26 | CiTUB | 0.26 | CiTUA | 0.28 |
| 7 | CiEIF3 | 0.36 | CiTUA | 0.35 | CiEF1α | 0.34 | CiACT | 0.27 | CiGAPDH | 0.36 |
| 8 | CiUBQ | 0.37 | CiGAPDH | 0.36 | CiTUA | 0.43 | Ci18S | 0.27 | Ci18S | 0.37 |
| Sample | Rank | Gene | Min | Max | SD (±Cq) | CV (%Cq) |
|---|---|---|---|---|---|---|
| Pe | 1 | CiUBQ | 18.96 | 20.94 | 0.53 | 2.68 |
| 2 | CiTUB | 18.27 | 20.54 | 0.63 | 3.25 | |
| 3 | CiGAPDH | 14.24 | 16.76 | 0.66 | 4.29 | |
| 4 | Ci18S | 14.73 | 16.97 | 0.77 | 4.75 | |
| 5 | CiEF1α | 18.12 | 20.63 | 0.80 | 4.08 | |
| 6 | CiACT | 18.30 | 20.95 | 0.82 | 4.25 | |
| 7 | CiEIF3 | 19.18 | 22.92 | 1.01 | 4.75 | |
| 8 | CiTUA | 21.06 | 25.20 | 1.30 | 5.57 | |
| Pew | 1 | CiTUB | 21.25 | 22.71 | 0.44 | 1.99 |
| 2 | Ci18S | 15.84 | 17.46 | 0.47 | 2.80 | |
| 3 | CiUBQ | 21.65 | 24.90 | 0.79 | 3.34 | |
| 4 | CiTUA | 24.13 | 26.80 | 0.83 | 3.26 | |
| 5 | CiEF1α | 18.77 | 21.94 | 0.88 | 4.38 | |
| 6 | CiGAPDH | 18.97 | 22.51 | 0.98 | 4.76 | |
| 7 | CiEIF3 | 22.81 | 26.17 | 1.03 | 4.21 | |
| 8 | CiACT | 17.90 | 21.84 | 1.03 | 5.24 | |
| St | 1 | CiGAPDH | 18.56 | 20.01 | 0.39 | 2.04 |
| 2 | CiUBQ | 20.99 | 23.48 | 0.63 | 2.81 | |
| 3 | Ci18S | 14.90 | 17.75 | 0.68 | 4.17 | |
| 4 | CiTUA | 18.88 | 22.62 | 0.70 | 3.35 | |
| 5 | CiEF1α | 18.07 | 21.23 | 0.70 | 3.63 | |
| 6 | CiEIF3 | 22.17 | 25.09 | 0.73 | 3.08 | |
| 7 | CiTUB | 21.17 | 24.32 | 0.94 | 4.11 | |
| 8 | CiACT | 19.03 | 22.26 | 1.12 | 5.41 | |
| Pi | 1 | CiTUA | 24.51 | 26.25 | 0.56 | 2.19 |
| 2 | Ci18S | 16.04 | 17.85 | 0.57 | 3.34 | |
| 3 | CiEF1α | 19.03 | 21.89 | 0.81 | 3.96 | |
| 4 | CiTUB | 21.58 | 24.74 | 0.89 | 3.90 | |
| 5 | CiUBQ | 22.04 | 24.73 | 0.91 | 3.95 | |
| 6 | CiGAPDH | 19.63 | 22.64 | 0.93 | 4.36 | |
| 7 | CiEIF3 | 23.69 | 26.92 | 1.20 | 4.78 | |
| 8 | CiACT | 19.10 | 23.86 | 1.66 | 7.81 | |
| Se | 1 | CiTUB | 25.01 | 26.79 | 0.44 | 1.71 |
| 2 | Ci18S | 16.24 | 18.20 | 0.52 | 3.03 | |
| 3 | CiTUA | 26.34 | 28.91 | 0.68 | 2.47 | |
| 4 | CiACT | 22.22 | 24.91 | 0.78 | 3.36 | |
| 5 | CiGAPDH | 21.72 | 26.09 | 0.99 | 4.23 | |
| 6 | CiEF1α | 20.19 | 24.45 | 1.03 | 4.53 | |
| 7 | CiEIF3 | 24.32 | 27.99 | 1.09 | 4.14 | |
| 8 | CiUBQ | 24.13 | 27.86 | 1.24 | 4.73 |
| Rank | Se | Pew | Pe | St | Pi | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Gene | Geomean of Ranking Values | Gene | Geomean of Ranking Values | Gene | Geomean of Ranking Values | Gene | Geomean of Ranking Values | Gene | Geomean of Ranking Values | |
| 1 | CiTUB | 1.00 | CiTUB | 1.71 | CiTUB | 1.26 | CiTUA | 1.59 | CiTUB | 1.59 |
| 2 | Ci18S | 1.59 | CiEF1a | 2.47 | Ci18S | 2.29 | CiEF1a | 2.47 | CiUBQ | 2.71 |
| 3 | CiGAPDH | 3.91 | Ci18S | 2.88 | CiUBQ | 3.30 | CiGAPDH | 3.11 | CiEF1α | 2.88 |
| 4 | CiEF1a | 4.16 | CiACT | 3.63 | CiGAPDH | 3.56 | CiUBQ | 3.17 | CiTUA | 3.11 |
| 5 | CiACT | 4.64 | CiUBQ | 3.91 | CiEIF3 | 3.83 | CiEIF3 | 3.30 | CiEIF3 | 3.98 |
| 6 | CiTUA | 4.76 | CiEIF3 | 4.38 | CiACT | 5.52 | Ci18S | 5.52 | Ci18S | 4.58 |
| 7 | CiEIF3 | 7.00 | CiTUA | 5.81 | CiEF1a | 5.59 | CiTUB | 5.94 | CiGAPDH | 6.65 |
| 8 | CiUBQ | 8.00 | CiGAPDH | 7.27 | CiTUA | 8.00 | CiACT | 7.65 | CiACT | 6.84 |
| Gene | Gene ID | Primer Sequences (5′–3′) | Purpose |
|---|---|---|---|
| Ci18S | JinHuaCha00367404 | F: CGTTCGTCTGGCTTCTTAGTCCTTC | Reference gene |
| R: AACTCGCACAAACCAAACACAACTC | |||
| CiACT | JinHuaCha00305809 | F: CTCTCGTCTTCTCCGTCTCCTCAC | |
| R: AGCCTTCACCATTCCAGTTCCATTG | |||
| CiEF1α | JinHuaCha00372976 | F: TCGATTGCCACACTTCCCACATTG | |
| R: CCCAGCGTCACCGTTCTTCAAG | |||
| CiEIF3 | JinHuaCha00379840 | F: ACCGGCTTATGCGTTATGCTCATC | |
| R: TGGTTCATGGCTGCTGTATGTCAC | |||
| CiGAPDH | JinHuaCha00022420 | F: AGCAAGGACTGGAGAGGTGGAAG | |
| R: TCAACAGTGGGAACACGGAAAGC | |||
| CiTUA | JinHuaCha00361094 | F: GACTGTTGGAGGAGGTGATGATGC | |
| R: GGTGGAAGAGTTGGCGGTATGTTC | |||
| CiTUB | JinHuaCha00329993 | F: AGTTGAGAACGCCGATGAGTGTATG | |
| R: GTGGTGAGCTTGAGTGTACGGAAG | |||
| CiUBQ | JinHuaCha00084829 | F: TGCAGAAGGACCCTCCCACATC | |
| R: CCAGAAATACGCCTCCAGCATACG | |||
| CiCHS | JinHuaCha00343068 | F: TCCCAGATAGTGACGGTGCCATC | Flower pigment synthesis-related genes |
| R: GTTCCAATCAGAGATGCCCAAGGG | |||
| CiFLS | JinHuaCha00037405 | F: ACCAGCAATCACCACCGTCAAAG | |
| R: CAGCCTCCTCCACCATCCTCAC | |||
| CiF3′H | JinHuaCha00381658 | F: ATCTGCTCCGTCCATCTCTTCTCC | |
| R: CTAGGTTCACTGCTGCCGCTTG | |||
| CiF3H | JinHuaCha00051595 | F: TGGAGGTGTTGTCTGAGGCTATGG | |
| R: GAGGTCGGGTTGTGGGCATTTC | |||
| CiBCH | JinHuaCha00348661 | F: ACGAAGAGGAGGGTGAGCAAGAG | |
| R: CTAGACATGACTGCGGCGACAAG | |||
| CiNSY | JinHuaCha00334476 | F: CGGTCCTGGCTGATGTCATTGC | |
| R: CCACAACCCATTCCACACCCTATG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, P.; Chen, S.; Chen, S.; Zhu, Y.; Lin, Y.; Xu, X.; Liu, Z.; Zou, S. Selection and Validation of qRT-PCR Internal Reference Genes to Study Flower Color Formation in Camellia impressinervis. Int. J. Mol. Sci. 2024, 25, 3029. https://doi.org/10.3390/ijms25053029
Zhang P, Chen S, Chen S, Zhu Y, Lin Y, Xu X, Liu Z, Zou S. Selection and Validation of qRT-PCR Internal Reference Genes to Study Flower Color Formation in Camellia impressinervis. International Journal of Molecular Sciences. 2024; 25(5):3029. https://doi.org/10.3390/ijms25053029
Chicago/Turabian StyleZhang, Peilan, Shuying Chen, Siyu Chen, Yuanming Zhu, Yuqing Lin, Xinyu Xu, Zhongjian Liu, and Shuangquan Zou. 2024. "Selection and Validation of qRT-PCR Internal Reference Genes to Study Flower Color Formation in Camellia impressinervis" International Journal of Molecular Sciences 25, no. 5: 3029. https://doi.org/10.3390/ijms25053029
APA StyleZhang, P., Chen, S., Chen, S., Zhu, Y., Lin, Y., Xu, X., Liu, Z., & Zou, S. (2024). Selection and Validation of qRT-PCR Internal Reference Genes to Study Flower Color Formation in Camellia impressinervis. International Journal of Molecular Sciences, 25(5), 3029. https://doi.org/10.3390/ijms25053029