
Platelet-Activating Factor Receptor (PAFR) Regulates Retinal Progenitor/Stem Cells Profile in Ciliary Epithelium Cells

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
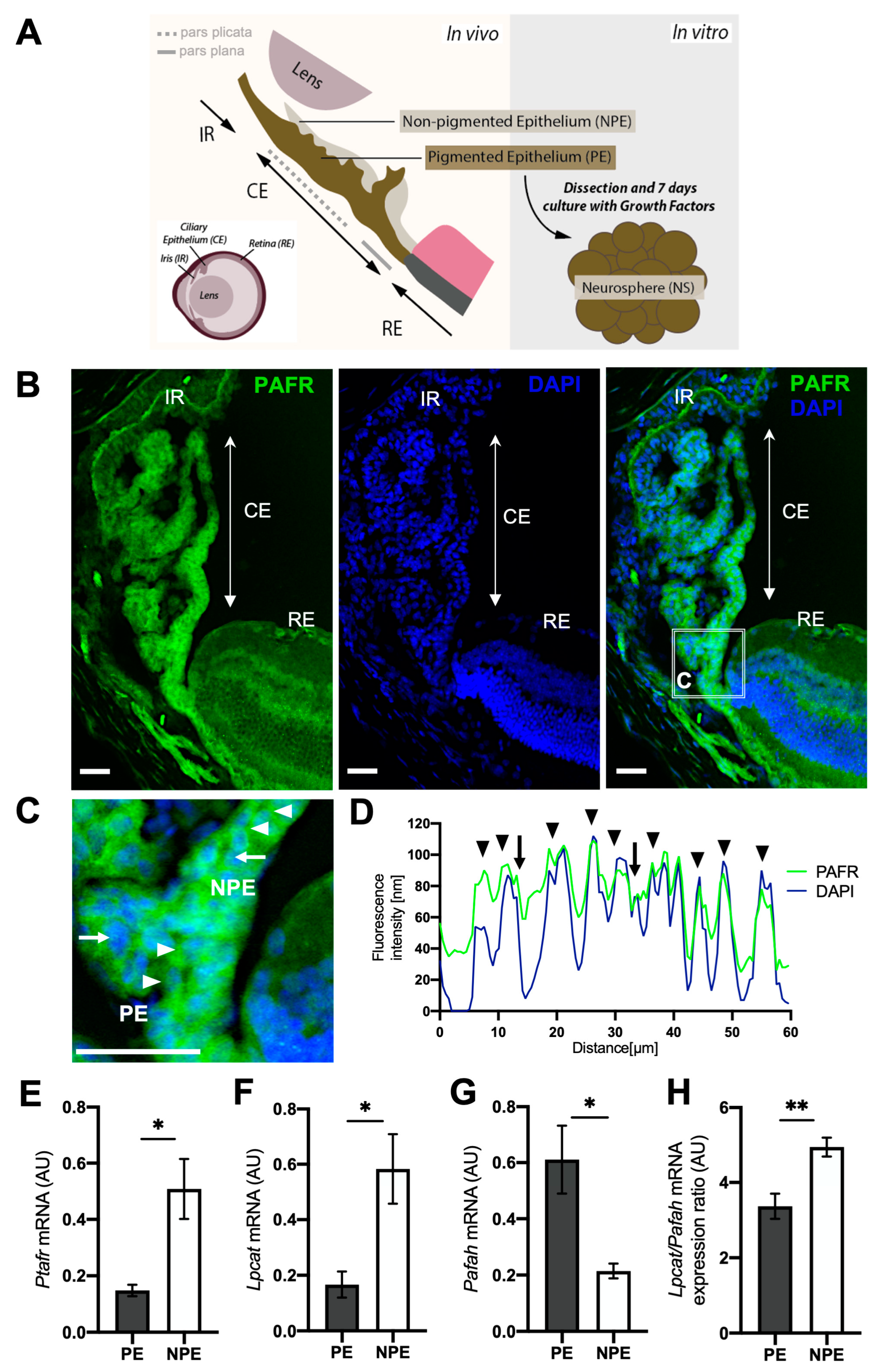
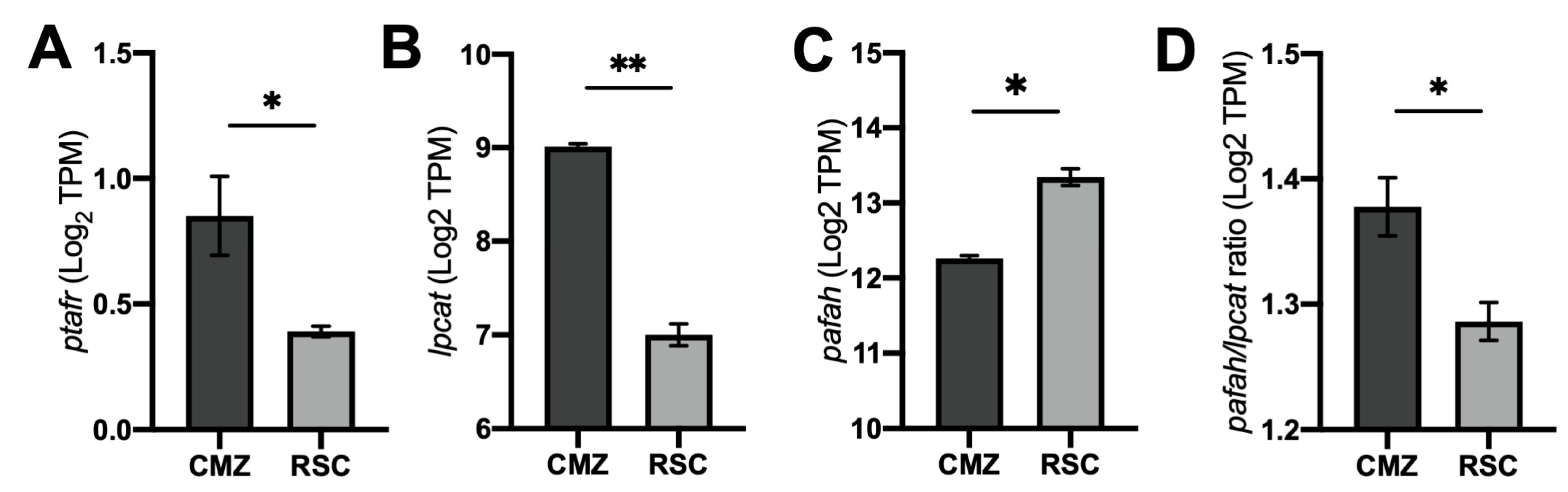
2.1. Differential Expression of PAF Receptor (PAFR) and Associated Pathway Enzymes in Ciliary Epithelium
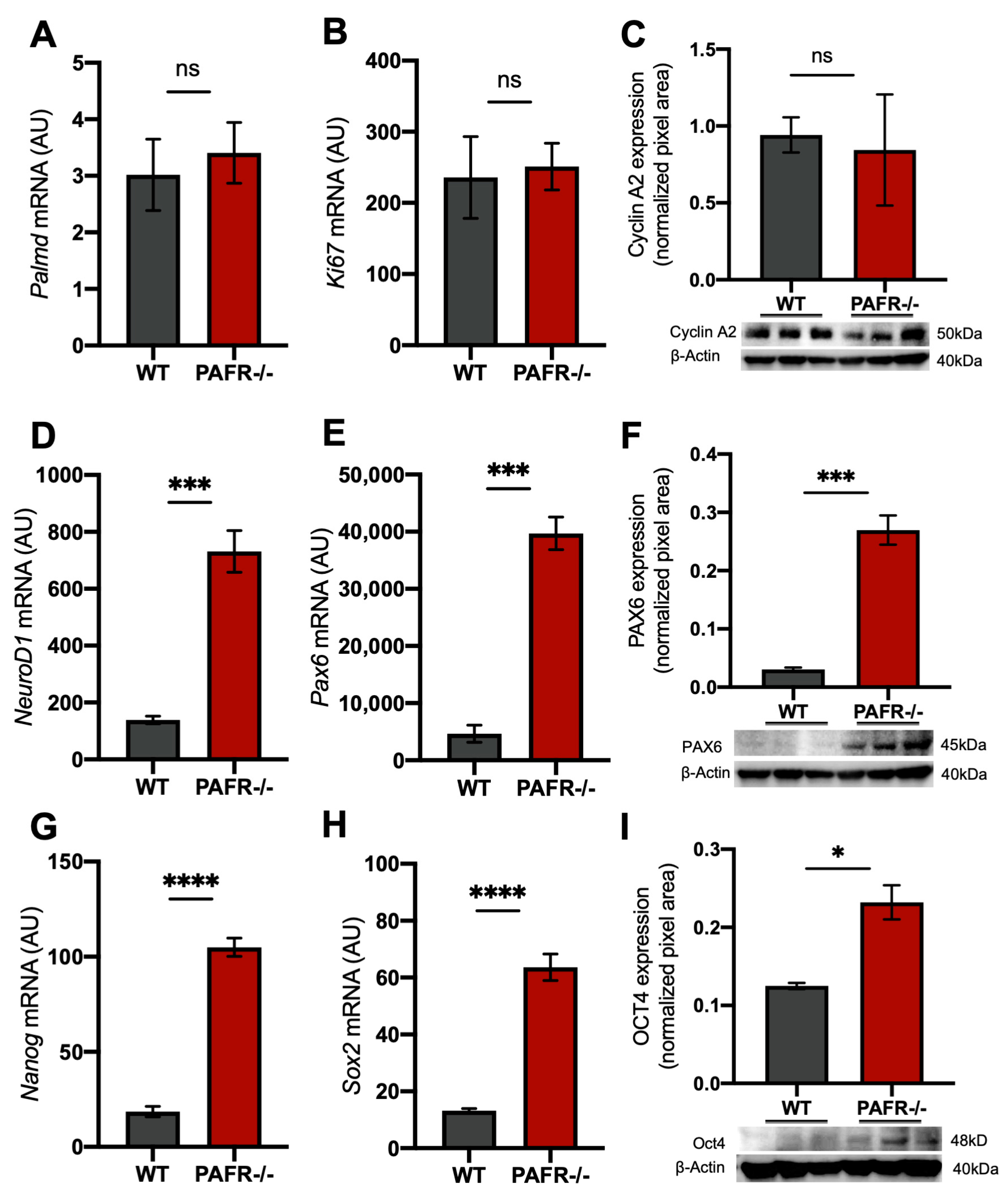
2.2. PAFR Expression in Quiescent Retinal Progenitor/Stem Cells
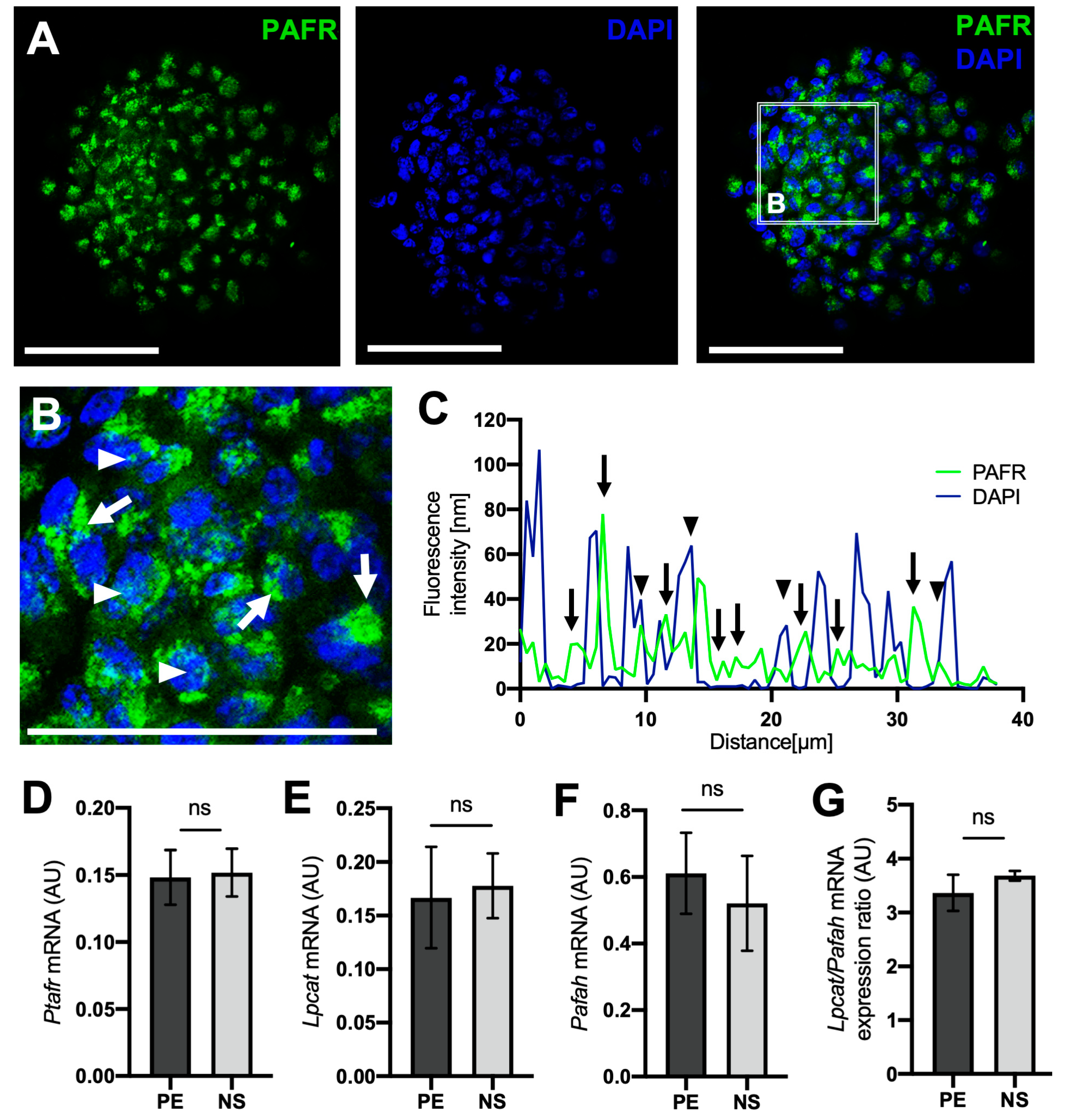
2.3. PAFR and PAF-Related Enzyme Expression Is Similar in PE and PE-Derived Neurospheres
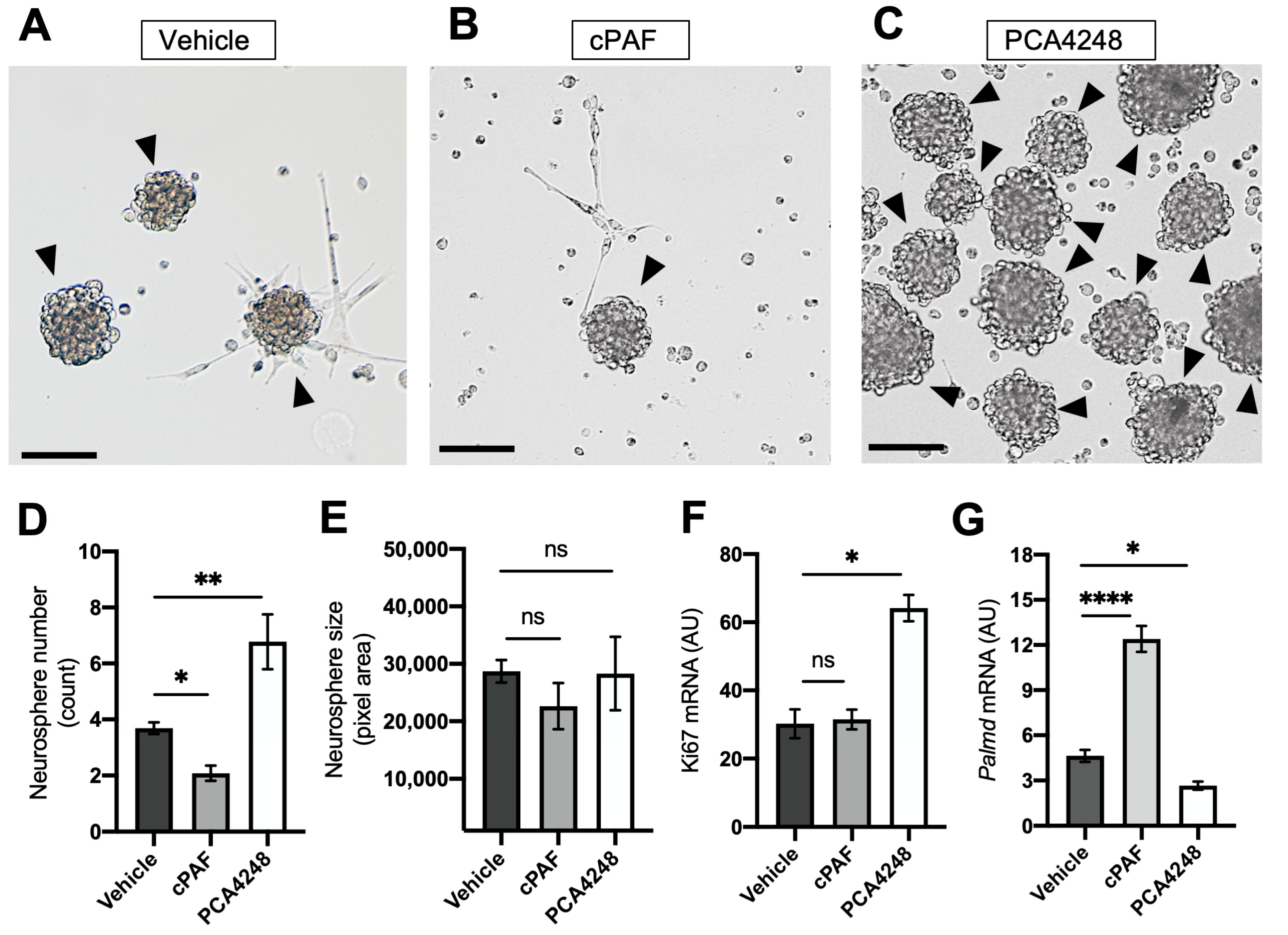
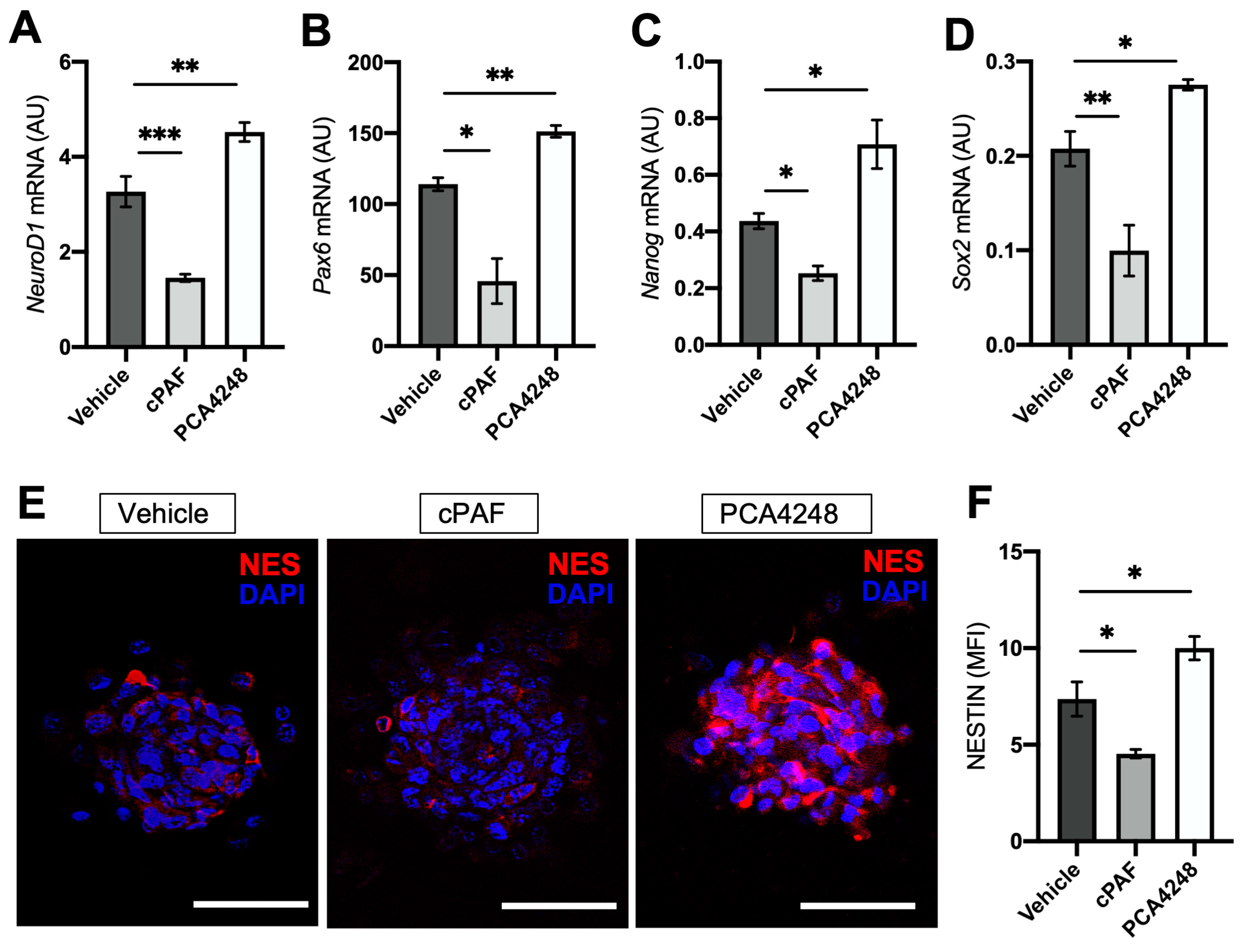
2.4. PAFR Pharmacological Modulation-Regulated Neurosphere Formation and Retinal Stem Cell Marker Expression
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. CE Tissue Dissection and Neurosphere Formation Assay
4.3. mRNA Analysis
4.4. PAFR Agonist and Antagonist Treatment and Cytotoxicity Assay
4.5. Immunofluorescence
4.6. Western Blot (WB)
4.7. Gene Expression Datasets
4.8. Samples and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fischer, A.J.; Bosse, J.L.; El-Hodiri, H.M. The ciliary marginal zone (CMZ) in development and regeneration of the vertebrate eye. Exp. Eye Res. 2013, 116, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Tang, L.; Pham, H. Identification of neural progenitors in the adult mammalian eye. Biochem. Biophys. Res. Commun. 2000, 270, 517–521. [Google Scholar] [CrossRef] [PubMed]
- Tropepe, V.; Coles, B.L.; Chiasson, B.J.; Horsford, D.J.; Elia, A.J.; McInnes, R.R.; van der Kooy, D. Retinal stem cells in the adult mammalian eye. Science 2000, 287, 2032–2036. [Google Scholar] [CrossRef]
- McLaughlin, C.W.; Zellhuber-McMillan, S.; Macknight, A.D.; Civan, M.M. Electron microprobe analysis of ouabain-exposed ciliary epithelium: PE-NPE cell couplets form the functional units. Am. J. Physiol. Cell. Physiol. 2004, 286, C1376–C1389. [Google Scholar] [CrossRef]
- Li, S.K.; Shan, S.W.; Li, H.L.; Cheng, A.K.; Pan, F.; Yip, S.P.; Civan, M.M.; To, C.H.; Do, C.W. Characterization and Regulation of Gap Junctions in Porcine Ciliary Epithelium. Investig. Ophthalmol. Vis. Sci. 2018, 59, 3461–3468. [Google Scholar] [CrossRef]
- Abdouh, M.; Bernier, G. In vivo reactivation of a quiescent cell population located in the ocular ciliary body of adult mammals. Exp. Eye Res. 2006, 83, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Engelhardt, M.; Bogdahn, U.; Aigner, L. Adult retinal pigment epithelium cells express neural progenitor properties and the neuronal precursor protein doublecortin. Brain Res. 2005, 1040, 98–111. [Google Scholar] [CrossRef]
- Lord-Grignon, J.; Abdouh, M.; Bernier, G. Identification of genes expressed in retinal progenitor/stem cell colonies isolated from the ocular ciliary body of adult mice. Gene Expr. Patterns 2006, 6, 992–999. [Google Scholar] [CrossRef]
- Del Debbio, C.B.; Peng, X.; Xiong, H.; Ahmad, I. Adult ciliary epithelial stem cells generate functional neurons and differentiate into both early and late born retinal neurons under non-cell autonomous influences. BMC Neurosci. 2013, 14, 130. [Google Scholar] [CrossRef]
- Cicero, S.A.; Johnson, D.; Reyntjens, S.; Frase, S.; Connell, S.; Chow, L.M.; Baker, S.J.; Sorrentino, B.P.; Dyer, M.A. Cells previously identified as retinal stem cells are pigmented ciliary epithelial cells. Proc. Natl. Acad. Sci. USA 2009, 106, 6685–6690. [Google Scholar] [CrossRef]
- Prescott, S.M.; Zimmerman, G.A.; Stafforini, D.M.; McIntyre, T.M. Platelet-activating factor and related lipid mediators. Annu. Rev. Biochem. 2000, 69, 419–445. [Google Scholar] [CrossRef] [PubMed]
- Harayama, T.; Shindou, H.; Ogasawara, R.; Suwabe, A.; Shimizu, T. Identification of a novel noninflammatory biosynthetic pathway of platelet-activating factor. J. Biol. Chem. 2008, 283, 11097–11106. [Google Scholar] [CrossRef] [PubMed]
- Snyder, F. CDP-choline:alkylacetylglycerol cholinephosphotransferase catalyzes the final step in the de novo synthesis of platelet-activating factor. Biochim. Biophys. Acta 1997, 1348, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Snyder, F.; Fitzgerald, V.; Blank, M.L. Biosynthesis of platelet-activating factor and enzyme inhibitors. Adv. Exp. Med. Biol. 1996, 416, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Prescott, S.M. A thematic series on phospholipases. J. Biol. Chem. 1997, 272, 15043. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yang, L.; Foulks, J.M.; Weyrich, A.S.; Marathe, G.K.; McIntyre, T.M. Intracellular PAF catabolism by PAF acetylhydrolase counteracts continual PAF synthesis. J. Lipid Res. 2007, 48, 2365–2376. [Google Scholar] [CrossRef] [PubMed]
- Honda, Z.; Ishii, S.; Shimizu, T. Platelet-activating factor receptor. J. Biochem. 2002, 131, 773–779. [Google Scholar] [CrossRef]
- Ishii, S.; Shimizu, T. Platelet-activating factor (PAF) receptor and genetically engineered PAF receptor mutant mice. Prog. Lipid Res. 2000, 39, 41–82. [Google Scholar] [CrossRef]
- Choi, H.Y.; Saha, S.K.; Kim, K.; Kim, S.; Yang, G.M.; Kim, B.; Kim, J.H.; Cho, S.G. G protein-coupled receptors in stem cell maintenance and somatic reprogramming to pluripotent or cancer stem cells. BMB Rep. 2015, 48, 68–80. [Google Scholar] [CrossRef]
- Bussolino, F.; Gremo, F.; Tetta, C.; Pescarmona, G.P.; Camussi, G. Production of platelet-activating factor by chick retina. J. Biol. Chem. 1986, 261, 16502–16508. [Google Scholar] [CrossRef]
- Fragel-Madeira, L.; Meletti, T.; Mariante, R.M.; Monteiro, R.Q.; Einicker-Lamas, M.; Bernardo, R.R.; Lopes, A.H.; Linden, R. Platelet activating factor blocks interkinetic nuclear migration in retinal progenitors through an arrest of the cell cycle at the S/G2 transition. PLoS ONE 2011, 6, e16058. [Google Scholar] [CrossRef]
- Mori, M.; Aihara, M.; Shimizu, T. Localization of platelet-activating factor receptor messenger RNA in the rat eye. Investig. Ophthalmol. Vis. Sci. 1997, 38, 2672–2678. [Google Scholar]
- Xu, B.; Tang, X.; Jin, M.; Zhang, H.; Du, L.; Yu, S.; He, J. Unifying developmental programs for embryonic and postembryonic neurogenesis in the zebrafish retina. Development 2020, 147, dev185660. [Google Scholar] [CrossRef] [PubMed]
- Chistyakov, D.V.; Baksheeva, V.E.; Tiulina, V.V.; Goriainov, S.V.; Azbukina, N.V.; Gancharova, O.S.; Arifulin, E.A.; Komarov, S.V.; Chistyakov, V.V.; Tikhomirova, N.K.; et al. Mechanisms and Treatment of Light-Induced Retinal Degeneration-Associated Inflammation: Insights from Biochemical Profiling of the Aqueous Humor. Int. J. Mol. Sci. 2020, 21, 704. [Google Scholar] [CrossRef] [PubMed]
- Aladdad, A.M.; Kador, K.E. Adult Stem Cells, Tools for Repairing the Retina. Curr. Ophthalmol. Rep. 2019, 7, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.B.; Gao, M.L.; Deng, W.L.; Wu, K.C.; Sugita, S.; Mandai, M.; Takahashi, M. Stemming retinal regeneration with pluripotent stem cells. Prog. Retin. Eye Res. 2019, 69, 38–56. [Google Scholar] [CrossRef] [PubMed]
- Maeda, A.; Mandai, M.; Takahashi, M. Gene and Induced Pluripotent Stem Cell Therapy for Retinal Diseases. Annu. Rev. Genom. Hum. Genet. 2019, 20, 201–216. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.; Oh, I.H. Regeneration of the retina: Toward stem cell therapy for degenerative retinal diseases. BMB Rep. 2015, 48, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gao, Z.; Zhang, Y.; Feng, S.Q.; Liu, Y.; Shields, L.B.E.; Zhao, Y.Z.; Zhu, Q.; Gozal, D.; Shields, C.B.; et al. Attenuated Reactive Gliosis and Enhanced Functional Recovery Following Spinal Cord Injury in Null Mutant Mice of Platelet-Activating Factor Receptor. Mol. Neurobiol. 2016, 53, 3448–3461. [Google Scholar] [CrossRef] [PubMed]
- Hattori, M.; Arai, H. Intracellular PAF-Acetylhydrolase Type I. Enzymes 2015, 38, 23–36. [Google Scholar] [CrossRef]
- Hines, T.J.; Gao, X.; Sahu, S.; Lange, M.M.; Turner, J.R.; Twiss, J.L.; Smith, D.S. An Essential Postdevelopmental Role for Lis1 in Mice. eNeuro 2018, 5, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Jheng, G.W.; Hur, S.S.; Chang, C.M.; Wu, C.C.; Cheng, J.S.; Lee, H.H.; Chung, B.C.; Wang, Y.K.; Lin, K.H.; Del Álamo, J.C.; et al. Lis1 dysfunction leads to traction force reduction and cytoskeletal disorganization during cell migration. Biochem. Biophys. Res. Commun. 2018, 497, 869–875. [Google Scholar] [CrossRef] [PubMed]
- Lanctot, A.A.; Peng, C.Y.; Pawlisz, A.S.; Joksimovic, M.; Feng, Y. Spatially dependent dynamic MAPK modulation by the Nde1-Lis1-Brap complex patterns mammalian CNS. Dev. Cell 2013, 25, 241–255. [Google Scholar] [CrossRef]
- Livnat, I.; Finkelshtein, D.; Ghosh, I.; Arai, H.; Reiner, O. PAF-AH Catalytic Subunits Modulate the Wnt Pathway in Developing GABAergic Neurons. Front. Cell. Neurosci. 2010, 4, 19. [Google Scholar] [CrossRef]
- Manya, H.; Aoki, J.; Watanabe, M.; Adachi, T.; Asou, H.; Inoue, Y.; Arai, H.; Inoue, K. Switching of platelet-activating factor acetylhydrolase catalytic subunits in developing rat brain. J. Biol. Chem. 1998, 273, 18567–18572. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Zhao, R.; Yao, L.; Xu, C.; Cong, Q.; Jiang, W. Platelet-activating factor induces the stemness of ovarian cancer cells via the PAF/PAFR signaling pathway. Am. J. Transl. Res. 2020, 12, 7249–7261. [Google Scholar] [PubMed]
- Yu, Y.; Zhang, X.; Hong, S.; Zhang, M.; Cai, Q.; Jiang, W.; Xu, C. Epidermal growth factor induces platelet-activating factor production through receptors transactivation and cytosolic phospholipase A2 in ovarian cancer cells. J. Ovarian Res. 2014, 7, 39. [Google Scholar] [CrossRef]
- Bhosle, V.K.; Rivera, J.C.; Zhou, T.E.; Omri, S.; Sanchez, M.; Hamel, D.; Zhu, T.; Rouget, R.; Rabea, A.A.; Hou, X.; et al. Nuclear localization of platelet-activating factor receptor controls retinal neovascularization. Cell Discov. 2016, 2, 16017. [Google Scholar] [CrossRef]
- Kume, K.; Shimizu, T. Platelet-activating factor (PAF) induces growth stimulation, inhibition, and suppression of oncogenic transformation in NRK cells overexpressing the PAF receptor. J. Biol. Chem. 1997, 272, 22898–22904. [Google Scholar] [CrossRef]
- Ahmad, I.; Dooley, C.M.; Thoreson, W.B.; Rogers, J.A.; Afiat, S. In vitro analysis of a mammalian retinal progenitor that gives rise to neurons and glia. Brain Res. 1999, 831, 1–10. [Google Scholar] [CrossRef]
- Knezevic, I.I.; Predescu, S.A.; Neamu, R.F.; Gorovoy, M.S.; Knezevic, N.M.; Easington, C.; Malik, A.B.; Predescu, D.N. Tiam1 and Rac1 are required for platelet-activating factor-induced endothelial junctional disassembly and increase in vascular permeability. J. Biol. Chem. 2009, 284, 5381–5394. [Google Scholar] [CrossRef]
- Xu, C.; Wu, X.; Lu, M.; Tang, L.; Yao, H.; Wang, J.; Ji, X.; Hussain, M.; Wu, J. Protein tyrosine phosphatase 11 acts through RhoA/ROCK to regulate eosinophil accumulation in the allergic airway. FASEB J. 2019, 33, 11706–11720. [Google Scholar] [CrossRef]
- Del Debbio, C.B.; Santos, M.F.; Yan, C.Y.; Ahmad, I.; Hamassaki, D.E. Rho GTPases control ciliary epithelium cells proliferation and progenitor profile induction in vivo. Investig. Ophthalmol. Vis. Sci. 2014, 55, 2631–2641. [Google Scholar] [CrossRef] [PubMed]
- Dalmaso, B.; da Silva-Junior, I.A.; Fragel-Madeira, L.; Jancar, S.; Del Debbio, C.B. Platelet activating factor in the eye: Physiological roles, diseases and future perspectives. Prostaglandins Other Lipid Mediat. 2020, 153, 106522. [Google Scholar] [CrossRef] [PubMed]
- Travers, J.B.; Rohan, J.G.; Sahu, R.P. New Insights Into the Pathologic Roles of the Platelet-Activating Factor System. Front. Endocrinol. 2021, 12, 624132. [Google Scholar] [CrossRef] [PubMed]
- Magner, E.; Sandoval-Sanchez, P.; Kramer, A.C.; Thummel, R.; Hitchcock, P.F.; Taylor, S.M. Disruption of miR-18a Alters Proliferation, Photoreceptor Replacement Kinetics, Inflammatory Signaling, and Microglia/Macrophage Numbers During Retinal Regeneration in Zebrafish. Mol. Neurobiol. 2022, 59, 2910–2931. [Google Scholar] [CrossRef] [PubMed]
- Iribarne, M.; Hyde, D.R. Different inflammation responses modulate Müller glia proliferation in the acute or chronically damaged zebrafish retina. Front. Cell Dev. Biol. 2022, 10, 892271. [Google Scholar] [CrossRef] [PubMed]
- Grabinski, S.E.; Parsana, D.; Perkins, B.D. Comparative analysis of transcriptional changes in zebrafish. Front. Mol. Neurosci. 2023, 16, 1148840. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Zhang, X.; Chen, Y.; Lin, Z.; Zhang, S.; Zhang, Z.; Cao, L.; Guan, H.; Lu, J.; Xu, H. Context-dependent effects of inflammation on retina regeneration. Mol. Neurobiol. 2022, 59, 4351–4367. [Google Scholar] [CrossRef]
- Grisé, K.N.; Bautista, N.X.; Jacques, K.; Coles, B.L.K.; van der Kooy, D. Glucocorticoid agonists enhance retinal stem cell self-renewal and proliferation. Stem Cell Res. Ther. 2021, 12, 83. [Google Scholar] [CrossRef]
- Muller, P.Y.; Janovjak, H.; Miserez, A.R.; Dobbie, Z. Processing of gene expression data generated by quantitative real-time RT-PCR. Biotechniques 2002, 32, 1372–1374, 1376, 1378–1379. [Google Scholar]
- Community, G. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2022 update. Nucleic Acids Res. 2022, 50, W345–W351. [Google Scholar] [CrossRef]
- Sloggett, C.; Goonasekera, N.; Afgan, E. BioBlend: Automating pipeline analyses within Galaxy and CloudMan. Bioinformatics 2013, 29, 1685–1686. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence | Accession Number |
|---|---|---|
| Ptafr | F: 5′-AGCAGAGTTGGGCTACCAGA-3′ R: 5′-TGCGCATGCTGTAAAACTTC-3′ | NM_001081211.2 |
| Lpcat2 | F: 5′-CCAGGTGGCATTTAAGCTCT-3′ R: 5′-TCTTGGCATATTCTGGGTGC-3′ | NM_173014.2 NM_001357375.1 NM_001357374.1 |
| Pafah | F: 5′-GTCTCTGCTTCAGAGGATGC-3′ R: 5′-ACATTGTGATCGTGACCGTG-3′ | NM_013625.4 |
| Palmd | F: 5′-ATTCTCTTCCTCTCTCCCTGCTGC-3′ R: 5′-GCTACCATAAATCAAGGTGCGTCC-3′ | NM_023245.3 |
| Ki67 | F: 5′-TAACGCCACCGAGGACAAAT-3′ R: 5′-TTCAGAAGCTCCACTTCGCC-3′ | NM_001081117.2 |
| NeuroD1 | F: 5′- ACGCAGAAGGCAAGGTGTCC -3′ R: 5′- TTGGTCATGTTTCCACTTCC -3′ | NM_010894.3 |
| Pax6 | F: 5′-TCTGGGCAGGTATTACGAGAC-3′ R: 5′-TTATCGTTGGTACAGACCCCCT-3′ | NM_001310146.1 NM_001310145.1 NM_001310144.1 NM_001244202.2 NM_001244201.2 NM_013627.6 NM_001244200.2 NM_001244198.2 |
| Nanog | F: 5′-CACAGTTTGCCTAGTTCTGAGG-3′ R: 5′-GCAAGAATAGTTCTCGGGATGAA-3′ | NM_028016.3 NM_001289828.1 NM_001289831.1 NM_001289830.1 |
| Sox2 | F: 5′-GCGGAGTGGAAACTTTTGTCC-3′ R: 5′-GGGAAGCGTGTACTTATCCTTCT-3′ | NM_011443.4 |
| CaM | F: 5′-GTCATGCGGTCACTGGGTCAG-3′ R: 5′-CAAGAACTCTGGGAAGTCAATGG-3′ | NM_009790.5 NM_001313934.1 |
| β-actin | F: 5′-TGAGCTGCGTTTTACACCCT-3′ R: 5′-GCCTTCACCGTTCCAGTTTT-3′ | NM_007393.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dalmaso, B.; Silva-Junior, I.A.d.; Jancar, S.; Del Debbio, C.B. Platelet-Activating Factor Receptor (PAFR) Regulates Retinal Progenitor/Stem Cells Profile in Ciliary Epithelium Cells. Int. J. Mol. Sci. 2024, 25, 3084. https://doi.org/10.3390/ijms25063084
Dalmaso B, Silva-Junior IAd, Jancar S, Del Debbio CB. Platelet-Activating Factor Receptor (PAFR) Regulates Retinal Progenitor/Stem Cells Profile in Ciliary Epithelium Cells. International Journal of Molecular Sciences. 2024; 25(6):3084. https://doi.org/10.3390/ijms25063084
Chicago/Turabian StyleDalmaso, Barbara, Ildefonso Alves da Silva-Junior, Sonia Jancar, and Carolina Beltrame Del Debbio. 2024. "Platelet-Activating Factor Receptor (PAFR) Regulates Retinal Progenitor/Stem Cells Profile in Ciliary Epithelium Cells" International Journal of Molecular Sciences 25, no. 6: 3084. https://doi.org/10.3390/ijms25063084
APA StyleDalmaso, B., Silva-Junior, I. A. d., Jancar, S., & Del Debbio, C. B. (2024). Platelet-Activating Factor Receptor (PAFR) Regulates Retinal Progenitor/Stem Cells Profile in Ciliary Epithelium Cells. International Journal of Molecular Sciences, 25(6), 3084. https://doi.org/10.3390/ijms25063084