
Local Production of Acute Phase Proteins: A Defense Reaction of Cancer Cells to Injury with Focus on Fibrinogen


Abstract
:1. Introduction of the Topic
2. Background: Description of the Model Used in the Reviewed Experiments
2.1. Breast Cancer (BC)
2.2. Modulated Electro-Hyperthermia (mEHT)
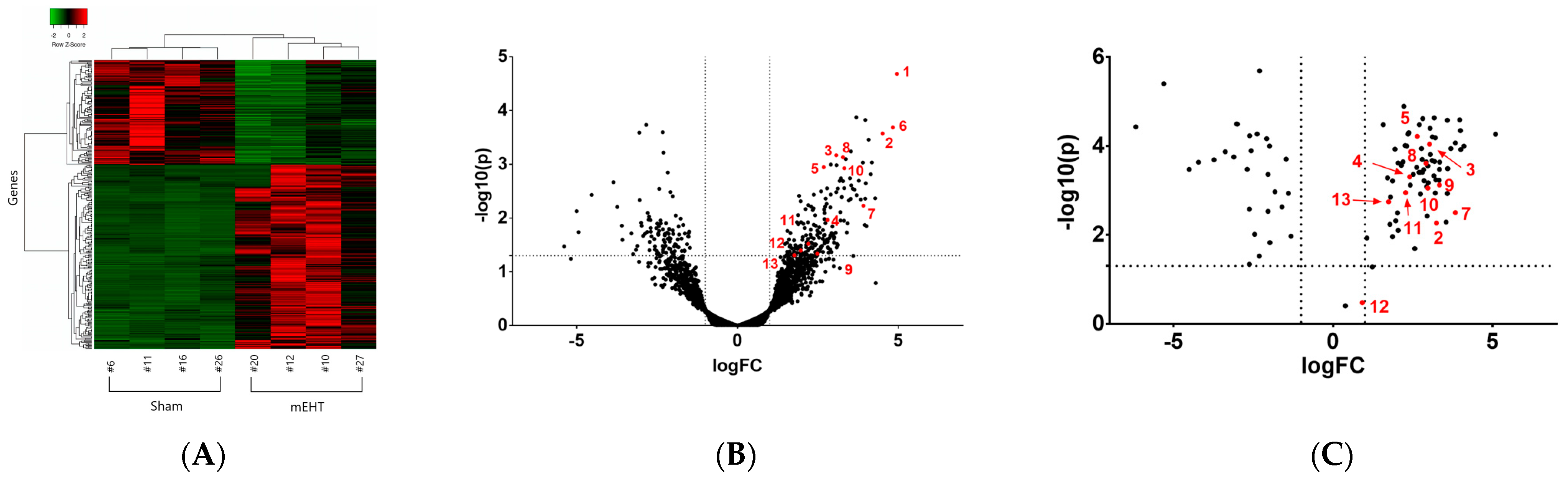
3. Experimental Evidence Demonstrating Local Acute Phase Protein and Fibrinogen (FN) Production in Cancer Treated with Modulated Electro-Hyperthermia
The Local Acute Phase Response (lAPR)
4. Fibrin(ogen) (FN)
4.1. The Structure of FN
4.2. The Physiological (Canonical and Non-Canonical) Functions of Fibrin(ogen) (FN) in the Blood and in the Extracellular Matrix (ECM)
4.3. Documented Roles of Fibrinogens in Non-Hematologic Disease
4.3.1. Fibrin(ogen) Plays a Role in Inflammatory Conditions
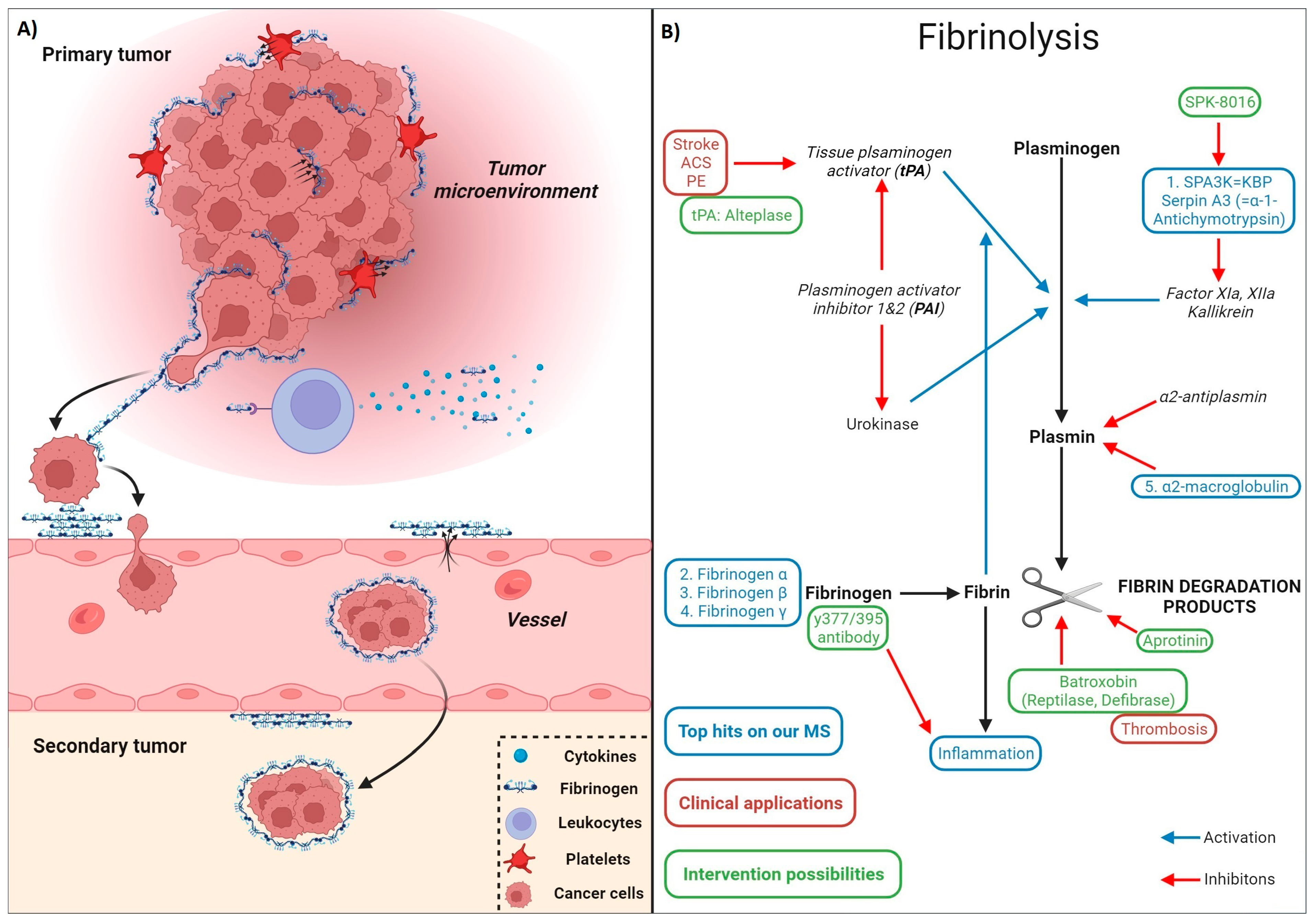
4.3.2. Documented Roles of Fibrin(ogen) in Cancer Progression

{kind=link}
{kind=link}
| FN Function/Molecular Mechanism | FN in Cancer Progression | Experimental Evidence | Reference |
|---|---|---|---|
| FN filtered from the circulation (FN deposition in the vicinity of blood vessels in tumors) | Compact stroma hinders therapy delivery | Removal of the compact stroma enhances therapy delivery | [44,45,49] |
| Enhancement of cancer cell extravasation | Lactadherin—by competing with FN for PS binding sites may delay tumor progression | [26,50] | |
| FN stimulates ECM formation | FN scaffold in the TME → adhesion, migration, invasion and metastasis of cancer cells | FN is a predominant component of the BC stroma. FN KO mice (Fgα−/−) were protected against hematogenous metastasis | [26,27,51,52] |
| FN binds to + surrounds cancer cells → protective structure from treatments and immune system | FN interaction with platelets via β3-integrins facilitates the protection of tumor cells from NK-cell cytotoxicity, permitting escape from host immune surveillance | [26,27,30] | |
| FN is important for the assembly of ECM to enhance host defense | FN contributes to the tumor favorable TME | In fibrinogen KO mice, the carcinogenesis gene-network was downregulated | [27,53] |
| FN supports cancer cell growth, survival and proliferation | Primary colon cancer development diminished in Fgα−/− mice | [54,55] | |
| FN net: growth factor reservoir | FN contributes to the proinflammatory TME to favor tumor progression | Interaction with αMβ2 induce leukocyte adhesion to ECs + proinflammatory cytokine production | [47,55] |
4.3.3. Fibrinolysis

4.3.4. Fibrin Induces a Tumor-Favorable Microenvironment (TME)
4.3.5. Fibrin Deposition-Induced Chronic Inflammation of the Tumor Microenvironment (TME)
4.3.6. Drugability of Fibrinogen
5. Summary and Future Perspectives
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| A1M | alpha-1-microglobulin |
| a2-MG | alpha-2-macroglobulin |
| AMBP | bikunin precursor |
| ACS | acute coronary syndrome |
| AKI | acute kidney injury |
| APP | acute-phase proteins |
| APR | acute-phase response |
| AT-III | antithrombin-3 |
| CKD | chronic kidney disease |
| CPB | carboxypeptidase-B |
| CRC | colorectal cancer |
| DVT | deep vein thrombosis |
| ECM | extracellular matrix |
| EMF | focused electromagnetic field |
| FN | fibrin(ogen) |
| GS | glomerular sclerosis |
| HIF-1 | hypoxia-inducible factor-1 |
| Hp | haptoglobin |
| HSR | heat shock response |
| IalphaI/ITI | inter-alpha-trypsin inhibitor |
| IL | interleukin |
| LMWH | low-molecular-weight heparin |
| LPS | endotoxin/lipopolysaccharide |
| mEHT | modulated electro-hyperthermia |
| MS | mass spectrometry |
| NGS | next-generation sequencing |
| NSCLC | non-small-cell lung cancer |
| PAI | plasminogen activator inhibitor |
| PCR | polymerase chain reaction |
| PE | pulmonary embolism |
| P | properidin |
| PTX3 | pentraxin-related gene |
| RT-PCR | real-time polymerase chain reaction |
| SERPIN | serine protease inhibitor |
| SPA3K | serine protease inhibitor (Serpin)-A3 = alpha-1-anitchymotrypsin |
| SPK | Spa3K inhibitor |
| TAFI | thrombin-activatable fibrinolysis inhibitor |
| TEC | tubular epithelial cell |
| TGF-beta | transforming growth factor beta |
| TME | tumor microenvironment |
| TNBC | triple-negative breast cancer |
| TNF-alpha | tumor necrosis factor-alpha |
| tPA | tissue plasminogen activator |
| TSL | thermo-sensitive liposome |
References
- Gabay, C.; Kushner, I. Acute-phase proteins and other systemic responses to inflammation. N. Engl. J. Med. 1999, 340, 448–454. [Google Scholar] [CrossRef]
- Moshage, H. Cytokines and the hepatic acute phase response. J. Pathol. 1997, 181, 257–266. [Google Scholar] [CrossRef]
- Schrodl, W.; Buchler, R.; Wendler, S.; Reinhold, P.; Muckova, P.; Reindl, J.; Rhode, H. Acute phase proteins as promising biomarkers: Perspectives and limitations for human and veterinary medicine. Proteom. Clin. Appl. 2016, 10, 1077–1092. [Google Scholar] [CrossRef] [PubMed]
- Schvarcz, C.A.; Danics, L.; Krenacs, T.; Viana, P.; Beres, R.; Vancsik, T.; Nagy, A.; Gyenesei, A.; Kun, J.; Fonovic, M.; et al. Modulated Electro-Hyperthermia Induces a Prominent Local Stress Response and Growth Inhibition in Mouse Breast Cancer Isografts. Cancers 2021, 13, 1744. [Google Scholar] [CrossRef] [PubMed]
- Roka, B.; Tod, P.; Kaucsar, T.; Vizovisek, M.; Vidmar, R.; Turk, B.; Fonovic, M.; Szenasi, G.; Hamar, P. The Acute Phase Response Is a Prominent Renal Proteome Change in Sepsis in Mice. Int. J. Mol. Sci. 2019, 21, 200. [Google Scholar] [CrossRef] [PubMed]
- Bukosza, E.N.; Kornauth, C.; Hummel, K.; Schachner, H.; Huttary, N.; Krieger, S.; Nobauer, K.; Oszwald, A.; Razzazi Fazeli, E.; Kratochwill, K.; et al. ECM Characterization Reveals a Massive Activation of Acute Phase Response during FSGS. Int. J. Mol. Sci. 2020, 21, 2095. [Google Scholar] [CrossRef] [PubMed]
- Kaucsar, T.; Bodor, C.; Godo, M.; Szalay, C.; Revesz, C.; Nemeth, Z.; Mozes, M.; Szenasi, G.; Rosivall, L.; Soti, C.; et al. LPS-induced delayed preconditioning is mediated by Hsp90 and involves the heat shock response in mouse kidney. PLoS ONE 2014, 9, e92004. [Google Scholar] [CrossRef]
- Danics, L.; Schvarcz, C.A.; Viana, P.; Vancsik, T.; Krenacs, T.; Benyo, Z.; Kaucsar, T.; Hamar, P. Exhaustion of Protective Heat Shock Response Induces Significant Tumor Damage by Apoptosis after Modulated Electro-Hyperthermia Treatment of Triple Negative Breast Cancer Isografts in Mice. Cancers 2020, 12, 2581. [Google Scholar] [CrossRef]
- Mantovani, A.; Garlanda, C. Humoral Innate Immunity and Acute-Phase Proteins. N. Engl. J. Med. 2023, 388, 439–452. [Google Scholar] [CrossRef]
- Janciauskiene, S.; Wrenger, S.; Gunzel, S.; Grunding, A.R.; Golpon, H.; Welte, T. Potential Roles of Acute Phase Proteins in Cancer: Why Do Cancer Cells Produce or Take Up Exogenous Acute Phase Protein Alpha1-Antitrypsin? Front. Oncol. 2021, 11, 622076. [Google Scholar] [CrossRef]
- Serrano, I.; Luque, A.; Aran, J.M. Exploring the Immunomodulatory Moonlighting Activities of Acute Phase Proteins for Tolerogenic Dendritic Cell Generation. Front. Immunol. 2018, 9, 892. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Huang, Y.; Zhou, M.; Guo, Y.; Zhou, Y.; Wang, R.; Jin, W.; Wen, C.; Zhang, Y.; Tang, Y. Artesunate attenuates serum amyloid A-induced M1 macrophage differentiation through the promotion of PHGDH. Int. Immunopharmacol. 2024, 127, 111462. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Rao, G.; Heimberger, A.; Li, S. Fibrinogen-like protein 2: Its biological function across cell types and the potential to serve as an immunotherapy target for brain tumors. Cytokine Growth Factor Rev. 2023, 69, 73–79. [Google Scholar] [CrossRef]
- Shephard, E.G.; Anderson, R.; Strachan, A.F.; Kuhn, S.H.; De Beer, F.C. CRP and neutrophils: Functional effects and complex uptake. Clin. Exp. Immunol. 1986, 63, 718–727. [Google Scholar]
- Jimenez, R.V.; Wright, T.T.; Jones, N.R.; Wu, J.; Gibson, A.W.; Szalai, A.J. C-Reactive Protein Impairs Dendritic Cell Development, Maturation, and Function: Implications for Peripheral Tolerance. Front. Immunol. 2018, 9, 372. [Google Scholar] [CrossRef] [PubMed]
- Tremble, P.; Chiquet-Ehrismann, R.; Werb, Z. The extracellular matrix ligands fibronectin and tenascin collaborate in regulating collagenase gene expression in fibroblasts. Mol. Biol. Cell 1994, 5, 439–453. [Google Scholar] [CrossRef]
- Ban, H.S.; Han, T.S.; Hur, K.; Cho, H.S. Epigenetic Alterations of Heat Shock Proteins (HSPs) in Cancer. Int. J. Mol. Sci. 2019, 20, 4758. [Google Scholar] [CrossRef]
- Asea, A.; Ara, G.; Teicher, B.A.; Stevenson, M.A.; Calderwood, S.K. Effects of the flavonoid drug quercetin on the response of human prostate tumours to hyperthermia in vitro and in vivo. Int. J. Hyperth. 2001, 17, 347–356. [Google Scholar] [CrossRef]
- Le, M.T.; Hamar, P.; Guo, C.; Basar, E.; Perdigao-Henriques, R.; Balaj, L.; Lieberman, J. miR-200-containing extracellular vesicles promote breast cancer cell metastasis. J. Clin. Investig. 2014, 124, 5109–5128. [Google Scholar] [CrossRef]
- Ackermann, M.R. Chapter 3—Inflammation and Healing1. In Pathologic Basis of Veterinary Disease, 6th ed.; Zachary, J.F., Ed.; Mosby: Amsterdam, The Netherlands, 2017; pp. 73–131.e2. [Google Scholar]
- Ehlting, C.; Wolf, S.D.; Bode, J.G. Acute-phase protein synthesis: A key feature of innate immune functions of the liver. Biol. Chem. 2021, 402, 1129–1145. [Google Scholar] [CrossRef]
- Sabina, J.; Tobias, W.; Ravi, M. Acute Phase Proteins: Structure and Function Relationship. In Acute Phase Proteins; Francisco, V., Ed.; IntechOpen: Rijeka, Croatia, 2011; p. 2. [Google Scholar]
- Pepys, M.B. Acute Phase Proteins. In Encyclopedia of Immunology, 2nd ed.; Delves, P.J., Ed.; Elsevier: Oxford, UK, 1998; pp. 18–20. [Google Scholar]
- Mosesson, M.W. Fibrinogen and fibrin structure and functions. J. Thromb. Haemost. 2005, 3, 1894–1904. [Google Scholar] [CrossRef]
- Davalos, D.; Ryu, J.K.; Merlini, M.; Baeten, K.M.; Le Moan, N.; Petersen, M.A.; Deerinck, T.J.; Smirnoff, D.S.; Bedard, C.; Hakozaki, H.; et al. Fibrinogen-induced perivascular microglial clustering is required for the development of axonal damage in neuroinflammation. Nat. Commun. 2012, 3, 1227. [Google Scholar] [CrossRef]
- Vilar, R.; Fish, R.J.; Casini, A.; Neerman-Arbez, M. Fibrin(ogen) in human disease: Both friend and foe. Haematologica 2020, 105, 284–296. [Google Scholar] [CrossRef]
- Simpson-Haidaris, P.J.; Rybarczyk, B. Tumors and fibrinogen. The role of fibrinogen as an extracellular matrix protein. Ann. N. Y. Acad. Sci. 2001, 936, 406–425. [Google Scholar] [CrossRef]
- Bottazzi, B.; Doni, A.; Garlanda, C.; Mantovani, A. An integrated view of humoral innate immunity: Pentraxins as a paradigm. Annu. Rev. Immunol. 2010, 28, 157–183. [Google Scholar] [CrossRef]
- Zuliani-Alvarez, L.; Midwood, K.S. Fibrinogen-Related Proteins in Tissue Repair: How a Unique Domain with a Common Structure Controls Diverse Aspects of Wound Healing. Adv. Wound Care 2015, 4, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Mendiola, A.S.; Yan, Z.; Dixit, K.; Johnson, J.R.; Bouhaddou, M.; Meyer-Franke, A.; Shin, M.G.; Yong, Y.; Agrawal, A.; MacDonald, E.; et al. Defining blood-induced microglia functions in neurodegeneration through multiomic profiling. Nat. Immunol. 2023, 24, 1173–1187. [Google Scholar] [CrossRef] [PubMed]
- Degen, J.L.; Drew, A.F.; Palumbo, J.S.; Kombrinck, K.W.; Bezerra, J.A.; Danton, M.J.; Holmbäck, K.; Suh, T.T. Genetic manipulation of fibrinogen and fibrinolysis in mice. Ann. N. Y. Acad. Sci. 2001, 936, 276–290. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Topete, D.; Iwaki, T.; Ploplis, V.; Castellino, F. Delayed inflammatory responses to endotoxin in fibrinogen-deficient mice. J. Pathol. 2006, 210, 325–333. [Google Scholar]
- Sörensen, I.; Susnik, N.; Inhester, T.; Degen, J.L.; Melk, A.; Haller, H.; Schmitt, R. Fibrinogen, acting as a mitogen for tubulointerstitial fibroblasts, promotes renal fibrosis. Kidney Int. 2011, 80, 1035–1044. [Google Scholar] [CrossRef] [PubMed]
- Petzelbauer, P.; Zacharowski, P.A.; Miyazaki, Y.; Friedl, P.; Wickenhauser, G.; Castellino, F.J.; Gröger, M.; Wolff, K.; Zacharowski, K. The fibrin-derived peptide Bbeta15-42 protects the myocardium against ischemia-reperfusion injury. Nat. Med. 2005, 11, 298–304. [Google Scholar] [CrossRef]
- Kannel, W.B.; Wolf, P.A.; Castelli, W.P.; D’Agostino, R.B. Fibrinogen and Risk of Cardiovascular Disease: The Framingham Study. JAMA 1987, 258, 1183–1186. [Google Scholar] [CrossRef]
- Rooney, T.; Scherzer, R.; Shigenaga, J.K.; Graf, J.; Imboden, J.B.; Grunfeld, C. Levels of plasma fibrinogen are elevated in well-controlled rheumatoid arthritis. Rheumatology 2011, 50, 1458–1465. [Google Scholar] [CrossRef]
- Chen, X.F.; Zhao, Y.; Guo, Y.; Huang, Z.M.; Huang, X.L. Predictive value of fibrinogen in identifying inflammatory bowel disease in active stage. BMC Gastroenterol. 2021, 21, 472. [Google Scholar] [CrossRef]
- Martín-Timón, I.; Sevillano-Collantes, C.; Segura-Galindo, A.; Del Cañizo-Gómez, F.J. Type 2 diabetes and cardiovascular disease: Have all risk factors the same strength? World J. Diabetes 2014, 5, 444–470. [Google Scholar] [CrossRef]
- Solá, E.; Vayá, A.; Simó, M.; Hernández-Mijares, A.; Morillas, C.; España, F.; Estellés, A.; Corella, D. Fibrinogen, plasma viscosity and blood viscosity in obesity. Relationship with insulin resistance. Clin. Hemorheol. Microcirc. 2007, 37, 309–318. [Google Scholar]
- Petersen, M.A.; Ryu, J.K.; Akassoglou, K. Fibrinogen in neurological diseases: Mechanisms, imaging and therapeutics. Nat. Rev. Neurosci. 2018, 19, 283–301. [Google Scholar] [CrossRef] [PubMed]
- Akassoglou, K. The immunology of blood: Connecting the dots at the neurovascular interface. Nat. Immunol. 2020, 21, 710–712. [Google Scholar] [CrossRef] [PubMed]
- Merlini, M.; Rafalski, V.A.; Rios Coronado, P.E.; Gill, T.M.; Ellisman, M.; Muthukumar, G.; Subramanian, K.S.; Ryu, J.K.; Syme, C.A.; Davalos, D.; et al. Fibrinogen Induces Microglia-Mediated Spine Elimination and Cognitive Impairment in an Alzheimer’s Disease Model. Neuron 2019, 101, 1099–1108.e1096. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Cao, Z.; Ma, Y.; Wang, J.; Zhang, L.; Luo, Z. Fibrinogen, a Promising Marker to Evaluate Severity and Prognosis of Acute Exacerbation of Chronic Obstructive Pulmonary Disease: A Retrospective Observational Study. Int. J. Chron. Obs. Pulmon Dis. 2022, 17, 1299–1310. [Google Scholar] [CrossRef] [PubMed]
- Mei, T.; Shashni, B.; Maeda, H.; Nagasaki, Y. Fibrinolytic tissue plasminogen activator installed redox-active nanoparticles (t-PA@iRNP) for cancer therapy. Biomaterials 2020, 259, 120290. [Google Scholar] [CrossRef]
- Zhang, B.; Jiang, T.; She, X.; Shen, S.; Wang, S.; Deng, J.; Shi, W.; Mei, H.; Hu, Y.; Pang, Z.; et al. Fibrin degradation by rtPA enhances the delivery of nanotherapeutics to A549 tumors in nude mice. Biomaterials 2016, 96, 63–71. [Google Scholar] [CrossRef]
- Sahni, A.; Simpson-Haidaris, P.J.; Sahni, S.K.; Vaday, G.G.; Francis, C.W. Fibrinogen synthesized by cancer cells augments the proliferative effect of fibroblast growth factor-2 (FGF-2). J. Thromb. Haemost. 2008, 6, 176–183. [Google Scholar] [CrossRef]
- Rybarczyk, B.J.; Simpson-Haidaris, P.J. Fibrinogen assembly, secretion, and deposition into extracellular matrix by MCF-7 human breast carcinoma cells. Cancer Res. 2000, 60, 2033–2039. [Google Scholar]
- Hamar, P. A New Role of Acute Phase Proteins: Local Production Is an Ancient, General Stress-Response System of Mammalian Cells. Int. J. Mol. Sci. 2022, 23, 2972. [Google Scholar] [CrossRef] [PubMed]
- Nagy, J.A.; Brown, L.F.; Senger, D.R.; Lanir, N.; Van de Water, L.; Dvorak, A.M.; Dvorak, H.F. Pathogenesis of tumor stroma generation: A critical role for leaky blood vessels and fibrin deposition. Biochim. Biophys. Acta 1989, 948, 305–326. [Google Scholar] [CrossRef] [PubMed]
- Jing, H.; Wu, X.; Xiang, M.; Wang, C.; Novakovic, V.A.; Shi, J. Microparticle Phosphatidylserine Mediates Coagulation: Involvement in Tumor Progression and Metastasis. Cancers 2023, 15, 1957. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, H.F.; Senger, D.R.; Dvorak, A.M. Fibrin as a component of the tumor stroma: Origins and biological significance. Cancer Metastasis Rev. 1983, 2, 41–73. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Li, Y.; Zhang, S.; Wang, X.; Dou, H.; Yu, X.; Zhang, Z.; Yang, S.; Xiao, M. Extracellular matrix remodeling in tumor progression and immune escape: From mechanisms to treatments. Mol. Cancer 2023, 22, 48. [Google Scholar] [CrossRef] [PubMed]
- Hadjipanayi, E.; Kuhn, P.H.; Moog, P.; Bauer, A.T.; Kuekrek, H.; Mirzoyan, L.; Hummel, A.; Kirchhoff, K.; Salgin, B.; Isenburg, S.; et al. The Fibrin Matrix Regulates Angiogenic Responses within the Hemostatic Microenvironment through Biochemical Control. PLoS ONE 2015, 10, e0135618. [Google Scholar] [CrossRef] [PubMed]
- Adams, G.N.; Rosenfeldt, L.; Frederick, M.; Miller, W.; Waltz, D.; Kombrinck, K.; McElhinney, K.E.; Flick, M.J.; Monia, B.P.; Revenko, A.S.; et al. Colon Cancer Growth and Dissemination Relies upon Thrombin, Stromal PAR-1, and Fibrinogen. Cancer Res. 2015, 75, 4235–4243. [Google Scholar] [CrossRef]
- Gupta, M.K.; Qin, R.Y. Mechanism and its regulation of tumor-induced angiogenesis. World J. Gastroenterol. 2003, 9, 1144–1155. [Google Scholar] [CrossRef]
- Lin, H.; Xu, L.; Yu, S.; Hong, W.; Huang, M.; Xu, P. Therapeutics targeting the fibrinolytic system. Exp. Mol. Med. 2020, 52, 367–379. [Google Scholar] [CrossRef]
- Kryvalap, Y.; Czyzyk, J. The Role of Proteases and Serpin Protease Inhibitors in beta-Cell Biology and Diabetes. Biomolecules 2022, 12, 67. [Google Scholar] [CrossRef]
- Baskin, J.L.; Pui, C.H.; Reiss, U.; Wilimas, J.A.; Metzger, M.L.; Ribeiro, R.C.; Howard, S.C. Management of occlusion and thrombosis associated with long-term indwelling central venous catheters. Lancet 2009, 374, 159–169. [Google Scholar] [CrossRef]
- Gomez-Outes, A.; Terleira-Fernandez, A.I.; Calvo-Rojas, G.; Suarez-Gea, M.L.; Vargas-Castrillon, E. Dabigatran, Rivaroxaban, or Apixaban versus Warfarin in Patients with Nonvalvular Atrial Fibrillation: A Systematic Review and Meta-Analysis of Subgroups. Thrombosis 2013, 2013, 640723. [Google Scholar] [CrossRef]
- Therapeutics, S. Hemophilia a Inhibitor. Available online: https://sparktx.com/pipelines/spk-8016-hemophilia-a-inhibitor-market/ (accessed on 6 November 2018).
- Reddy, R.; Reddy, N. Biomimetic approaches for tissue engineering. J. Biomater. Sci. Polym. Ed. 2018, 29, 1667–1685. [Google Scholar] [CrossRef] [PubMed]
- Peltanova, B.; Raudenska, M.; Masarik, M. Effect of tumor microenvironment on pathogenesis of the head and neck squamous cell carcinoma: A systematic review. Mol. Cancer 2019, 18, 63. [Google Scholar] [CrossRef] [PubMed]
- Popova, N.V.; Jucker, M. The Functional Role of Extracellular Matrix Proteins in Cancer. Cancers 2022, 14, 238. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Cai, J.; Xie, J.; Dai, Y. Extracellular fibrin promotes non-small cell lung cancer progression through integrin beta1/PTEN/AKT signaling. Open Life Sci. 2023, 18, 20220716. [Google Scholar] [CrossRef] [PubMed]
- Laurens, N.; Koolwijk, P.; de Maat, M.P. Fibrin structure and wound healing. J. Thromb. Haemost. 2006, 4, 932–939. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhang, L.; Wan, D.; Zhou, L.; Zheng, S.; Lin, S.; Qiao, Y. Extracellular matrix and its therapeutic potential for cancer treatment. Signal Transduct. Target. Ther. 2021, 6, 153. [Google Scholar] [CrossRef]
- Greten, F.R.; Grivennikov, S.I. Inflammation and Cancer: Triggers, Mechanisms, and Consequences. Immunity 2019, 51, 27–41. [Google Scholar] [CrossRef]
- Chen, C.; Wang, Z.; Ding, Y.; Qin, Y. Tumor microenvironment-mediated immune evasion in hepatocellular carcinoma. Front. Immunol. 2023, 14, 1133308. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.K.; Rafalski, V.A.; Meyer-Franke, A.; Adams, R.A.; Poda, S.B.; Rios Coronado, P.E.; Pedersen, L.; Menon, V.; Baeten, K.M.; Sikorski, S.L.; et al. Fibrin-targeting immunotherapy protects against neuroinflammation and neurodegeneration. Nat. Immunol. 2018, 19, 1212–1223. [Google Scholar] [CrossRef] [PubMed]
- Aloss, K.; Hamar, P. Recent Preclinical and Clinical Progress in Liposomal Doxorubicin. Pharmaceutics 2023, 15, 893. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamar, P. Local Production of Acute Phase Proteins: A Defense Reaction of Cancer Cells to Injury with Focus on Fibrinogen. Int. J. Mol. Sci. 2024, 25, 3435. https://doi.org/10.3390/ijms25063435
Hamar P. Local Production of Acute Phase Proteins: A Defense Reaction of Cancer Cells to Injury with Focus on Fibrinogen. International Journal of Molecular Sciences. 2024; 25(6):3435. https://doi.org/10.3390/ijms25063435
Chicago/Turabian StyleHamar, Péter. 2024. "Local Production of Acute Phase Proteins: A Defense Reaction of Cancer Cells to Injury with Focus on Fibrinogen" International Journal of Molecular Sciences 25, no. 6: 3435. https://doi.org/10.3390/ijms25063435
APA StyleHamar, P. (2024). Local Production of Acute Phase Proteins: A Defense Reaction of Cancer Cells to Injury with Focus on Fibrinogen. International Journal of Molecular Sciences, 25(6), 3435. https://doi.org/10.3390/ijms25063435