
Standard Radio-Iodine Labeling Protocols Impaired the Functional Integrity of Mesenchymal Stem/Stromal Cell Exosomes

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
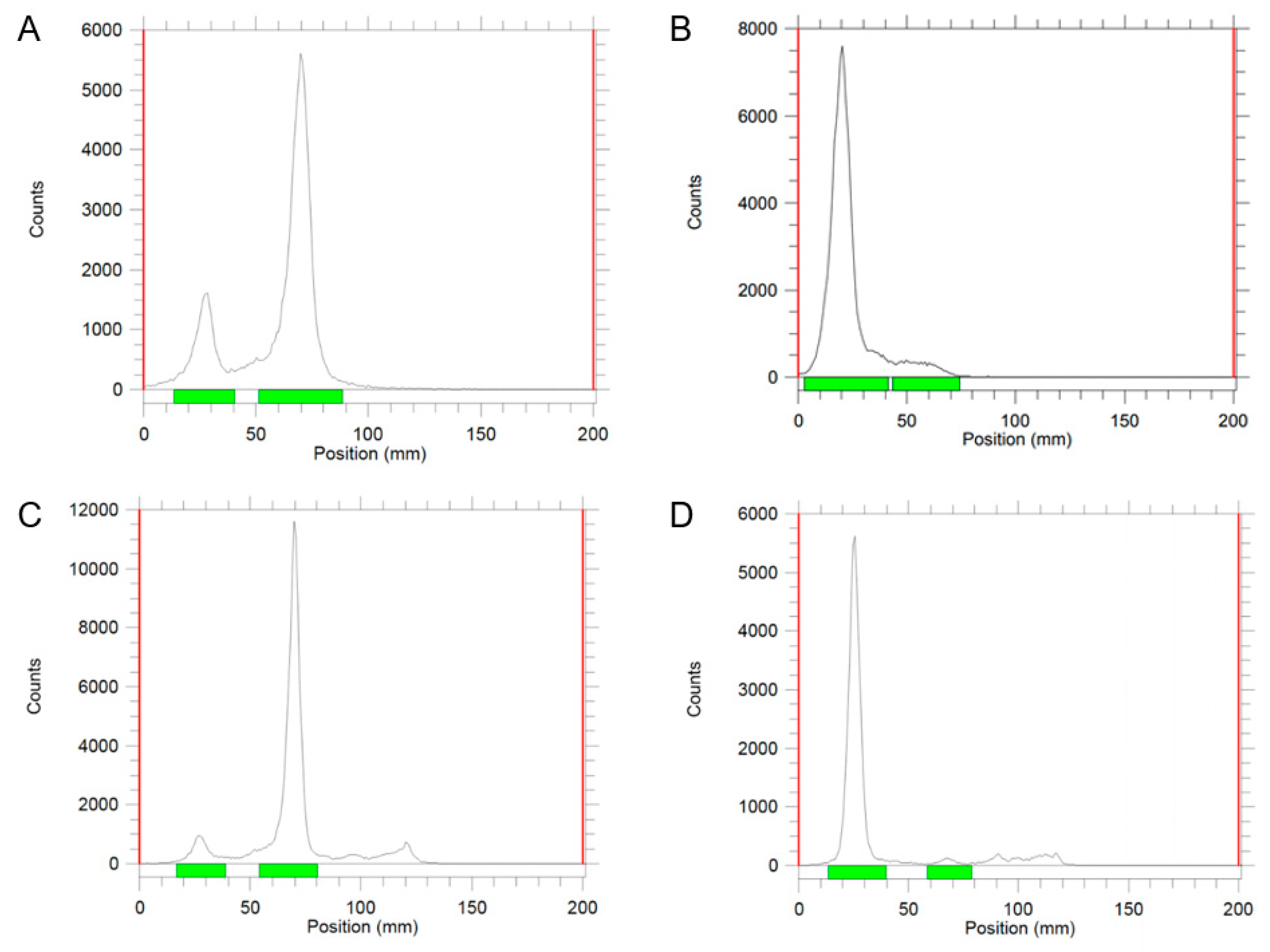
2.1. Radiolabeling
2.2. Integrity of Exosomes after Iodine Labeling
3. Discussion
4. Materials and Methods
4.1. Preparation and Culture Conditions of MSC Exosomes
4.2. Radiolabeling
4.3. Nanoparticle Tracking Analysis
4.4. CD73 Enzymatic Activity Measurements
4.5. Statistical Tests
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal Criteria for Defining Multipotent Mesenchymal Stromal Cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. Available online: http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&list_uids=16923606 (accessed on 20 March 2024). [CrossRef]
- Hass, R.; Kasper, C.; Böhm, S.; Jacobs, R. Different populations and sources of human mesenchymal stem cells (MSC): A comparison of adult and neonatal tissue-derived MSC. Cell Commun. Signal. 2011, 9, 12. [Google Scholar] [CrossRef]
- Chopp, M.; Li, Y. Treatment of neural injury with marrow stromal cells. Lancet Neurol. 2002, 1, 92–100. [Google Scholar] [CrossRef]
- Miclau, K.; Hambright, W.S.; Huard, J.; Stoddart, M.J.; Bahney, C.S. Cellular Expansion of MSCs: Shifting the Regenerative Potential. Aging Cell 2023, 22, e13759. Available online: https://www.ncbi.nlm.nih.gov/pubmed/36536521 (accessed on 20 March 2024). [CrossRef]
- Han, Y.; Yang, J.; Fang, J.; Zhou, Y.; Candi, E.; Wang, J.; Hua, D.; Shao, C.; Shi, Y. The Secretion Profile of Mesenchymal Stem Cells and Potential Applications in Treating Human Diseases. Signal Transduct. Target. Ther. 2022, 7, 92. Available online: https://www.ncbi.nlm.nih.gov/pubmed/35314676 (accessed on 20 March 2024). [CrossRef]
- Timmers, L.; Lim, S.-K.; Arslan, F.; Armstrong, J.S.; Hoefler, I.E.; Doevendans, P.A.; Piek, J.J.; El Oakley, R.M.; Choo, A.; Lee, C.N.; et al. Reduction of myocardial infarct size by human mesenchymal stem cell conditioned medium. Stem Cell Res. 2008, 1, 129–137. [Google Scholar] [CrossRef]
- Bruno, S.; Grange, C.; Deregibus, M.C.; Calogero, R.A.; Saviozzi, S.; Collino, F.; Morando, L.; Busca, A.; Falda, M.; Bussolati, B.; et al. Mesenchymal stem cell-derived microvesicles protect against acute tubular injury. J. Am. Soc. Nephrol. 2009, 20, 1053–1067. [Google Scholar] [CrossRef]
- Lai, R.C.; Arslan, F.; Lee, M.M.; Sze, N.S.; Choo, A.; Chen, T.S.; Salto-Tellez, M.; Timmers, L.; Lee, C.N.; El Oakley, R.M.; et al. Exosome secreted by MSC reduces myocardial ischemia/reperfusion injury. Stem Cell Res. 2010, 4, 214–222. [Google Scholar] [CrossRef]
- Bruno, S.; Tapparo, M.; Collino, F.; Chiabotto, G.; Deregibus, M.C.; Soares Lindoso, R.; Neri, F.; Kholia, S.; Giunti, S.; Wen, S. Renal regenerative potential of different extracellular vesicle populations derived from bone marrow mesenchymal stromal cells. Tissue Eng. Part A 2017, 23, 1262–1273. [Google Scholar] [CrossRef]
- Doeppner, T.R.; Herz, J.; Görgens, A.; Schlechter, J.; Ludwig, A.-K.; Radtke, S.; de Miroschedji, K.; Horn, P.A.; Giebel, B.; Hermann, D.M. Extracellular Vesicles Improve Post-Stroke Neuroregeneration and Prevent Postischemic Immunosuppression. Stem Cells Transl. Med. 2015, 4, 1131–1143. [Google Scholar] [CrossRef]
- He, J.; Wang, Y.; Sun, S.; Yu, M.; Wang, C.; Pei, X.; Zhu, B.; Wu, J.; Zhao, W. Bone Marrow Stem Cells-Derived Microvesicles Protect against Renal Injury in the Mouse Remnant Kidney Model. Nephrology 2012, 17, 493–500. Available online: http://www.ncbi.nlm.nih.gov/pubmed/22369283 (accessed on 20 March 2024). [CrossRef]
- Tan, F.; Li, X.; Wang, Z.; Li, J.; Shahzad, K.; Zheng, J. Clinical Applications of Stem Cell-Derived Exosomes. Signal Transduct. Target. Ther. 2024, 9, 17. Available online: https://www.ncbi.nlm.nih.gov/pubmed/38212307 (accessed on 20 March 2024). [CrossRef]
- Witwer, K.W.; Van Balkom, B.W.M.; Bruno, S.; Choo, A.; Dominici, M.; Gimona, M.; Hill, A.F.; De Kleijn, D.; Koh, M.; Lai, R.C.; et al. Defining Mesenchymal Stromal Cell (MSC)-Derived Small Extracellular Vesicles for Therapeutic Applications. J. Extracell. Vesicles 2019, 8, 1609206. Available online: https://www.ncbi.nlm.nih.gov/pubmed/31069028 (accessed on 20 March 2024). [CrossRef]
- Witwer, K.W.; Thery, C. Extracellular Vesicles or Exosomes? On Primacy, Precision, and Popularity Influencing a Choice of Nomenclature. J. Extracell. Vesicles 2019, 8, 1648167. Available online: https://www.ncbi.nlm.nih.gov/pubmed/31489144 (accessed on 20 March 2024). [CrossRef]
- Rezaie, J.; Feghhi, M.; Etemadi, T. A Review on Exosomes Application in Clinical Trials: Perspective, Questions, and Challenges. Cell Commun. Signal. 2022, 20, 145. Available online: https://www.ncbi.nlm.nih.gov/pubmed/36123730 (accessed on 20 March 2024). [CrossRef]
- Song, Y.; Kim, Y.; Ha, S.; Sheller-Miller, S.; Yoo, J.; Choi, C.; Park, C.H. The Emerging Role of Exosomes as Novel Therapeutics: Biology, Technologies, Clinical Applications, and the Next. Am. J. Reprod. Immunol. 2021, 85, e13329. Available online: https://www.ncbi.nlm.nih.gov/pubmed/32846024 (accessed on 20 March 2024). [CrossRef]
- Tai, Y.L.; Chen, K.C.; Hsieh, J.T.; Shen, T.L. Exosomes in Cancer Development and Clinical Applications. Cancer Sci. 2018, 109, 2364–2374. Available online: https://www.ncbi.nlm.nih.gov/pubmed/29908100 (accessed on 20 March 2024). [CrossRef]
- Tan, S.S.; Yin, Y.; Lee, T.; Lai, R.C.; Yeo, R.W.Y.; Zhang, B.; Choo, A.; Lim, S.K. Therapeutic MSC Exosomes Are Derived from Lipid Raft Microdomains in the Plasma Membrane. J. Extracell. Vesicles 2013, 2, 22614. Available online: https://pubmed.ncbi.nlm.nih.gov/24371518 (accessed on 20 March 2024). [CrossRef]
- Lai, R.C.; Chen, T.S.; Lim, S.K. Mesenchymal Stem Cell Exosome: A Novel Stem Cell-Based Therapy for Cardiovascular Disease. Regen. Med. 2011, 6, 481–492. Available online: https://www.ncbi.nlm.nih.gov/pubmed/21749206 (accessed on 20 March 2024). [CrossRef] [PubMed]
- Gimona, M.; Brizzi, M.F.; Choo, A.B.H.; Dominici, M.; Davidson, S.M.; Grillari, J.; Hermann, D.M.; Hill, A.F.; de Kleijn, D.; Lai, R.C.; et al. Critical Considerations for the Development of Potency Tests for Therapeutic Applications of Mesenchymal Stromal Cell-Derived Small Extracellular Vesicles. Cytotherapy 2021, 23, 373–380. Available online: https://www.sciencedirect.com/science/article/pii/S1465324921000013 (accessed on 20 March 2024). [CrossRef] [PubMed]
- Yi, Y.W.; Lee, J.H.; Kim, S.Y.; Pack, C.G.; Ha, D.H.; Park, S.R.; Youn, J.; Cho, B.S. Advances in Analysis of Biodistribution of Exosomes by Molecular Imaging. Int. J. Mol. Sci. 2020, 21, 665. Available online: https://www.ncbi.nlm.nih.gov/pubmed/31963931 (accessed on 20 March 2024). [CrossRef]
- Di Rocco, G.; Baldari, S.; Toietta, G. Towards Therapeutic Delivery of Extracellular Vesicles: Strategies for In Vivo Tracking and Biodistribution Analysis. Stem Cells Int. 2016, 2016, 5029619. Available online: https://www.ncbi.nlm.nih.gov/pubmed/27994623 (accessed on 20 March 2024). [CrossRef]
- Almeida, S.; Santos, L.; Falcão, A.; Gomes, C.; Abrunhosa, A. In Vivo Tracking of Extracellular Vesicles by Nuclear Imaging: Advances in Radiolabeling Strategies. Int. J. Mol. Sci. 2020, 21, 9443. Available online: https://www.mdpi.com/1422-0067/21/24/9443 (accessed on 20 March 2024). [CrossRef]
- Khan, A.A.; de Rosales, R.T.M. Radiolabelling of Extracellular Vesicles for PET and SPECT imaging. Nanotheranostics 2021, 5, 256–274. Available online: https://www.ncbi.nlm.nih.gov/pubmed/33654653 (accessed on 20 March 2024). [CrossRef]
- Hong, C.M.; Gangadaran, P.; Oh, J.M.; Rajendran, R.L.; Gopal, A.; Zhu, L.; Ahn, B.-C. Radioiodine labeling and in vivo trafficking of extracellular vesicles. Sci. Rep. 2021, 11, 5041. [Google Scholar] [CrossRef]
- Rashid, M.H.; Borin, T.F.; Ara, R.; Angara, K.; Cai, J.; Achyut, B.R.; Liu, Y.; Arbab, A.S. Differential in vivo biodistribution of (131)I-labeled exosomes from diverse cellular origins and its implication for theranostic application. Nanomedicine 2019, 21, 102072. [Google Scholar] [CrossRef]
- Guo, Q.; Zhao, C.; Gao, X.; Ding, L.; Wang, P.; Ren, Y.n.; Hou, X.; Yao, Y.; Zhang, C.; Yang, X.; et al. One-Minute Iodine Isotope Labeling Technology Enables Noninvasive Tracking and Quantification of Extracellular Vesicles in Tumor Lesions and Intact Animals. Mol. Pharm. 2023, 20, 3672–3682. [Google Scholar] [CrossRef]
- Rein, D.T.; Volkmer, A.K.; Volkmer, J.; Beyer, I.M.; Janni, W.; Fleisch, M.C.; Welter, A.K.; Bauerschlag, D.; Schöndorf, T.; Breidenbach, M. Systemic administration of bevacizumab prolongs survival in an in vivo model of platinum pre-treated ovarian cancer. Oncol. Lett. 2012, 3, 530–534. [Google Scholar] [CrossRef]
- Takov, K.; Yellon, D.M.; Davidson, S.M. Confounding factors in vesicle uptake studies using fluorescent lipophilic membrane dyes. J. Extracell. Vesicles 2017, 6, 1388731. [Google Scholar] [CrossRef]
- Tertel, T.; Schoppet, M.; Stambouli, O.; Al-Jipouri, A.; James, P.F.; Giebel, B. Imaging Flow Cytometry Challenges the Usefulness of Classically Used Extracellular Vesicle Labeling Dyes and Qualifies the Novel Dye Exoria for the Labeling of Mesenchymal Stromal Cell–Extracellular Vesicle Preparations. Cytotherapy 2022, 24, 619–628. Available online: https://www.sciencedirect.com/science/article/pii/S1465324922000494 (accessed on 20 March 2024). [CrossRef]
- Hansen, M.S.; Gadegaard, I.S.E.; Arnspang, E.C.; Blans, K.; Nejsum, L.N.; Rasmussen, J.T. Specific and Non-Invasive Fluorescent Labelling of Extracellular Vesicles for Evaluation of Intracellular Processing by Intestinal Epithelial Cells. Biomedicines 2020, 8, 211. Available online: https://www.ncbi.nlm.nih.gov/pubmed/32674302 (accessed on 20 March 2024). [CrossRef]
- Petrov, S.A.; Yusubov, M.S.; Beloglazkina, E.K.; Nenajdenko, V.G. Synthesis of Radioiodinated Compounds. Classical Approaches and Achievements of Recent Years. Int. J. Mol. Sci. 2022, 23, 3789. [Google Scholar] [CrossRef]
- Morishita, M.; Takahashi, Y.; Nishikawa, M.; Sano, K.; Kato, K.; Yamashita, T.; Imai, T.; Saji, H.; Takakura, Y. Quantitative analysis of tissue distribution of the B16BL6-derived exosomes using a streptavidin-lactadherin fusion protein and iodine-125-labeled biotin derivative after intravenous injection in mice. J. Pharm. Sci. 2015, 104, 705–713. [Google Scholar] [CrossRef]
- Chen, T.S.; Arslan, F.; Yin, Y.; Tan, S.S.; Lai, R.C.; Choo, A.B.; Padmanabhan, J.; Lee, C.N.; de Kleijn, D.P.; Lim, S.K. Enabling a Robust Scalable Manufacturing Process for Therapeutic Exosomes through Oncogenic Immortalization of Human ESC-Derived MSCs. J. Transl. Med. 2011, 9, 47. Available online: https://www.ncbi.nlm.nih.gov/pubmed/21513579 (accessed on 20 March 2024). [CrossRef]
- Accarie, A.; l’Homme, B.; Benadjaoud, M.A.; Lim, S.K.; Guha, C.; Benderitter, M.; Tamarat, R.; Semont, A. Extracellular Vesicles Derived from Mesenchymal Stromal Cells Mitigate Intestinal Toxicity in a Mouse Model of Acute Radiation Syndrome. Stem Cell Res. Ther. 2020, 11, 371. Available online: https://www.ncbi.nlm.nih.gov/pubmed/32854778 (accessed on 20 March 2024). [CrossRef]
- Gupta, S.; Batra, S.; Jain, M. Antibody labeling with radioiodine and radiometals. Methods Mol. Biol. 2014, 1141, 147–157. [Google Scholar] [CrossRef]
- Hunter, W.M.; Greenwood, F.C. Preparation of Iodine-131 Labelled Human Growth Hormone of High Specific Activity. Nature 1962, 194, 495–496. Available online: https://www.ncbi.nlm.nih.gov/pubmed/14450081 (accessed on 20 March 2024). [CrossRef]
- Greenwood, F.C.; Hunter, W.M.; Glover, J.S. The Preparation of I-131-Labelled Human Growth Hormone of High Specific Radioactivity. Biochem. J. 1963, 89, 114–123. Available online: https://www.ncbi.nlm.nih.gov/pubmed/14097352 (accessed on 20 March 2024). [CrossRef]
- Boonkitticharoen, V.; Laohathai, K. Assessing performances of Iodogen-coated surfaces used for radioiodination of proteins. Nucl. Med. Commun. 1990, 11, 295–304. [Google Scholar] [CrossRef]
- Markwell, M.A.; Fox, C.F. Surface-Specific Iodination of Membrane Proteins of Viruses and Eucaryotic Cells Using 1,3,4,6-tetrachloro-3alpha,6alpha-diphenylglycoluril. Biochemistry 1978, 17, 4807–4817. Available online: https://www.ncbi.nlm.nih.gov/pubmed/215191 (accessed on 20 March 2024). [CrossRef]
- Visser, G.W.; Klok, R.P.; Gebbinck, J.W.; ter Linden, T.; van Dongen, G.A.; Molthoff, C.F. Optimal Quality (131)I-Monoclonal Antibodies on High-Dose Labeling in a Large Reaction Volume and Temporarily Coating the Antibody with IODO-GEN. J. Nucl. Med. 2001, 42, 509–519. Available online: https://www.ncbi.nlm.nih.gov/pubmed/11337531 (accessed on 20 March 2024).
- Varga, Z.; Gyurko, I.; Paloczi, K.; Buzas, E.I.; Horvath, I.; Hegedus, N.; Mathe, D.; Szigeti, K. Radiolabeling of Extracellular Vesicles with (99m)Tc for Quantitative In Vivo Imaging Studies. Cancer Biother. Radiopharm. 2016, 31, 168–173. Available online: https://www.ncbi.nlm.nih.gov/pubmed/27310303 (accessed on 20 March 2024). [CrossRef]
- Jung, K.O.; Kim, Y.H.; Chung, S.J.; Lee, C.H.; Rhee, S.; Pratx, G.; Chung, J.K.; Youn, H. Identification of Lymphatic and Hematogenous Routes of Rapidly Labeled Radioactive and Fluorescent Exosomes through Highly Sensitive Multimodal Imaging. Int. J. Mol. Sci. 2020, 21, 7850. Available online: https://www.ncbi.nlm.nih.gov/pubmed/33105908 (accessed on 20 March 2024). [CrossRef]
- Khan, A.A.; Man, F.; Faruqu, F.N.; Kim, J.; Al-Salemee, F.; Carrascal-Minino, A.; Volpe, A.; Liam-Or, R.; Simpson, P.; Fruhwirth, G.O.; et al. PET Imaging of Small Extracellular Vesicles via [(89)Zr]Zr(oxinate)(4) Direct Radiolabeling. Bioconjug. Chem. 2022, 33, 473–485. Available online: https://www.ncbi.nlm.nih.gov/pubmed/35224973 (accessed on 20 March 2024). [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| CD73 Activity | Particle Size | |
|---|---|---|
| No labeling | 100% | 131.25 ± 15.91 nm |
| Chloramine T | 9.7 ± 3.1% | 148.39 ± 11.85 nm |
| Pierce | 66.6 ± 1.9% | 132.57 ± 27.9 nm |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C.-T.; Lai, R.C.; Phua, V.J.X.; Aw, S.E.; Zhang, B.; Sim, W.K.; Lim, S.K.; Ng, D.C.E. Standard Radio-Iodine Labeling Protocols Impaired the Functional Integrity of Mesenchymal Stem/Stromal Cell Exosomes. Int. J. Mol. Sci. 2024, 25, 3742. https://doi.org/10.3390/ijms25073742
Yang C-T, Lai RC, Phua VJX, Aw SE, Zhang B, Sim WK, Lim SK, Ng DCE. Standard Radio-Iodine Labeling Protocols Impaired the Functional Integrity of Mesenchymal Stem/Stromal Cell Exosomes. International Journal of Molecular Sciences. 2024; 25(7):3742. https://doi.org/10.3390/ijms25073742
Chicago/Turabian StyleYang, Chang-Tong, Ruenn Chai Lai, Vanessa Jing Xin Phua, Swee Eng Aw, Bin Zhang, Wei Kian Sim, Sai Kiang Lim, and David Chee Eng Ng. 2024. "Standard Radio-Iodine Labeling Protocols Impaired the Functional Integrity of Mesenchymal Stem/Stromal Cell Exosomes" International Journal of Molecular Sciences 25, no. 7: 3742. https://doi.org/10.3390/ijms25073742
APA StyleYang, C. -T., Lai, R. C., Phua, V. J. X., Aw, S. E., Zhang, B., Sim, W. K., Lim, S. K., & Ng, D. C. E. (2024). Standard Radio-Iodine Labeling Protocols Impaired the Functional Integrity of Mesenchymal Stem/Stromal Cell Exosomes. International Journal of Molecular Sciences, 25(7), 3742. https://doi.org/10.3390/ijms25073742