
Neutrophils and Anesthetic Drugs: Implications in Onco-Anesthesia


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Neutrophils’ Functions
2.1. Phagocytosis
2.2. Degranulation
2.3. ROS
2.4. Neutrophil Extracellular Traps (NETs)
2.4.1. Formation of NETs
2.4.2. Assessment of NET Formation
2.4.3. Clinical Implications of NETs
NETs in Critical Patients
NETs and Cancer and Cancer Associated Thrombosis
3. Cytokine Release
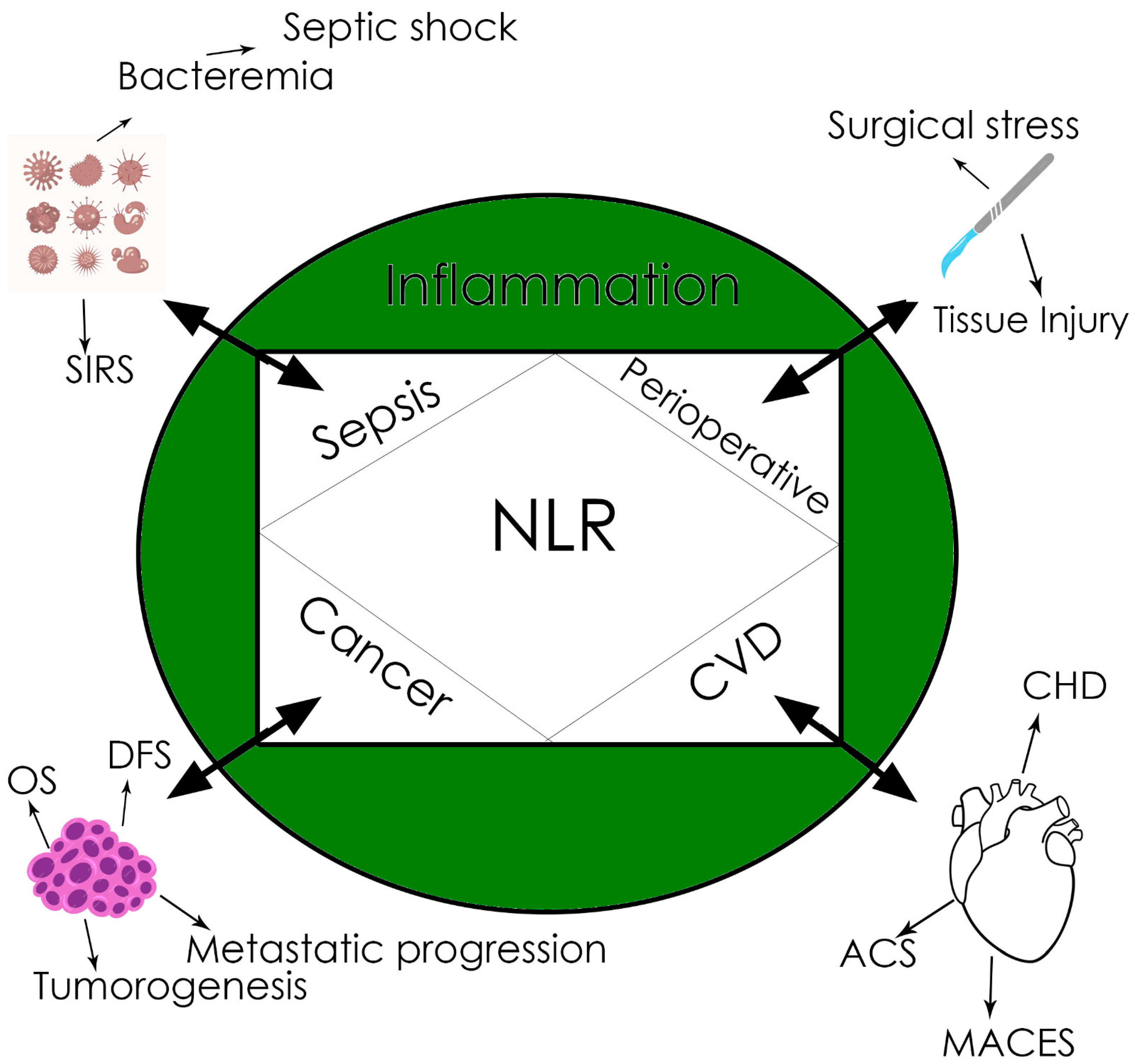
4. Neutrophil-to-Lymphocyte Ratio (NLR)
The Role of Neutrophils in Cancer
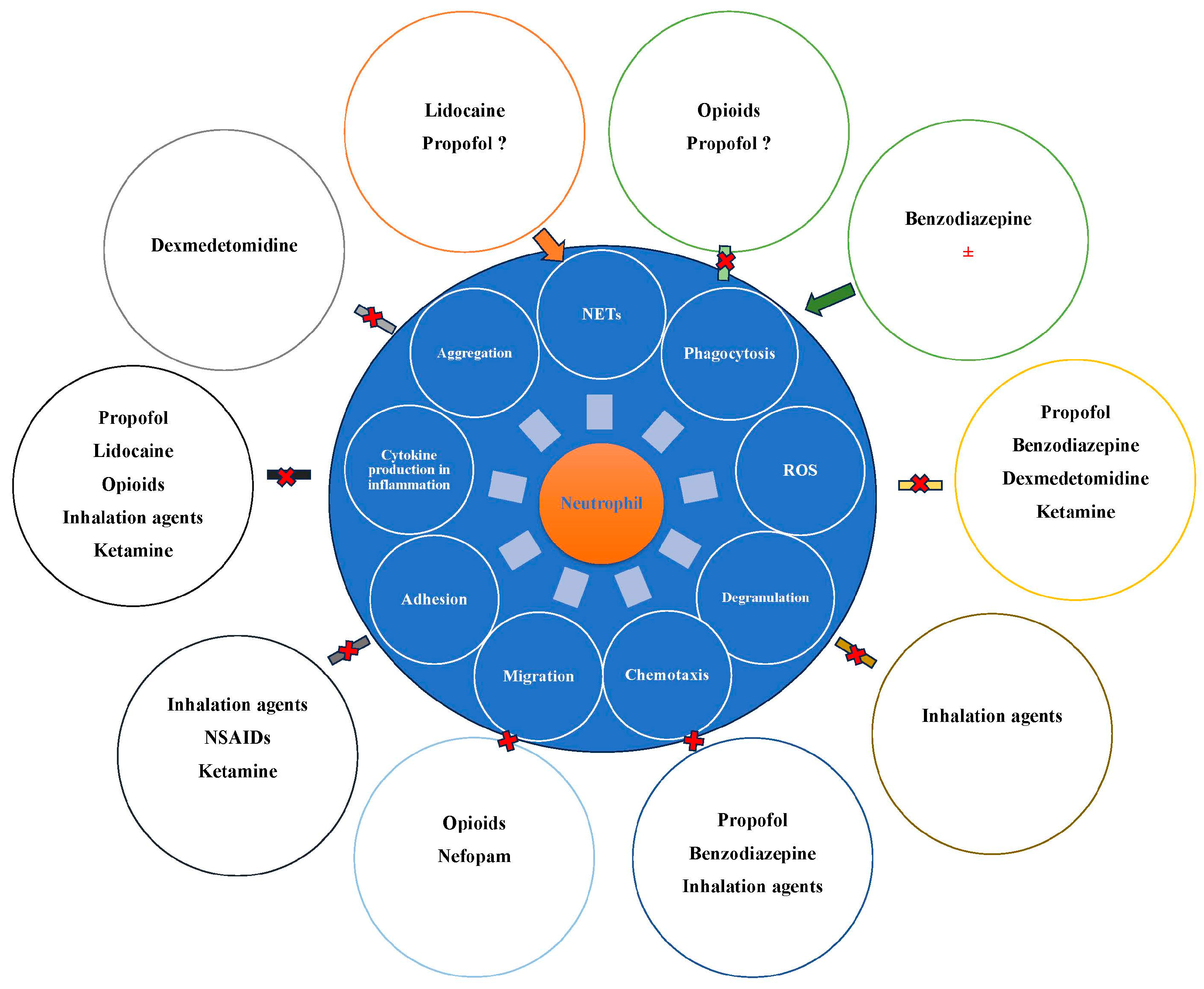
5. The Influence of Anesthetic Drugs on Neutrophils and Their Functions
5.1. Intravenous Anesthetic Agents and Neutrophils
5.1.1. Propofol
5.1.2. Ketamine
5.1.3. Benzodiazepines
5.1.4. Alpha-2 Agonists
5.1.5. Inhalation Agents and Neutrophils
5.1.6. Local Anesthetics (LAs)
5.1.7. Opioids
5.2. Minor Analgesics and Neutrophils
5.2.1. NSAIDs/Aspirin
5.2.2. Nefopam
5.2.3. Impacts on Short- and Long-Term Outcomes
5.3. Anesthetic Drugs and the NLR
5.4. Anesthetic Agents and the Efficacy of Anti-Cancer Drugs Administered after Surgery
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mestas, J.; Hughes, C.C. Of mice and not men: Differences between mouse and human immunology. J. Immunol. 2004, 72, 2731–2738. [Google Scholar] [CrossRef]
- Rosales, C. Neutrophils at the crossroads of innate and adaptive immunity. J. Leukoc. Biol. 2020, 108, 377–396. [Google Scholar] [CrossRef]
- Rosales, C. Neutrophil: A cell with many roles in inflammation or several cell types? Front. Physiol. 2018, 9, 113. [Google Scholar] [CrossRef] [PubMed]
- Chavakis, E.; Choi, E.Y.; Chavakis, T. Novel aspects in the regulation of the leukocyte adhesion cascade. Thromb. Haemost. 2009, 102, 191–197. [Google Scholar] [CrossRef]
- Ley, K.; Laudanna, C.; Cybulsky, M.I.; Nourshargh, S. Getting to the site of inflammation: The leukocyte adhesion cascade updated. Nat. Rev. Immunol. 2007, 7, 678–689. [Google Scholar] [CrossRef]
- Borregaard, N. Neutrophils, from marrow to microbes. Immunity 2010, 33, 657–670. [Google Scholar] [CrossRef] [PubMed]
- Nauseef, W.M. How human neutrophils kill and degrade microbes: An integrated view. Immunol. Rev. 2007, 219, 88–102. [Google Scholar] [CrossRef] [PubMed]
- Burn, G.L.; Foti, A.; Marsman, G.; Patel, D.F.; Zychlinsky, A. The neutrophil. Immunity 2021, 54, 1377–1391. [Google Scholar] [CrossRef]
- Mayadas, T.N.; Cullere, X.; Lowell, C.A. The multifaceted functions of neutrophils. Annu. Rev. Pathol. 2014, 9, 181–218. [Google Scholar] [CrossRef]
- Rosales, C.; Uribe-Querol, E. Phagocytosis: A fundamental process in immunity. Biomed. Res. Int. 2017, 2017, 9042851. [Google Scholar] [CrossRef]
- Brinkmann, V. Neutrophil extracellular traps in the second decade. J. Innate Immun. 2018, 10, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Papayannopoulos, V. Neutrophil extracellular traps in immunity and disease. Nat. Rev. Immunol. 2018, 18, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Malech, H.L.; Deleo, F.R.; Quinn, M.T. The role of neutrophils in the immune system: An overview. Methods Mol. Biol. 2014, 1124, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Freeman, S.A.; Grinstein, S. Phagocytosis: Receptors, signal integration, and the cytoskeleton. Immunol. Rev. 2014, 262, 193–215. [Google Scholar] [CrossRef]
- Lim, J.J.; Grinstein, S.; Roth, Z. Diversity and versatility of phagocytosis: Roles in innate immunity, tissue remodeling, and homeostasis. Front. Cell Infect. Microbiol. 2017, 7, 191. [Google Scholar] [CrossRef] [PubMed]
- Ramadass, M.; Catz, S.D. Molecular mechanisms regulating secretory organelles and endosomes in neutrophils and their implications for inflammation. Immunol. Rev. 2016, 273, 249–265. [Google Scholar] [CrossRef] [PubMed]
- Mol, S.; Hafkamp, F.M.J.; Varela, L.; Simkhada, N.; Taanman-Kueter, E.W.; Tas, S.W.; Wauben, M.H.M.; Kormelink, T.G.; de Jong, E.C. Efficient neutrophil activation requires two simultaneous activating stimuli. Int. J. Mol. Sci. 2021, 22, 10106. [Google Scholar] [CrossRef] [PubMed]
- El-Benna, J.; Hurtado-Nedelec, M.; Marzaioli, V.; Marie, J.C.; Gougerot-Pocidalo, M.A.; Dang, P.M. Priming of the neutrophil respiratory burst: Role in host defense and inflammation. Immunol. Rev. 2016, 273, 180–193. [Google Scholar] [CrossRef]
- Nguyen, G.T.; Green, E.R.; Mecsas, J. Neutrophils to the ROScue: Mechanisms of NADPH oxidase activation and bacterial resistance. Front. Cell. Infect. Microbiol. 2017, 7, 373. [Google Scholar] [CrossRef]
- Potera, R.M.; Jensen, M.J.; Hilkin, B.M.; South, G.K.; Hook, J.S.; Gross, E.A.; Moreland, J.G. Neutrophil azurophilic granule exocytosis is primed by TNF-α and partially regulated by NADPH oxidase. Innate Immun. 2016, 22, 635–646. [Google Scholar] [CrossRef]
- Wolach, B.; Gavrieli, R.; de Boer, M.; van Leeuwen, K.; Berger-Achituv, S.; Stauber, T.; Ben Ari, J.; Rottem, M.; Schlesinger, Y.; Grisaru-Soen, G.; et al. Chronic granulomatous disease: Clinical, functional, molecular, and genetic studies. The Israeli experience with 84 patients. Am. J. Hematol. 2017, 92, 28–36. [Google Scholar] [CrossRef]
- Roos, D. Chronic granulomatous disease. Br. Med. Bull. 2016, 118, 50–63. [Google Scholar] [CrossRef] [PubMed]
- Veenith, T.; Martin, H.; Le Breuilly, M.; Whitehouse, T.; Gao-Smith, F.; Duggal, N.; Lord, J.M.; Mian, R.; Sarphie, D.; Moss, P. High generation of reactive oxygen species from neutrophils in patients with severe COVID-19. Sci. Rep. 2022, 12, 10484. [Google Scholar] [CrossRef]
- Dupré-Crochet, S.; Erard, M.; Nüβe, O. ROS production in phagocytes: Why, when, and where? J. Leukoc. Biol. 2013, 94, 657–670. [Google Scholar] [CrossRef]
- Winterbourn, C.C.; Kettle, A.J.; Hampton, M.B. Reactive Oxygen species and neutrophil function. Annu. Rev. Biochem. 2016, 85, 765–792. [Google Scholar] [CrossRef]
- Herb, M.; Schramm, M. Functions of ROS in macrophages and antimicrobial immunity. Antioxidants 2021, 10, 313. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Suh, J.K.; Jeong, J.S.; Cho, S.Y.; Kim, D.W. Antioxidant effect of lidocaine and procaine on reactive oxygen species-induced endothelial dysfunction in the rabbit abdominal aorta. Korean J. Anesthesiol. 2010, 59, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Papayannopoulos, V.; Metzler, K.D.; Hakkim, A.; Zychlinsky, A. Neutrophil elastase and myeloperoxidase regulate the formation of neutrophil extracellular traps. J. Cell Biol. 2010, 191, 677–691. [Google Scholar] [CrossRef]
- Kasprzycka, W.; Homa-Mlak, I.; Mlak, R.; Małecka-Massalska, T. Direct and indirect methods of evaluating the NETosis process. J. Pre-Clin. Clin. Res. 2019, 13, 50–56. [Google Scholar] [CrossRef]
- Efrimescu, C.I.; Buggy, P.M.; Buggy, D.J. Neutrophil extracellular trapping role in cancer, metastases, and cancer-related thrombosis: A narrative review of the current evidence base. Curr. Oncol. Rep. 2021, 23, 118. [Google Scholar] [CrossRef]
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P. Molecular mechanisms of cell death: Recommendations of the nomenclature committee on cell death 2018. Cell Death Differ. 2018, 25, 486–541. [Google Scholar] [CrossRef] [PubMed]
- Yipp, B.G.; Kubes, P. NETosis: How vital is it? Blood 2013, 122, 2784–2794. [Google Scholar] [CrossRef] [PubMed]
- Ravindran, M.; Khan, M.A.; Palaniyar, N. Neutrophil extracellular trap formation: Physiology, pathology, and pharmacology. Biomolecules 2019, 9, 365. [Google Scholar] [CrossRef]
- Khan, M.A.; Palaniyar, N. Transcriptional firing helps to drive NETosis. Sci. Rep. 2017, 7, 41749. [Google Scholar] [CrossRef] [PubMed]
- Stoimenou, M.; Tzoros, G.; Skendros, P.; Chrysanthopoulou, A. Methods for the assessment of NET formation: From neutrophil biology to translational research. Int. J. Mol. Sci. 2022, 23, 15823. [Google Scholar] [CrossRef]
- De Buhr, N.; Von Köckritz-Blickwede, M. How neutrophil extracellular traps become visible. J. Immunol. Res. 2016, 2016, 4604713. [Google Scholar] [CrossRef]
- Condello, M.; Caraglia, M.; Castellano, M.; Arancia, G.; Meschini, S. Structural and functional alterations of cellular components as revealed by electron microscopy. Microsc. Res. Tech. 2013, 76, 1057–1069. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Chan, D.W.; Zaal, K.J.; Kaplan, M.J. A high-throughput real-time imaging technique to quantify NETosis and distinguish mechanisms of cell death in human neutrophils. J. Immunol. 2018, 200, 869–879. [Google Scholar] [CrossRef]
- Denning, N.L.; Aziz, M.; Gurien, S.D.; Wang, P. DAMPs and NETs in sepsis. Front. Immunol. 2019, 10, 2536. [Google Scholar] [CrossRef]
- Mikacenic, C.; Moore, R.; Dmyterko, V.; West, T.E.; Altemeier, W.A.; Liles, W.C.; Lood, C. Neutrophil extracellular traps (NETs) are increased in the alveolar spaces of patients with ventilator-associated pneumonia. Crit. Care 2018, 22, 358. [Google Scholar] [CrossRef]
- Lefrançais, E.; Mallavia, B.; Zhuo, H.; Calfee, C.S.; Looney, M.R. Maladaptive role of neutrophil extracellular traps in pathogen-induced lung injury. JCI Insight 2018, 3, e98178. [Google Scholar] [CrossRef] [PubMed]
- Maruchi, Y.; Tsuda, M.; Mori, H.; Takenaka, N.; Gocho, T.; Huq, M.A.; Takeyama, N. Plasma myeloperoxidase-conjugated DNA level predicts outcomes and organ dysfunction in patients with septic shock. Crit. Care 2018, 22, 176. [Google Scholar] [CrossRef] [PubMed]
- Szturmowicz, M.; Demkow, U. Neutrophil extracellular traps (NETs) in severe SARS-CoV-2 lung disease. Int. J. Mol. Sci. 2021, 22, 8854. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Jin, J. Neutrophil extracellular traps: New players in cancer research. Front. Immunol. 2022, 13, 937565. [Google Scholar] [CrossRef] [PubMed]
- Hahn, J.; Knopf, J.; Maueröder, C.; Kienhöfer, D.; Leppkes, M.; Herrmann, M. Neutrophils and neutrophil extracellular traps orchestrate initiation and resolution of inflammation. Clin. Exp. Rheumatol. 2016, 34, 6–8. [Google Scholar]
- Kaltenmeier, C.; Yazdani, H.O.; Morder, K.; Geller, D.A.; Simmons, R.L.; Tohme, S. Neutrophil extracellular traps promote T cell exhaustion in the tumor microenvironment. Front. Immunol. 2021, 12, 785222. [Google Scholar] [CrossRef]
- Jiang, Z.Z.; Peng, Z.P.; Liu, X.C.; Guo, H.F.; Zhou, M.M.; Jiang, D.; Ning, W.R.; Huang, Y.F.; Zheng, L.; Wu, Y. Neutrophil extracellular traps induce tumor metastasis through dual effects on cancer and endothelial cells. Oncoimmunology 2022, 11, 2052418. [Google Scholar] [CrossRef]
- Thålin, C.; Lundström, S.; Seignez, C.; Daleskog, M.; Lundström, A.; Henriksson, P.; Helleday, T.; Phillipson, M.; Wallén, H.; Demers, M. Citrullinated histone H3 as a novel prognostic blood marker in patients with advanced cancer. PLoS ONE 2018, 13, e0191231. [Google Scholar] [CrossRef]
- Snoderly, H.T.; Boone, B.A.; Bennewitz, M.F. Neutrophil extracellular traps in breast cancer and beyond: Current perspectives on NET stimuli, thrombosis and metastasis, and clinical utility for diagnosis and treatment. Breast Cancer Res. 2019, 21, 145. [Google Scholar] [CrossRef]
- Albrengues, J.; Shields, M.A.; Ng, D.; Park, C.G.; Ambrico, A.; Poindexter, M.E.; Upadhyay, P.; Uyeminami, D.L.; Pommier, A.; Küttner, V.; et al. Neutrophil extracellular traps produced during inflammation awaken dormant cancer cells in mice. Science 2018, 361, eaao4227. [Google Scholar] [CrossRef]
- Lee, W.; Ko, S.Y.; Mohamed, M.S.; Kenny, H.A.; Lengyel, E.; Naora, H. Neutrophils facilitate ovarian cancer premetastatic niche formation in the omentum. J. Exp. Med. 2019, 216, 176–194. [Google Scholar] [CrossRef] [PubMed]
- Cristinziano, L.; Modestino, L.; Antonelli, A.; Marone, G.; Simon, H.U.; Varricchi, G.; Galdiero, M.R. Neutrophil extracellular traps in cancer. Semin. Cancer Biol. 2022, 79, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Wysocki, R.W.; Amoozgar, Z.; Maiorino, L.; Fein, M.R.; Jorns, J.; Schott, A.F.; Kinugasa-Katayama, Y.; Lee, Y.; Won, N.H.; et al. Cancer cells induce metastasis-supporting neutrophil extracellular DNA traps. Sci. Transl. Med. 2016, 8, 361ra138. [Google Scholar] [CrossRef]
- Yang, L.; Liu, Q.; Zhang, X.; Liu, X.; Zhou, B.; Chen, J.; Huang, D.; Li, J.; Li, H.; Chen, F.; et al. DNA of neutrophil extracellular traps promotes cancer metastasis via CCDC25. Nature 2020, 583, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Cong, M.; Li, J.; He, D.; Wu, Q.; Tian, P.; Wang, Y.; Yang, S.; Liang, C.; Liang, Y.; et al. Cathepsin C promotes breast cancer lung metastasis by modulating neutrophil infiltration and neutrophil extracellular trap formation. Cancer Cell 2021, 39, 423–437.e7. [Google Scholar] [CrossRef] [PubMed]
- Tamassia, N.; Bianchetto-Aguilera, F.; Arruda-Silva, F.; Gardiman, E.; Gasperini, S.; Calzetti, F.; Cassatella, M.A. Cytokine production by human neutrophils: Revisiting the “dark side of the moon”. Eur. J. Clin. Investig. 2018, 48, e12952. [Google Scholar] [CrossRef] [PubMed]
- Tecchio, C.; Scapini, P.; Pizzolo, G.; Cassatella, M.A. On the cytokines produced by human neutrophils in tumors. Semin. Cancer Biol. 2013, 23, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Gabrilovich, D. Cytokine production by neutrophils. In The Neutrophils, 3rd ed.; de Chaisemartin, L., Cassatella, M., Chollet-Martin, S., Eds.; Imperial College Press: London, UK, 2013; pp. 189–217. [Google Scholar]
- Buonacera, A.; Stancanelli, B.; Colaci, M.; Malatino, L. Neutrophil to lymphocyte ratio: An emerging marker of the relationships between the immune system and diseases. Int. J. Mol. Sci. 2022, 23, 3636. [Google Scholar] [CrossRef]
- Jiang, J.; Liu, R.; Yu, X.; Yang, R.; Xu, H.; Mao, Z.; Wang, Y. The neutrophil-lymphocyte count ratio as a diagnostic marker for bacteraemia: A systematic review and meta-analysis. Am. J. Emerg. Med. 2019, 37, 1482–1489. [Google Scholar] [CrossRef]
- Lee, M.J.; Park, S.D.; Kwon, S.W.; Woo, S.I.; Lee, M.D.; Shin, S.H.; Kim, D.H.; Kwan, J.; Park, K.S. Relation between neutrophil-to-lymphocyte ratio and index of microcirculatory resistance in patients with ST-segment elevation myocardial infarction undergoing primary percutaneous coronary intervention. Am. J. Cardiol. 2016, 118, 1323–1328. [Google Scholar] [CrossRef]
- Huang, Z.; Fu, Z.; Huang, W.; Huang, K. Prognostic value of neutrophil-to-lymphocyte ratio in sepsis: A meta-analysis. Am. J. Emerg. Med. 2020, 38, 641–647. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.Y.; Yoo, G.; Lee, T.; Uh, Y.; Kim, J. Identification of the robust predictor for sepsis based on clustering analysis. Sci. Rep. 2022, 12, 2336. [Google Scholar] [CrossRef] [PubMed]
- Hibino, S.; Kawazoe, T.; Kasahara, H.; Itoh, S.; Ishimoto, T.; Sakata-Yanagimoto, M.; Taniguchi, K. Inflammation-induced tumorigenesis and metastasis. Int. J. Mol. Sci. 2021, 22, 5421. [Google Scholar] [CrossRef] [PubMed]
- Templeton, A.J.; McNamara, M.G.; Šeruga, B.; Vera-Badillo, F.E.; Aneja, P.; Ocaña, A.; Leibowitz-Amit, R.; Sonpavde, G.; Knox, J.J.; Tran, B.; et al. Prognostic role of neutrophil-to-lymphocyte ratio in solid tumors: A systematic review and meta-analysis. J. Natl. Cancer Inst. 2014, 106, dju124. [Google Scholar] [CrossRef]
- Malietzis, G.; Giacometti, M.; Kennedy, R.H.; Athanasiou, T.; Aziz, O.; Jenkins, J.T. The emerging role of neutrophil to lymphocyte ratio in determining colorectal cancer treatment outcomes: A systematic review and meta-analysis. Ann. Surg. Oncol. 2014, 21, 3938–3946. [Google Scholar] [CrossRef] [PubMed]
- Silberman, S.; Abu-Yunis, U.; Tauber, R.; Shavit, L.; Grenader, T.; Fink, D.; Bitran, D.; Merin, O. Neutrophil-lymphocyte ratio: Prognostic impact in heart surgery. Early Outcomes Late Survival. Ann. Thorac. Surg. 2018, 105, 581–586. [Google Scholar] [CrossRef]
- Hajibandeh, S.; Hajibandeh, S.; Hobbs, N.; Mansour, M. Neutrophil-to-lymphocyte ratio predicts acute appendicitis and distinguishes between complicated and uncomplicated appendicitis: A systematic review and meta-analysis. Am. J. Surg. 2020, 219, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Coussens, L.M.; Werb, Z. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef]
- Schoppmann, S.F.; Birner, P.; Stöckl, J.; Kalt, R.; Ullrich, R.; Caucig, C.; Kriehuber, E.; Nagy, K.; Alitalo, K.; Kerjaschki, D. Tumor-associated macrophages express lymphatic endothelial growth factors and are related to peritumoral lymphangiogenesis. Am. J. Pathol. 2002, 161, 947–956. [Google Scholar] [CrossRef]
- Brandau, S.; Dumitru, C.A.; Lang, S. Protumor and antitumor functions of neutrophil granulocytes. Semin. Immunopathol. 2013, 35, 163–176. [Google Scholar] [CrossRef]
- Uribe-Querol, E.; Rosales, C. Neutrophils in Cancer: Two Sides of the Same Coin. J. Immunol. Res. 2015, 2015, 983698. [Google Scholar] [CrossRef] [PubMed]
- Houghton, A.M.; Rzymkiewicz, D.M.; Ji, H.; Gregory, A.D.; Egea, E.E.; Metz, H.E.; Stolz, D.B.; Land, S.R.; Marconcini, L.A.; Kliment, C.R.; et al. Neutrophil elastase-mediated degradation of IRS-1 accelerates lung tumor growth. Nat. Med. 2010, 16, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Zilio, S.; Serafini, P. Neutrophils and granulocytic MDSC: The janus God of cancer immunotherapy. Vaccines 2016, 4, 31. [Google Scholar] [CrossRef] [PubMed]
- Condamine, T.; Ramachandran, I.; Youn, J.I.; Gabrilovich, D.I. Regulation of tumor metastasis by myeloid-derived suppressor cells. Annu. Rev. Med. 2015, 66, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Tecchio, C.; Micheletti, A.; Cassatella, M.A. Neutrophil-derived cytokines: Facts beyond expression. Front. Immunol. 2014, 5, 508. [Google Scholar] [CrossRef] [PubMed]
- Wigmore, T.J.; Mohammed, K.; Jhanji, S. Long-term survival for patients undergoing volatile versus iv anesthesia for cancer surgery: A retrospective analysis. Anesthesiology 2016, 124, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Yap, A.; Lopez-Olivo, M.A.; Dubowitz, J.; Hiller, J.; Riedel, B. Global onco-anesthesia research collaboration group. In reply: Comment. on “Anesthetic technique and cancer outcomes: A meta-analysis of total intravenous versus volatile anesthesia”. Can. J. Anaesth. 2020, 67, 152–153. [Google Scholar] [CrossRef] [PubMed]
- Hasselager, R.P.; Hallas, J.; Gögenur, I. Inhalation or total intravenous anaesthesia and recurrence after colorectal cancer surgery: A propensity score matched Danish registry-based study. Br. J. Anaesth. 2021, 126, 921–930. [Google Scholar] [CrossRef]
- Che, X.; Li, T. Total versus inhaled intravenous anesthesia methods for prognosis of patients with lung, breast, or esophageal cancer: A cohort study. Front. Surg. 2023, 10, 1155351. [Google Scholar] [CrossRef]
- Meier, A.; Nizet, V. Impact of anesthetics on human neutrophil function. Anesth. Analg. 2019, 128, 569–574. [Google Scholar] [CrossRef]
- Lingamaneni, R.; Krasowski, M.D.; Jenkins, A.; Truong, T.; Giunta, A.L.; Blackbeer, J.; MacIver, M.B.; Harrison, N.L.; Hemmings, H.C., Jr. Anesthetic properties of 4-iodopropofol: Implications for mechanisms of anesthesia. Anesthesiology 2001, 94, 1050–1057. [Google Scholar] [CrossRef]
- Yi, S.; Tao, X.; Wang, Y.; Cao, Q.; Zhou, Z.; Wang, S. Effects of propofol on macrophage activation and function in diseases. Front. Pharmacol. 2022, 13, 964771. [Google Scholar] [CrossRef] [PubMed]
- Bhat, R.; Axtell, R.; Mitra, A.; Miranda, M.; Lock, C.; Tsien, R.W.; Steinman, L. Inhibitory role for GABA in autoimmune inflammation. Proc. Natl. Acad. Sci. USA 2010, 107, 2580–2585. [Google Scholar] [CrossRef]
- Haeseler, G.; Leuwer, M. High-affinity block of voltage-operated rat IIA neuronal sodium channels by 2,6 di-tert-butylphenol, a propofol analogue. Eur. J. Anaesthesiol. 2003, 20, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Kotani, Y.; Shimazawa, M.; Yoshimura, S.; Iwama, T.; Hara, H. The experimental and clinical pharmacology of propofol, an anesthetic agent with neuroprotective properties. CNS Neurosci. Ther. 2008, 14, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Krumholz, W.; Endrass, J.; Hempelmann, G. Propofol inhibits phagocytosis and killing of Staphylococcus aureus and Escherichia coli by polymorphonuclear leukocytes in vitro. Can. J. Anaesth. 1994, 41, 446–449. [Google Scholar] [CrossRef] [PubMed]
- Heller, A.; Heller, S.; Blecken, S.; Urbaschek, R.; Koch, T. Effects of intravenous anesthetics on bacterial elimination in human blood in vitro. Acta Anaesthesiol. Scand. 1998, 42, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Ionescu, D.C.; Margarit, S.C.; Hadade, A.N.; Mocan, T.N.; Miron, N.A.; Sessler, D.I. Choice of anesthetic technique on plasma concentrations of interleukins and cell adhesion molecules. Perioper. Med. 2013, 2, 8. [Google Scholar] [CrossRef] [PubMed]
- Koo, B.W.; Sim, J.B.; Shin, H.J.; Kim, D.W.; Kang, S.B.; Do, S.H.; Na, H.S. Surgical site infection after colorectal surgery according to the main anesthetic agent: A retrospective comparison between volatile anesthetics and propofol. Korean J. Anesthesiol. 2016, 69, 332–340. [Google Scholar] [CrossRef]
- Von Dossow, V.; Baur, S.; Sander, M.; Tønnesen, H.; Marks, C.; Paschen, C.; Berger, G.; Spies, C.D. Propofol increased the interleukin-6 to interleukin-10 ratio more than isoflurane after surgery in long-term alcoholic patients. J. Int. Med. Res. 2007, 35, 395–405. [Google Scholar] [CrossRef]
- Hughes, C.G.; Mailloux, P.T.; Devlin, J.W.; Swan, J.T.; Sanders, R.D.; Anzueto, A.; Jackson, J.C.; Hoskins, A.S.; Pun, B.T.; Orun, O.M.; et al. Dexmedetomidine or propofol for sedation in mechanically ventilated adults with sepsis. N. Engl. J. Med. 2021, 384, 1424–1436. [Google Scholar] [CrossRef]
- Sanders, R.D.; Grover, V.; Goulding, J.; Godlee, A.; Gurney, S.; Snelgrove, R.; Ma, D.; Singh, S.; Maze, M.; Hussell, T. Immune cell expression of GABAA receptors and the effects of diazepam on influenza infection. J. Neuroimmunol. 2015, 282, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.C.; Chung, P.J.; Ho, C.M.; Kuo, C.Y.; Hung, M.F.; Huang, Y.T.; Chang, W.Y.; Chang, Y.W.; Chan, K.H.; Hwang, T.L. Propofol inhibits superoxide production, elastase release, and chemotaxis in formyl peptide-activated human neutrophils by blocking formyl peptide receptor 1. J. Immunol. 2013, 190, 6511–5619. [Google Scholar] [CrossRef] [PubMed]
- Stahl, S.M. Mechanism of action of ketamine. CNS Spectr. 2013, 18, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.S.; Penna, A.; Orser, B.A. Ketamine increases the function of γ-aminobutyric acid type A receptors in hippocampal and cortical neurons. Anesthesiology 2017, 126, 666–677. [Google Scholar] [CrossRef]
- Zhou, C.; Douglas, J.E.; Kumar, N.N.; Shu, S.; Bayliss, D.A.; Chen, X. Forebrain HCN1 channels contribute to hypnotic actions of ketamine. Anesthesiology 2013, 118, 785–795. [Google Scholar] [CrossRef] [PubMed]
- Zilberstein, G.; Levy, R.; Rachinsky, M.; Fisher, A.; Greemberg, L.; Shapira, Y.; Appelbaum, A.; Roytblat, L. Ketamine attenuates neutrophil activation after cardiopulmonary bypass. Anesth. Analg. 2002, 95, 531–536. [Google Scholar] [CrossRef] [PubMed]
- Ghori, K.; O’Driscoll, J.; Shorten, G. The effect of midazolam on neutrophil mitogen-activated protein kinase. Eur. J. Anaesthesiol. 2010, 27, 562–565. [Google Scholar] [CrossRef]
- Marino, F.; Cattaneo, S.; Cosentino, M.; Rasini, E.; Di Grazia, L.; Fietta, A.M.; Lecchini, S.; Frigo, G. Diazepam stimulates migration and phagocytosis of human neutrophils: Possible contribution of peripheral-type benzodiazepine receptors and intracellular calcium. Pharmacology 2001, 63, 42–49. [Google Scholar] [CrossRef]
- Herrera-García, A.M.; Domínguez-Luis, M.J.; Arce-Franco, M.; Armas-González, E.; Álvarez de La Rosa, D.; Machado, J.D.; Pec, M.K.; Feria, M.; Barreiro, O.; Sánchez-Madrid, F.; et al. Prevention of neutrophil extravasation by α2-adrenoceptor-mediated endothelial stabilization. J. Immunol. 2014, 193, 3023–3035. [Google Scholar] [CrossRef]
- Chen, R.; Sun, Y.; Lv, J.; Dou, X.; Dai, M.; Sun, S.; Lin, Y. Effects of dexmedetomidine on immune cells: A narrative review. Front. Pharmacol. 2022, 13, 829951. [Google Scholar] [CrossRef] [PubMed]
- Carnet Le Provost, K.; Kepp, O.; Kroemer, G.; Bezu, L. Trial watch: Dexmedetomidine in cancer therapy. Oncoimmunology 2024, 13, 2327143. [Google Scholar] [CrossRef] [PubMed]
- Franks, N.P.; Lieb, W.R. Molecular and cellular mechanisms of general anaesthesia. Nature 1994, 367, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Liang, P.; Liu, J.; Ke, B.; Wang, X.; Li, F.; Li, T.; Bayliss, D.A.; Chen, X. HCN1 channels contribute to the effects of amnesia and hypnosis but not immobility of volatile anesthetics. Anesth. Analg. 2015, 121, 661–666. [Google Scholar] [CrossRef] [PubMed]
- Downie, D.L.; Hall, A.C.; Lieb, W.R.; Franks, N.P. Effects of inhalational general anaesthetics on native glycine receptors in rat medullary neurones and recombinant glycine receptors in Xenopus oocytes. Br. J. Pharmacol. 1996, 118, 493–502. [Google Scholar] [CrossRef]
- Stollings, L.M.; Jia, L.J.; Tang, P.; Dou, H.; Lu, B.; Xu, Y. Immune modulation by volatile anesthetics. Anesthesiology 2016, 125, 399–411. [Google Scholar] [CrossRef] [PubMed]
- Fröhlich, D.; Rothe, G.; Schwall, B.; Schmid, P.; Schmitz, G.; Taeger, K.; Hobbhahn, J. Effects of volatile anaesthetics on human neutrophil oxidative response to the bacterial peptide FMLP. Br. J. Anaesth. 1997, 78, 718–723. [Google Scholar] [CrossRef] [PubMed]
- Möbert, J.; Zahler, S.; Becker, B.F.; Conzen, P.F. Inhibition of neutrophil activation by volatile anesthetics decreases adhesion to cultured human endothelial cells. Anesthesiology 1999, 90, 1372–1381. [Google Scholar] [CrossRef] [PubMed]
- de Rossi, L.W.; Horn, N.A.; Hecker, K.E.; Robitzsch, T.; Hutschenreuter, G.; Rossaint, R. Effect of halothane and isoflurane on binding of ADP- and TRAP-6- activated platelets to leukocytes in Whole blood. Anesthesiology 2002, 96, 117–124. [Google Scholar] [CrossRef]
- Du, X.; Jiang, C.; Lv, Y.; Dull, R.O.; Zhao, Y.Y.; Schwartz, D.E.; Hu, G. Isoflurane promotes phagocytosis of apoptotic neutrophils through AMPK-mediated ADAM17/Mer signaling. PLoS ONE 2017, 12, e0180213. [Google Scholar] [CrossRef]
- Kong, H.Y.; Zhu, S.M.; Wang, L.Q.; He, Y.; Xie, H.Y.; Zheng, S.S. Sevoflurane protects against acute kidney injury in a small-size liver transplantation model. Am. J. Nephrol. 2010, 32, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Alam, A.; Rampes, S.; Patel, S.; Hana, Z.; Ma, D. Anesthetics or anesthetic techniques and cancer surgical outcomes: A possible link. Korean J. Anesthesiol. 2021, 74, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Buckley, A.; McQuaid, S.; Johnson, P.; Buggy, D.J. Effect of anaesthetic technique on the natural killer cell anti-tumour activity of serum from women undergoing breast cancer surgery: A pilot study. Br. J. Anaesth. 2014, 113, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Kim, R. Effects of surgery and anesthetic choice on immunosuppression and cancer recurrence. J. Transl. Med. 2018, 16, 8. [Google Scholar] [CrossRef] [PubMed]
- Tikhonov, D.B.; Zhorov, B.S. Mechanism of sodium channel block by local anesthetics, antiarrhythmics, and anticonvulsants. J. Gen. Physiol. 2017, 149, 465–481. [Google Scholar] [CrossRef] [PubMed]
- de Lera Ruiz, M.; Kraus, R.L. Voltage-gated sodium channels: Structure, function, pharmacology, and clinical indications. J. Med. Chem. 2015, 58, 7093–7118. [Google Scholar] [CrossRef]
- Kulińska, K.I.; Szałkowska, S.; Andrusiewicz, M.; Kotwicka, M.; Billert, H. The effect of local anaesthetics on apoptosis and NETosis of human neutrophils in vitro: Comparison between lidocaine and ropivacaine. Hum. Cell 2023, 36, 2027–2039. [Google Scholar] [CrossRef]
- Galoș, E.V.; Tat, T.F.; Popa, R.; Efrimescu, C.I.; Finnerty, D.; Buggy, D.J.; Ionescu, D.C.; Mihu, C.M. Neutrophil extracellular trapping and angiogenesis biomarkers after intravenous or inhalation anaesthesia with or without intravenous lidocaine for breast cancer surgery: A prospective, randomised trial. Br. J. Anaesth. 2020, 125, 712–721. [Google Scholar] [CrossRef]
- Kulinska, K.I.; Billert, M.; Sawinski, K.; Czerniak, K.; Gaca, M.; Kusza, K.; Nowak, K.W.; Siemionow, M.; Billert, H. Local anaesthetics upregulate nitric oxide generation in cord blood and adult human neutrophils. Sci. Rep. 2019, 9, 569. [Google Scholar] [CrossRef]
- Castro, I.; Carvalho, P.; Vale, N.; Monjardino, T.; Mourão, J. Systemic anti-inflammatory effects of intravenous lidocaine in surgical patients: A systematic review and meta-analysis. J. Clin. Med. 2023, 12, 3772. [Google Scholar] [CrossRef]
- Zhong, J.; Hu, J.; Mao, L.; Ye, G.; Qiu, K.; Zhao, Y.; Hu, S. Efficacy of intravenous lidocaine for pain relief in the emergency department: A systematic review and meta-analysis. Front. Med. 2022, 8, 706844. [Google Scholar] [CrossRef] [PubMed]
- Brejchova, J.; Holan, V.; Svoboda, P. Expression of opioid receptors in cells of the immune system. Int. J. Mol. Sci. 2020, 22, 315. [Google Scholar] [CrossRef] [PubMed]
- Boland, J.W.; Pockley, A.G. Influence of opioids on immune function in patients with cancer pain: From bench to bedside. Br. J. Pharmacol. 2018, 175, 2726–2736. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Ninkovic, J.; Banerjee, S.; Charboneau, R.G.; Das, S.; Dutta, R.; Kirchner, V.A.; Koodie, L.; Ma, J.; Meng, J.; et al. Opioid drug abuse and modulation of immune function: Consequences in the susceptibility to opportunistic infections. J. Neuroimmune Pharmacol. 2011, 6, 442–465. [Google Scholar] [CrossRef] [PubMed]
- Kelly, E.; Henderson, G.; Bailey, C.P. Emerging areas of opioid pharmacology. Br. J. Pharmacol. 2018, 175, 2715–2716. [Google Scholar] [CrossRef]
- Brogi, E.; Forfori, F. Anesthesia and cancer recurrence: An overview. J. Anesth. Analg. Crit. Care 2022, 2, 33. [Google Scholar] [CrossRef] [PubMed]
- Díaz-González, F.; Sánchez-Madrid, F. NSAIDs: Learning new tricks from old drugs. Eur. J. Immunol. 2015, 45, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Venuta, A.; Nasso, R.; Gisonna, A.; Iuliano, R.; Montesarchio, S.; Acampora, V.; Sepe, L.; Avagliano, A.; Arcone, R.; Arcucci, A.; et al. Celecoxib, a non-steroidal anti-inflammatory drug, exerts a toxic effect on human melanoma cells grown as 2D and 3D cell cultures. Life 2023, 13, 1067. [Google Scholar] [CrossRef] [PubMed]
- Tsoi, K.K.F.; Ho, J.M.W.; Chan, F.C.H.; Sung, J.J.Y. Long-term use of low-dose aspirin for cancer prevention: A 10-year population cohort study in Hong Kong. Int. J. Cancer 2019, 145, 267–273. [Google Scholar] [CrossRef]
- Hall, C.J.; Wicker, S.M.; Chien, A.T.; Tromp, A.; Lawrence, L.M.; Sun, X.; Krissansen, G.W.; Crosier, K.E.; Crosier, P.S. Repositioning drugs for inflammatory disease—Fishing for new anti-inflammatory agents. Dis. Model. Mech. 2014, 7, 1069–1081. [Google Scholar] [CrossRef]
- Sessler, D.; Pei, L.; Huang, Y.; Fleischmann, E.; Marhofer, P.; Kurz, A.; Mayers, D.B.; Meyer-Treschan, T.A.; Grady, M.; Tan, E.Y.; et al. Recurrence of breast cancer after regional or general anaesthesia: A randomised controlled trial. Lancet 2019, 394, 1807–1815. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, M.F.; Cata, J.P. Anesthesia techniques and long-term oncological outcomes. Front. Oncol. 2021, 11, 788918. [Google Scholar] [CrossRef] [PubMed]
- Murphy, O.; Forget, P.; Ma, D.; Buggy, D.J. Tumour excisional surgery, anaesthetic-analgesic techniques, and oncologic outcomes: A narrative review. Br. J. Anaesth. 2023, 131, 989–1001. [Google Scholar] [CrossRef] [PubMed]
- Ní Eochagáin, A.; Burns, D.; Riedel, B.; Sessler, D.I.; Buggy, D.J. The effect of anaesthetic technique during primary breast cancer surgery on neutrophil-lymphocyte ratio, platelet-lymphocyte ratio and return to intended oncological therapy. Anaesthesia 2018, 73, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Memary, E.; Mirkheshti, A.; Ghasemi, M.; Taheri, M.; Dolatabadi, A.; Kaboudvand, A. The effect of lidocaine infusion during general anesthesia on neutrophil-lymphocyte-ratio in breast cancer patient’s candidate for mastectomy; a clinical trial. J. Cell Mol. Anesth. 2016, 1, 146–153. [Google Scholar]
- Alexa, A.L.; Ciocan, A.; Zaharie, F.; Valean, D.; Sargarovschi, S.; Breazu, C.; Al Hajjar, N.; Ionescu, D. The influence of intravenous lidocaine infusion on postoperative outcome and neutrophil-to-lymphocyte ratio in colorectal cancer patients. A pilot study. J. Gastrointestin. Liver Dis. 2023, 32, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Domagalska, M.; Ciftsi, B.; Janusz, P.; Reysner, T.; Kolasinski, J.; Wieczorowska-Tobis, K.; Kowalski, G. The neutrophil-to-lymphocyte ratio (NLR) and platelet-to-lymphocyte ratio (PLR) levels following erector spinae plane block (ESPB) in posterior lumbar decompression: A randomized, controlled trial. Eur. Spine J. 2023, 32, 4192–4199. [Google Scholar] [CrossRef] [PubMed]
- Watson, J.; Ninh, M.K.; Ashford, S.; Cornett, E.M.; Kaye, A.D.; Urits, I.; Viswanath, O. Anesthesia medications and interaction with chemotherapeutic agents. Oncol. Ther. 2021, 9, 121–138. [Google Scholar] [CrossRef]
- Jiang, S.; Liu, Y.; Huang, L.; Zhang, F.; Kang, R. Effects of propofol on cancer development and chemotherapy: Potential mechanisms. Eur. J. Pharmacol. 2018, 831, 46–51. [Google Scholar] [CrossRef]
- Wang, P.; Chen, J.; Mu, L.H.; Du, Q.H.; Niu, X.H.; Zhang, M.Y. Propofol inhibits invasion and enhances paclitaxel-induced apoptosis in ovarian cancer cells through the suppression of the transcription factor slug. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 1722–1729, Erratum in: Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 6446. [Google Scholar]
- Xing, W.; Chen, D.T.; Pan, J.H.; Chen, Y.H.; Yan, Y.; Li, Q.; Xue, R.F.; Yuan, Y.F.; Zeng, W.A. Lidocaine induces apoptosis and suppresses tumor growth in human hepatocellular carcinoma cells in vitro and in a xenograft model in vivo. Anesthesiology 2017, 126, 868–881. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alexa, A.L.; Sargarovschi, S.; Ionescu, D. Neutrophils and Anesthetic Drugs: Implications in Onco-Anesthesia. Int. J. Mol. Sci. 2024, 25, 4033. https://doi.org/10.3390/ijms25074033
Alexa AL, Sargarovschi S, Ionescu D. Neutrophils and Anesthetic Drugs: Implications in Onco-Anesthesia. International Journal of Molecular Sciences. 2024; 25(7):4033. https://doi.org/10.3390/ijms25074033
Chicago/Turabian StyleAlexa, Alexandru Leonard, Sergiu Sargarovschi, and Daniela Ionescu. 2024. "Neutrophils and Anesthetic Drugs: Implications in Onco-Anesthesia" International Journal of Molecular Sciences 25, no. 7: 4033. https://doi.org/10.3390/ijms25074033
APA StyleAlexa, A. L., Sargarovschi, S., & Ionescu, D. (2024). Neutrophils and Anesthetic Drugs: Implications in Onco-Anesthesia. International Journal of Molecular Sciences, 25(7), 4033. https://doi.org/10.3390/ijms25074033