

Thermosensitive TRP Channels Are Functionally Expressed and Influence the Lipogenesis in Human Meibomian Gland Cells


 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
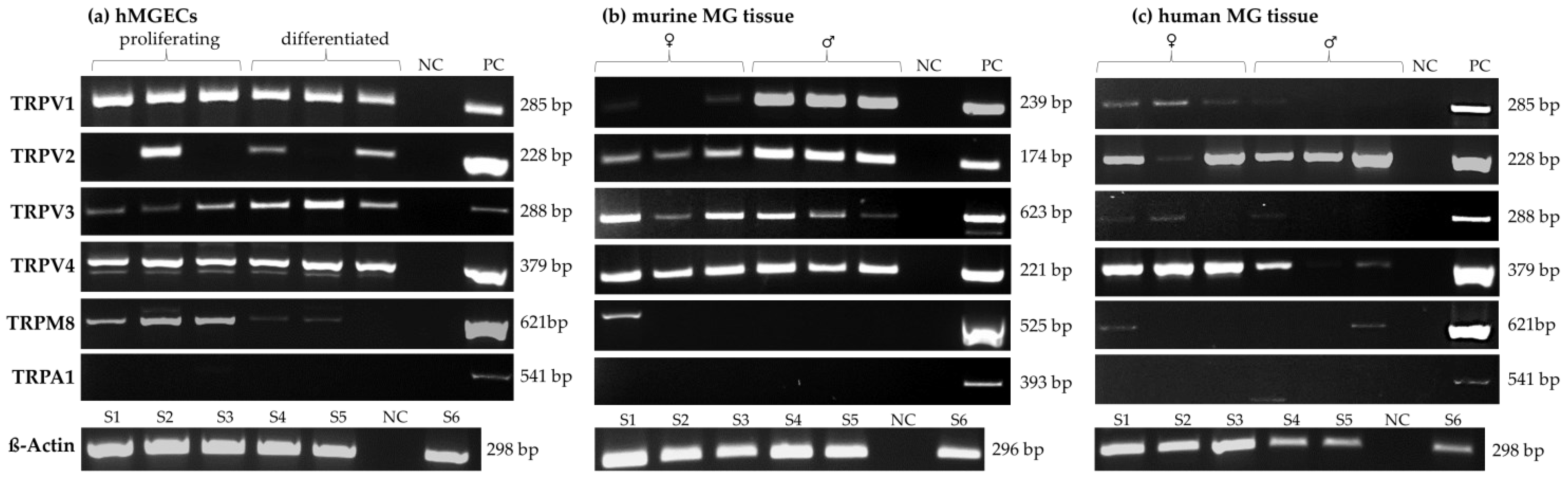
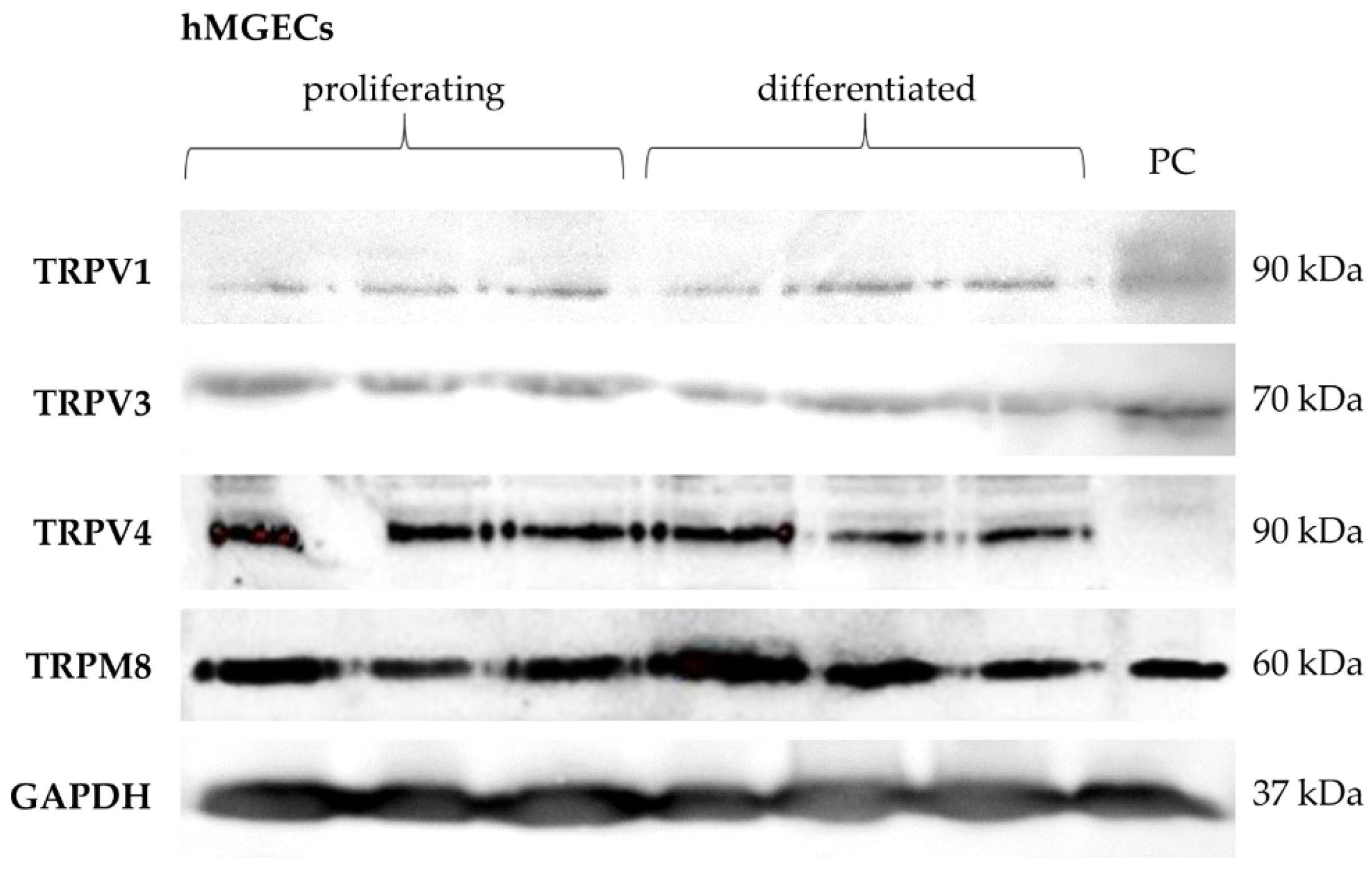
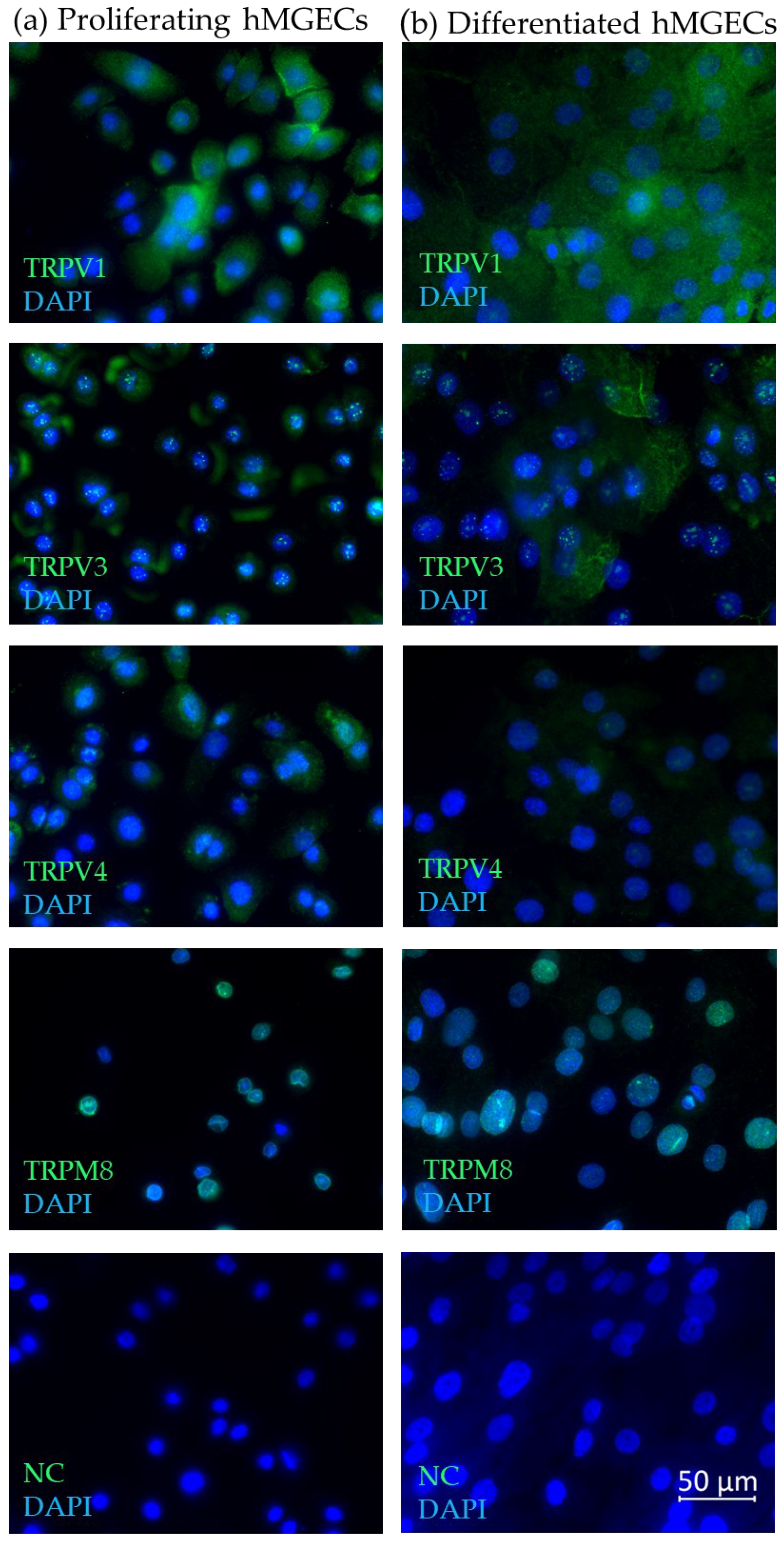
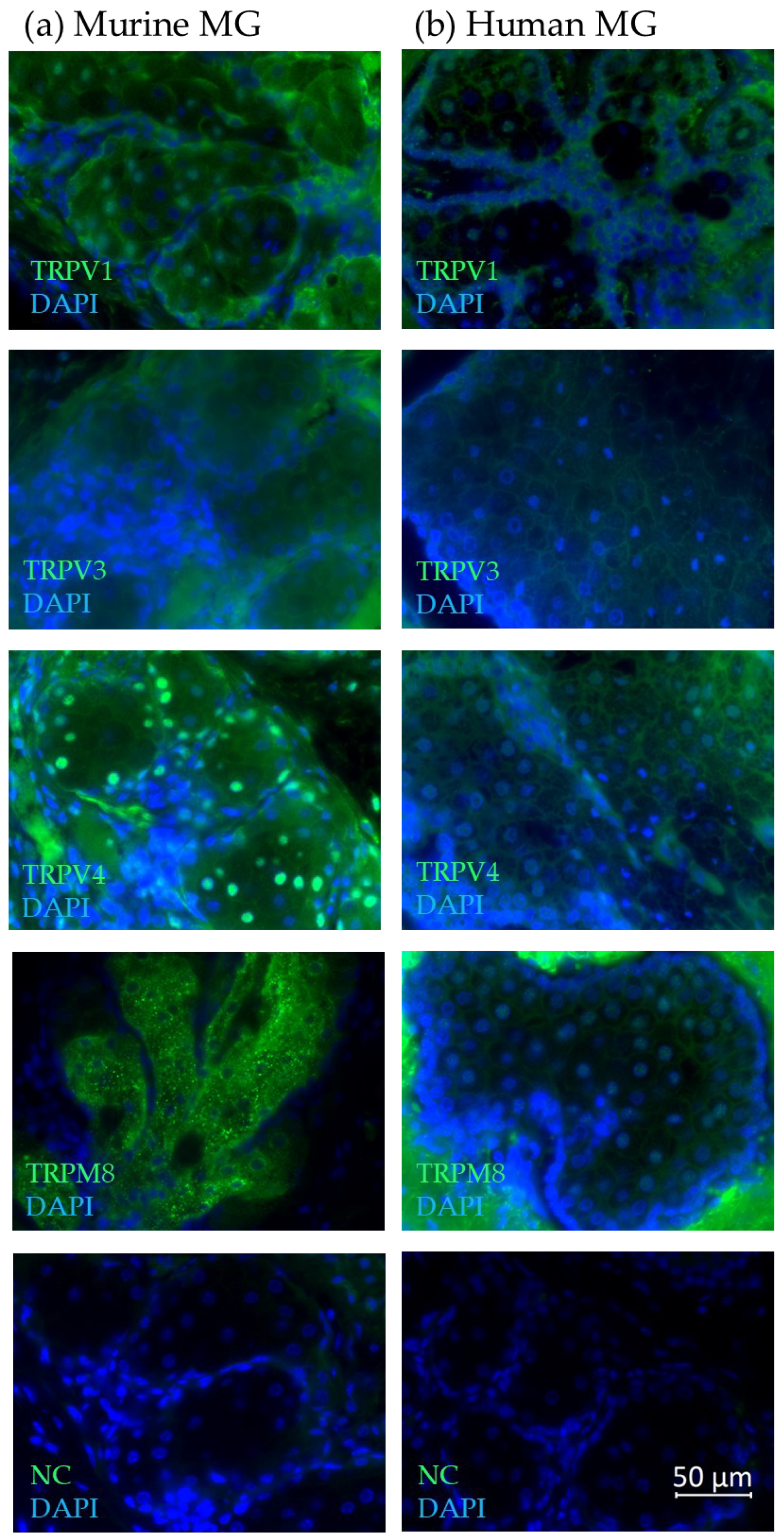
2.1. Temperature-Sensitive TRP Channel Expression in Meibomian Glands
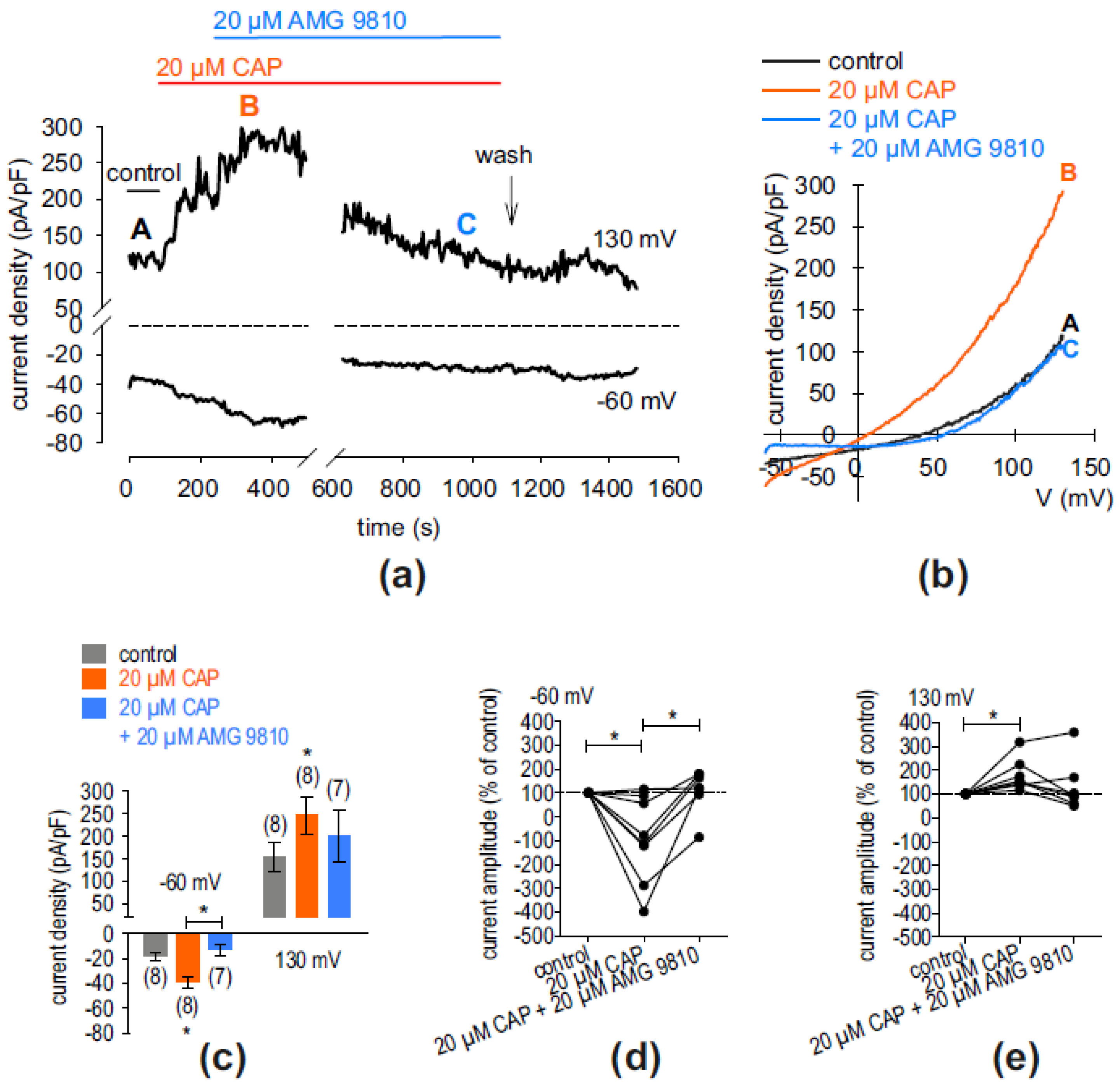
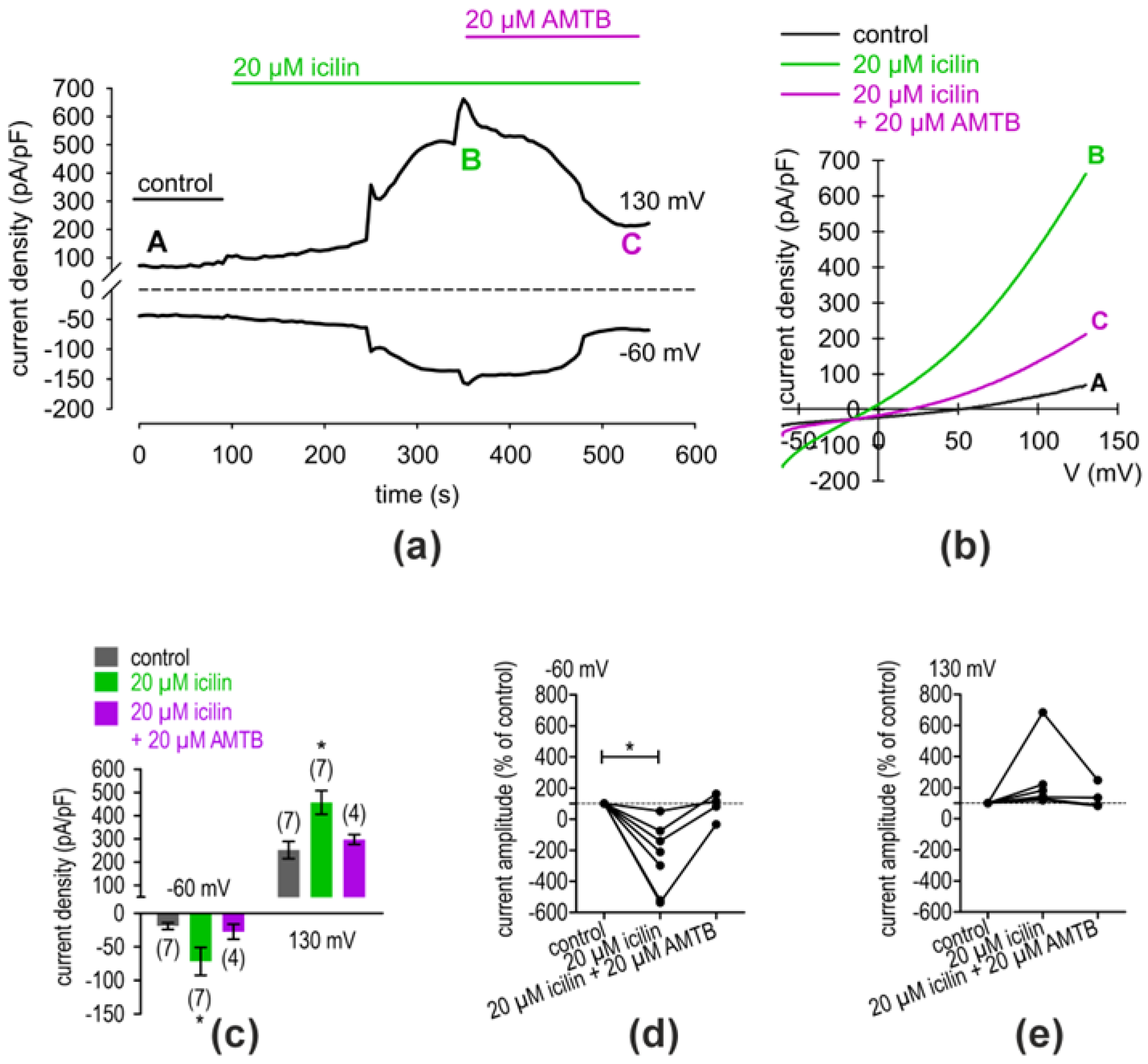
2.2. Whole-Cell Current Increase via TRPV1 and TRPM8 in hMGECs
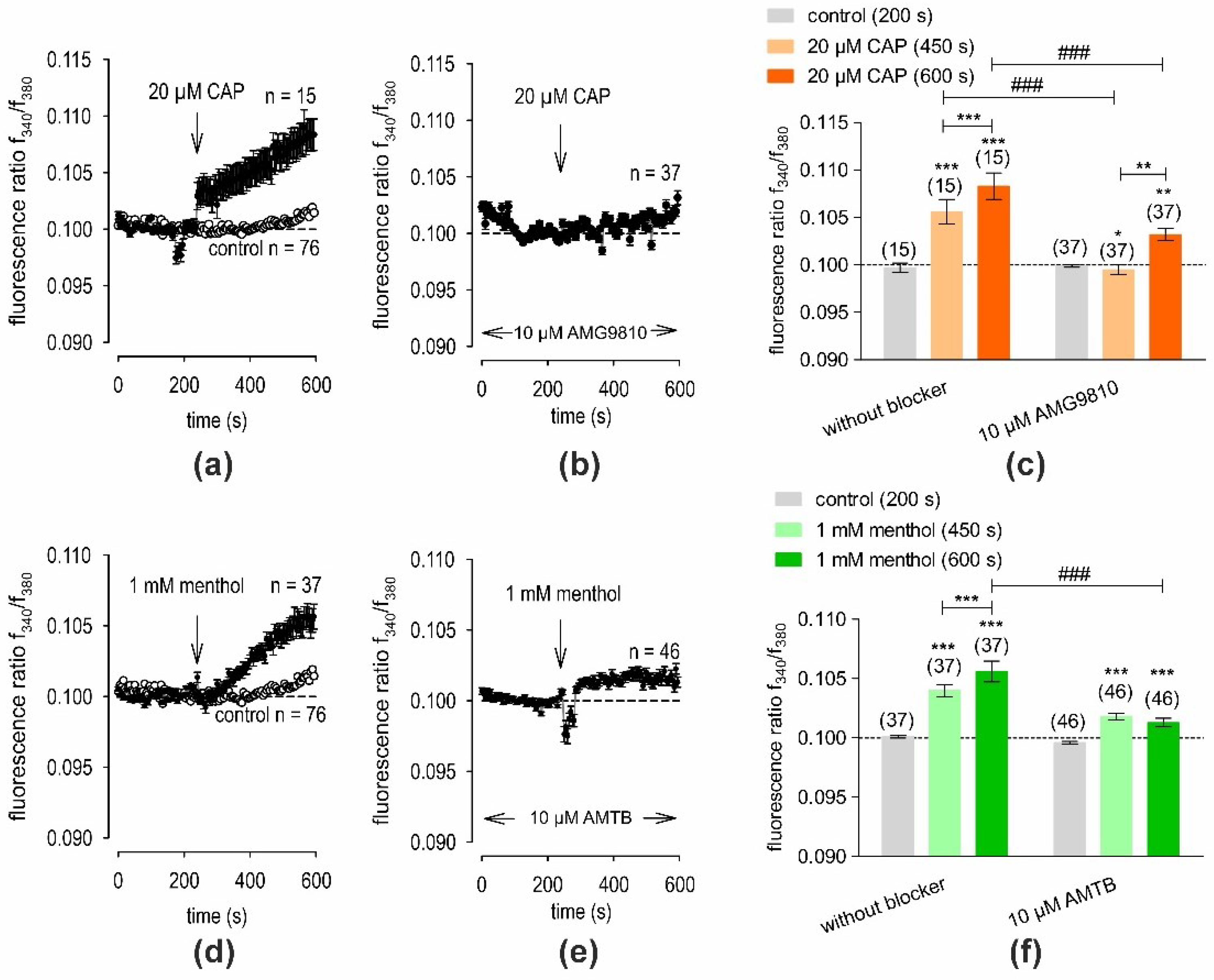
2.3. Calcium Regulation via TRPV1 and TRPM8 in hMGECs
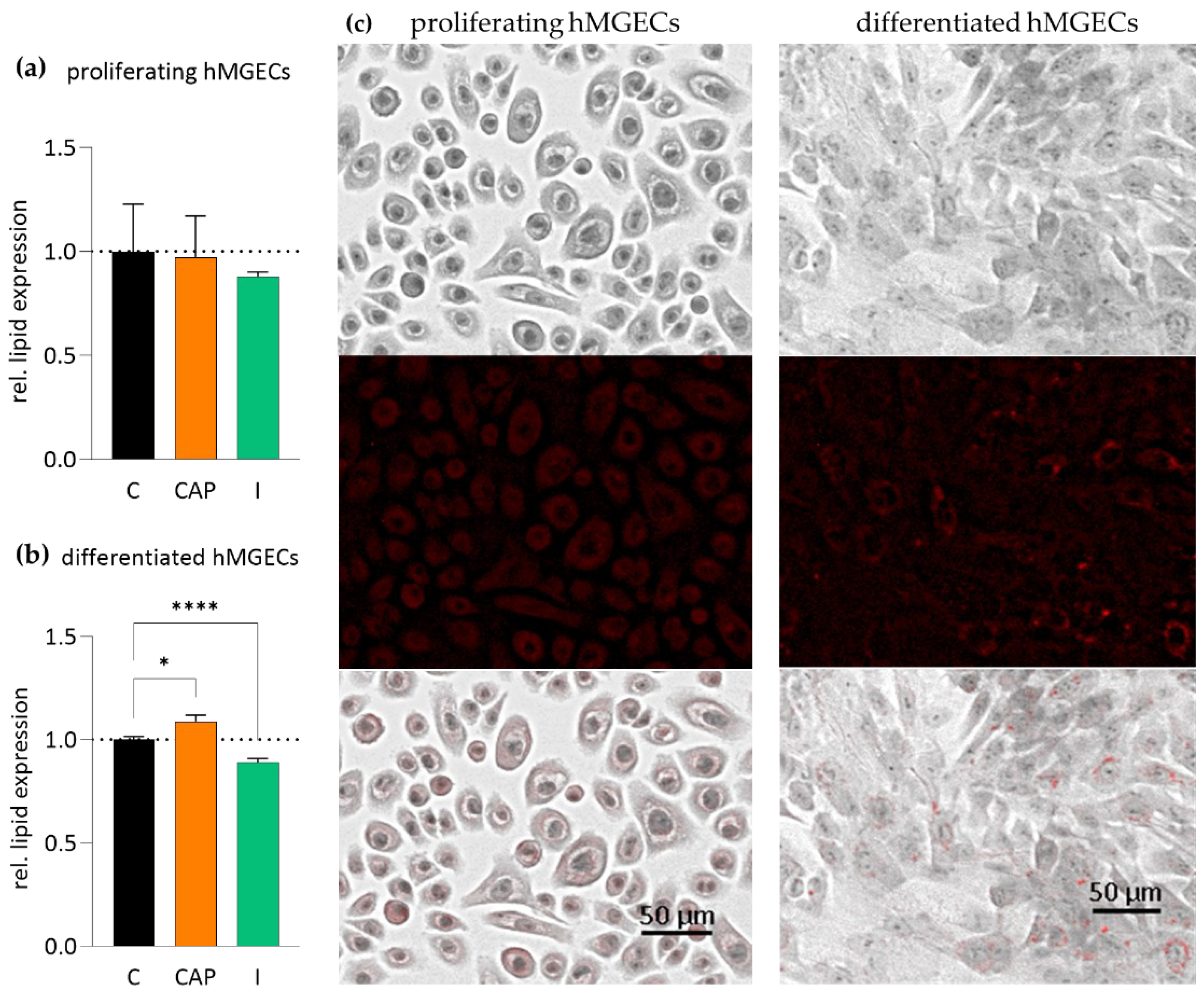
2.4. Influence of TRPV1 and TRPM8 Activation on hMGEC Lipogenesis
3. Discussion
3.1. Temperature-Sensitive TRPs Are Expressed and Functionally Active in the Meibomian Glands
3.2. TRPV1 and TRPM8 Activation Influence the Lipid Synthesis of Meibocytes
4. Materials and Methods
4.1. Cells
4.2. Tissues
4.3. RNA Isolation and cDNA Synthesis
4.4. Reverse Transcriptase Polymerase Chain Reaction (RT-PCR)
4.5. Quantitative Real-Time Polymerase Chain Reaction (qPCR)
4.6. Protein Isolation and Western Blot
4.7. Immunofluorescence
4.8. Planar Patch-Clamp Recordings
4.9. Calcium Fluorescence Imaging
4.10. Viability Assay
4.11. Oil Red O Assays
4.12. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zemanova, M. Dry Eye Disease. A Review. Cesk Slov. Oftalmol. 2021, 77, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Rouen, P.A.; White, M.L. Dry Eye Disease: Prevalence, Assessment, and Management. Home Healthc. Now. 2018, 36, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Schaumberg, D.A.; Sullivan, D.A.; Buring, J.E.; Dana, M.R. Prevalence of dry eye syndrome among US women. Am. J. Ophthalmol. 2003, 136, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Shanti, Y.; Shehada, R.; Bakkar, M.M.; Qaddumi, J. Prevalence and associated risk factors of dry eye disease in 16 northern West bank towns in Palestine: A cross-sectional study. BMC Ophthalmol. 2020, 20, 26. [Google Scholar] [CrossRef] [PubMed]
- Craig, J.P.; Nichols, K.K.; Akpek, E.K.; Caffery, B.; Dua, H.S.; Joo, C.K.; Liu, Z.; Nelson, J.D.; Nichols, J.J.; Tsubota, K.; et al. TFOS DEWS II Definition and Classification Report. Ocul. Surf. 2017, 15, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, J.; Garreis, F.; Paulsen, F. Pathophysiology of Meibomian Glands—An Overview. Ocul. Immunol. Inflamm. 2021, 29, 803–810. [Google Scholar] [CrossRef]
- Willcox, M.D.P.; Argueso, P.; Georgiev, G.A.; Holopainen, J.M.; Laurie, G.W.; Millar, T.J.; Papas, E.B.; Rolland, J.P.; Schmidt, T.A.; Stahl, U.; et al. TFOS DEWS II Tear Film Report. Ocul. Surf. 2017, 15, 366–403. [Google Scholar] [CrossRef]
- Jester, J.V.; Nicolaides, N.; Smith, R.E. Meibomian gland studies: Histologic and ultrastructural investigations. Investig. Ophthalmol. Vis. Sci. 1981, 20, 537–547. [Google Scholar]
- Suzuki, T.; Sutani, T.; Nakai, H.; Shirahige, K.; Kinoshita, S. The Microbiome of the Meibum and Ocular Surface in Healthy Subjects. Investig. Ophthalmol. Vis. Sci. 2020, 61, 18. [Google Scholar] [CrossRef]
- Olami, Y.; Zajicek, G.; Cogan, M.; Gnessin, H.; Pe’er, J. Turnover and migration of meibomian gland cells in rats’ eyelids. Ophthalmic Res. 2001, 33, 170–175. [Google Scholar] [CrossRef]
- Bron, A.J.; de Paiva, C.S.; Chauhan, S.K.; Bonini, S.; Gabison, E.E.; Jain, S.; Knop, E.; Markoulli, M.; Ogawa, Y.; Perez, V.; et al. TFOS DEWS II pathophysiology report. Ocul. Surf. 2017, 15, 438–510. [Google Scholar] [CrossRef] [PubMed]
- Knop, E.; Knop, N.; Millar, T.; Obata, H.; Sullivan, D.A. The international workshop on meibomian gland dysfunction: Report of the subcommittee on anatomy, physiology, and pathophysiology of the meibomian gland. Investig. Ophthalmol. Vis. Sci. 2011, 52, 1938–1978. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, D.A.; Sullivan, B.D.; Evans, J.E.; Schirra, F.; Yamagami, H.; Liu, M.; Richards, S.M.; Suzuki, T.; Schaumberg, D.A.; Sullivan, R.M.; et al. Androgen deficiency, Meibomian gland dysfunction, and evaporative dry eye. Ann. N. Y. Acad. Sci. 2002, 966, 211–222. [Google Scholar] [CrossRef]
- Yang, J.M.; Wei, E.T.; Kim, S.J.; Yoon, K.C. TRPM8 Channels and Dry Eye. Pharmaceuticals 2018, 11, 125. [Google Scholar] [CrossRef] [PubMed]
- Reinach, P.S.; Mergler, S.; Okada, Y.; Saika, S. Ocular transient receptor potential channel function in health and disease. BMC Ophthalmol. 2015, 15 (Suppl S1), 153. [Google Scholar] [CrossRef]
- Kaneko, Y.; Szallasi, A. Transient receptor potential (TRP) channels: A clinical perspective. Br. J. Pharmacol. 2014, 171, 2474–2507. [Google Scholar] [CrossRef] [PubMed]
- Nilius, B.; Owsianik, G. Transient receptor potential channelopathies. Pflugers Arch. 2010, 460, 437–450. [Google Scholar] [CrossRef]
- Fakih, D.; Migeon, T.; Moreau, N.; Baudouin, C.; Reaux-Le Goazigo, A.; Melik Parsadaniantz, S. Transient Receptor Potential Channels: Important Players in Ocular Pain and Dry Eye Disease. Pharmaceutics 2022, 14, 1859. [Google Scholar] [CrossRef]
- Bereiter, D.A.; Rahman, M.; Thompson, R.; Stephenson, P.; Saito, H. TRPV1 and TRPM8 Channels and Nocifensive Behavior in a Rat Model for Dry Eye. Investig. Ophthalmol. Vis. Sci. 2018, 59, 3739–3746. [Google Scholar] [CrossRef]
- Hilton, J.K.; Rath, P.; Helsell, C.V.; Beckstein, O.; Van Horn, W.D. Understanding thermosensitive transient receptor potential channels as versatile polymodal cellular sensors. Biochemistry 2015, 54, 2401–2413. [Google Scholar] [CrossRef]
- Mergler, S.; Garreis, F.; Sahlmuller, M.; Reinach, P.S.; Paulsen, F.; Pleyer, U. Thermosensitive transient receptor potential channels in human corneal epithelial cells. J. Cell Physiol. 2011, 226, 1828–1842. [Google Scholar] [CrossRef] [PubMed]
- Khajavi, N.; Reinach, P.S.; Slavi, N.; Skrzypski, M.; Lucius, A.; Strauss, O.; Kohrle, J.; Mergler, S. Thyronamine induces TRPM8 channel activation in human conjunctival epithelial cells. Cell Signal 2015, 27, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Yang, H.; Wang, Z.; Mergler, S.; Liu, H.; Kawakita, T.; Tachado, S.D.; Pan, Z.; Capo-Aponte, J.E.; Pleyer, U.; et al. Transient receptor potential vanilloid 1 activation induces inflammatory cytokine release in corneal epithelium through MAPK signaling. J. Cell Physiol. 2007, 213, 730–739. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.Y.; Liu, B.C.; Wang, L.H.; Yang, L.L.; Bao, Q.; Zhai, Y.J.; Alli, A.A.; Thai, T.L.; Eaton, D.C.; Wang, W.Z.; et al. Acute ethanol induces apoptosis by stimulating TRPC6 via elevation of superoxide in oxygenated podocytes. Biochim. Biophys. Acta 2015, 1853, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Skrzypski, M.; Khajavi, N.; Mergler, S.; Szczepankiewicz, D.; Kolodziejski, P.A.; Metzke, D.; Wojciechowicz, T.; Billert, M.; Nowak, K.W.; Strowski, M.Z. TRPV6 channel modulates proliferation of insulin secreting INS-1E beta cell line. Biochim. Biophys. Acta 2015, 1853, 3202–3210. [Google Scholar] [CrossRef] [PubMed]
- Toth, B.I.; Benko, S.; Szollosi, A.G.; Kovacs, L.; Rajnavolgyi, E.; Biro, T. Transient receptor potential vanilloid-1 signaling inhibits differentiation and activation of human dendritic cells. FEBS Lett. 2009, 583, 1619–1624. [Google Scholar] [CrossRef] [PubMed]
- Ou-Yang, Q.; Li, B.; Xu, M.; Liang, H. TRPV4 promotes the migration and invasion of glioma cells via AKT/Rac1 signaling. Biochem. Biophys. Res. Commun. 2018, 503, 876–881. [Google Scholar] [CrossRef] [PubMed]
- Su, K.H.; Lin, S.J.; Wei, J.; Lee, K.I.; Zhao, J.F.; Shyue, S.K.; Lee, T.S. The essential role of transient receptor potential vanilloid 1 in simvastatin-induced activation of endothelial nitric oxide synthase and angiogenesis. Acta Physiol 2014, 212, 191–204. [Google Scholar] [CrossRef] [PubMed]
- Smani, T.; Gomez, L.J.; Regodon, S.; Woodard, G.E.; Siegfried, G.; Khatib, A.M.; Rosado, J.A. TRP Channels in Angiogenesis and Other Endothelial Functions. Front. Physiol. 2018, 9, 1731. [Google Scholar] [CrossRef]
- Caterina, M.J. Transient receptor potential ion channels as participants in thermosensation and thermoregulation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R64–R76. [Google Scholar] [CrossRef]
- Tominaga, M.; Caterina, M.J. Thermosensation and pain. J. Neurobiol. 2004, 61, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Vay, L.; Gu, C.; McNaughton, P.A. The thermo-TRP ion channel family: Properties and therapeutic implications. Br. J. Pharmacol. 2012, 165, 787–801. [Google Scholar] [CrossRef] [PubMed]
- Parra, A.; Madrid, R.; Echevarria, D.; del Olmo, S.; Morenilla-Palao, C.; Acosta, M.C.; Gallar, J.; Dhaka, A.; Viana, F.; Belmonte, C. Ocular surface wetness is regulated by TRPM8-dependent cold thermoreceptors of the cornea. Nat. Med. 2010, 16, 1396–1399. [Google Scholar] [CrossRef]
- Reinach, P.S.; Chen, W.; Mergler, S. Polymodal roles of transient receptor potential channels in the control of ocular function. Eye Vis. 2015, 2, 5. [Google Scholar] [CrossRef] [PubMed]
- Mergler, S.; Cheng, Y.; Skosyrski, S.; Garreis, F.; Pietrzak, P.; Kociok, N.; Dwarakanath, A.; Reinach, P.S.; Kakkassery, V. Altered calcium regulation by thermosensitive transient receptor potential channels in etoposide-resistant WERI-Rb1 retinoblastoma cells. Exp. Eye Res. 2012, 94, 157–173. [Google Scholar] [CrossRef] [PubMed]
- Mergler, S.; Derckx, R.; Reinach, P.S.; Garreis, F.; Bohm, A.; Schmelzer, L.; Skosyrski, S.; Ramesh, N.; Abdelmessih, S.; Polat, O.K.; et al. Calcium regulation by temperature-sensitive transient receptor potential channels in human uveal melanoma cells. Cell. Signal. 2014, 26, 56–69. [Google Scholar] [CrossRef]
- Mergler, S.; Garreis, F.; Sahlmuller, M.; Lyras, E.M.; Reinach, P.S.; Dwarakanath, A.; Paulsen, F.; Pleyer, U. Calcium regulation by thermo- and osmosensing transient receptor potential vanilloid channels (TRPVs) in human conjunctival epithelial cells. Histochem. Cell Biol. 2012, 137, 743–761. [Google Scholar] [CrossRef] [PubMed]
- Garreis, F.; Schroder, A.; Reinach, P.S.; Zoll, S.; Khajavi, N.; Dhandapani, P.; Lucius, A.; Pleyer, U.; Paulsen, F.; Mergler, S. Upregulation of Transient Receptor Potential Vanilloid Type-1 Channel Activity and Ca2+ Influx Dysfunction in Human Pterygial Cells. Invest. Ophthalmol. Vis. Sci. 2016, 57, 2564–2577. [Google Scholar] [CrossRef]
- Talavera, K.; Startek, J.B.; Alvarez-Collazo, J.; Boonen, B.; Alpizar, Y.A.; Sanchez, A.; Naert, R.; Nilius, B. Mammalian Transient Receptor Potential TRPA1 Channels: From Structure to Disease. Physiol. Rev. 2020, 100, 725–803. [Google Scholar] [CrossRef]
- Katagiri, A.; Thompson, R.; Rahman, M.; Okamoto, K.; Bereiter, D.A. Evidence for TRPA1 involvement in central neural mechanisms in a rat model of dry eye. Neuroscience 2015, 290, 204–213. [Google Scholar] [CrossRef]
- Okada, Y.; Reinach, P.S.; Shirai, K.; Kitano-Izutani, A.; Miyajima, M.; Yamanaka, O.; Sumioka, T.; Saika, S. Transient Receptor Potential Channels and Corneal Stromal Inflammation. Cornea 2015, 34 (Suppl. 11), S136–S141. [Google Scholar] [CrossRef] [PubMed]
- Souza Monteiro de Araujo, D.; Nassini, R.; Geppetti, P.; De Logu, F. TRPA1 as a therapeutic target for nociceptive pain. Expert. Opin. Ther. Targets 2020, 24, 997–1008. [Google Scholar] [CrossRef] [PubMed]
- Vriens, J.; Appendino, G.; Nilius, B. Pharmacology of vanilloid transient receptor potential cation channels. Mol. Pharmacol. 2009, 75, 1262–1279. [Google Scholar] [CrossRef] [PubMed]
- Gavva, N.R.; Tamir, R.; Qu, Y.; Klionsky, L.; Zhang, T.J.; Immke, D.; Wang, J.; Zhu, D.; Vanderah, T.W.; Porreca, F.; et al. AMG 9810 [(E)-3-(4-t-butylphenyl)-N-(2,3-dihydrobenzo[b][1,4] dioxin-6-yl)acrylamide], a novel vanilloid receptor 1 (TRPV1) antagonist with antihyperalgesic properties. J. Pharmacol. Exp. Ther. 2005, 313, 474–484. [Google Scholar] [CrossRef] [PubMed]
- McKemy, D.D.; Neuhausser, W.M.; Julius, D. Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature 2002, 416, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Lashinger, E.S.; Steiginga, M.S.; Hieble, J.P.; Leon, L.A.; Gardner, S.D.; Nagilla, R.; Davenport, E.A.; Hoffman, B.E.; Laping, N.J.; Su, X. AMTB, a TRPM8 channel blocker: Evidence in rats for activity in overactive bladder and painful bladder syndrome. Am. J. Physiol. Renal Physiol. 2008, 295, F803–F810. [Google Scholar] [CrossRef] [PubMed]
- Chuang, H.H.; Neuhausser, W.M.; Julius, D. The super-cooling agent icilin reveals a mechanism of coincidence detection by a temperature-sensitive TRP channel. Neuron 2004, 43, 859–869. [Google Scholar] [CrossRef] [PubMed]
- Lucius, A.; Chhatwal, S.; Valtink, M.; Reinach, P.S.; Li, A.; Pleyer, U.; Mergler, S. L-Carnitine Suppresses Transient Receptor Potential Vanilloid Type 1 Activation in Human Corneal Epithelial Cells. Int. J. Mol. Sci. 2023, 24, 11815. [Google Scholar] [CrossRef]
- Rawls, S.M.; Gomez, T.; Ding, Z.; Raffa, R.B. Differential behavioral effect of the TRPM8/TRPA1 channel agonist icilin (AG-3-5). Eur. J. Pharmacol. 2007, 575, 103–104. [Google Scholar] [CrossRef]
- Taberner, F.J.; Fernandez-Ballester, G.; Fernandez-Carvajal, A.; Ferrer-Montiel, A. TRP channels interaction with lipids and its implications in disease. Biochim. Biophys. Acta 2015, 1848, 1818–1827. [Google Scholar] [CrossRef]
- Ciardo, M.G.; Ferrer-Montiel, A. Lipids as central modulators of sensory TRP channels. Biochim. Biophys. Acta Biomembr. 2017, 1859, 1615–1628. [Google Scholar] [CrossRef] [PubMed]
- Seki, N.; Shirasaki, H.; Kikuchi, M.; Sakamoto, T.; Watanabe, N.; Himi, T. Expression and localization of TRPV1 in human nasal mucosa. Rhinology 2006, 44, 128–134. [Google Scholar] [PubMed]
- Toth, B.I.; Geczy, T.; Griger, Z.; Dozsa, A.; Seltmann, H.; Kovacs, L.; Nagy, L.; Zouboulis, C.C.; Paus, R.; Biro, T. Transient receptor potential vanilloid-1 signaling as a regulator of human sebocyte biology. J. Investig. Dermatol. 2009, 129, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, Z.; Leger, S.; Bitton, E. Ex vivo heat retention of different eyelid warming masks. Contact Lens Anterior Eye 2015, 38, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Rantamaki, A.H.; Wiedmer, S.K.; Holopainen, J.M. Melting points--the key to the anti-evaporative effect of the tear film wax esters. Investig. Ophthalmol. Vis. Sci. 2013, 54, 5211–5217. [Google Scholar] [CrossRef] [PubMed]
- Belmonte, C.; Nichols, J.J.; Cox, S.M.; Brock, J.A.; Begley, C.G.; Bereiter, D.A.; Dartt, D.A.; Galor, A.; Hamrah, P.; Ivanusic, J.J.; et al. TFOS DEWS II pain and sensation report. Ocul. Surf. 2017, 15, 404–437. [Google Scholar] [CrossRef] [PubMed]
- Koivisto, A.P.; Belvisi, M.G.; Gaudet, R.; Szallasi, A. Advances in TRP channel drug discovery: From target validation to clinical studies. Nat. Rev. Drug Discov. 2022, 21, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, C.; Martin-Martinez, M.; Gomez-Monterrey, I.; Gonzalez-Muniz, R. TRPM8 Channels: Advances in Structural Studies and Pharmacological Modulation. Int. J. Mol. Sci. 2021, 22, 8502. [Google Scholar] [CrossRef] [PubMed]
- Hirata, H.; Oshinsky, M.L. Ocular dryness excites two classes of corneal afferent neurons implicated in basal tearing in rats: Involvement of transient receptor potential channels. J. Neurophysiol. 2012, 107, 1199–1209. [Google Scholar] [CrossRef]
- Yang, J.M.; Li, F.; Liu, Q.; Ruedi, M.; Wei, E.T.; Lentsman, M.; Lee, H.S.; Choi, W.; Kim, S.J.; Yoon, K.C. A novel TRPM8 agonist relieves dry eye discomfort. BMC Ophthalmol. 2017, 17, 101. [Google Scholar] [CrossRef]
- Yoon, H.J.; Kim, J.; Yang, J.M.; Wei, E.T.; Kim, S.J.; Yoon, K.C. Topical TRPM8 Agonist for Relieving Neuropathic Ocular Pain in Patients with Dry Eye: A Pilot Study. J. Clin. Med. 2021, 10, 250. [Google Scholar] [CrossRef] [PubMed]
- Wirta, D.L.; Senchyna, M.; Lewis, A.E.; Evans, D.G.; McLaurin, E.B.; Ousler, G.W.; Hollander, D.A. A randomized, vehicle-controlled, Phase 2b study of two concentrations of the TRPM8 receptor agonist AR-15512 in the treatment of dry eye disease (COMET-1). Ocul. Surf. 2022, 26, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Benitez-Del-Castillo, J.M.; Moreno-Montanes, J.; Jimenez-Alfaro, I.; Munoz-Negrete, F.J.; Turman, K.; Palumaa, K.; Sadaba, B.; Gonzalez, M.V.; Ruz, V.; Vargas, B.; et al. Safety and Efficacy Clinical Trials for SYL1001, a Novel Short Interfering RNA for the Treatment of Dry Eye Disease. Investig. Ophthalmol. Vis. Sci. 2016, 57, 6447–6454. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Montanes, J.; Bleau, A.M.; Jimenez, A.I. Tivanisiran, a novel siRNA for the treatment of dry eye disease. Expert. Opin. Investig. Drugs 2018, 27, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Seki, N.; Shirasaki, H.; Kikuchi, M.; Himi, T. Capsaicin induces the production of IL-6 in human upper respiratory epithelial cells. Life Sci. 2007, 80, 1592–1597. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, D.A.; Liu, Y.; Kam, W.R.; Ding, J.; Green, K.M.; Shaffer, S.A.; Hatton, M.P.; Liu, S. Serum-induced differentiation of human meibomian gland epithelial cells. Investig. Ophthalmol. Vis. Sci. 2014, 55, 3866–3877. [Google Scholar] [CrossRef]
- Hampel, U.; Garreis, F. The human meibomian gland epithelial cell line as a model to study meibomian gland dysfunction. Exp. Eye Res. 2017, 163, 46–52. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Feldt, J.; Schicht, M.; Welss, J.; Gelse, K.; Sesselmann, S.; Tsokos, M.; Socher, E.; Garreis, F.; Muller, T.; Paulsen, F. Production and Secretion of Gelsolin by Both Human Macrophage- and Fibroblast-like Synoviocytes and GSN Modulation in the Synovial Fluid of Patients with Various Forms of Arthritis. Biomedicines 2022, 10, 723. [Google Scholar] [CrossRef]
- Garreis, F.; Schlorf, T.; Worlitzsch, D.; Steven, P.; Brauer, L.; Jager, K.; Paulsen, F.P. Roles of human beta-defensins in innate immune defense at the ocular surface: Arming and alarming corneal and conjunctival epithelial cells. Histochem. Cell Biol. 2010, 134, 59–73. [Google Scholar] [CrossRef]
- Barry, P.H. JPCalc, a software package for calculating liquid junction potential corrections in patch-clamp, intracellular, epithelial and bilayer measurements and for correcting junction potential measurements. J. Neurosci. Methods 1994, 51, 107–116. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| n = 12 | Lipid Content Compared to a Control | p-Value |
|---|---|---|
| Prol. hMGECs 10 µM CAP | −2.8 ± 5.74% | >0.05 |
| Prol. hMGECs 100 µM I | −12.1 ± 0.65% | >0.05 |
| Diff. hMGECs 10 µM CAP | +8.8 ± 3.02% | <0.05 |
| Diff. hMGECs 100 µM I | −10.97 ± 1.73% | <0.0001 |
| RT-PCR | Forward | Reverse | Product Length |
|---|---|---|---|
| Primer | |||
| Human | |||
| TRPV1 | CTCCTACAACAGCCTGTAC | AAGGCCTTCCTCATGCACT | 285 bp |
| TRPV2 | CTCTGGTGGCTAGCCTGTCCTGACA | TGGGATCCCGGAGCTTCTCA | 228 bp |
| TRPV3 | GCTGAAGAAGCGCATCTTTGCA | TCATAGGCCTCCTCTGTGTACT | 288 bp |
| TRPV4 | TACCTGTGTGCCATGGTCATCT | TGCTATAGGTCCCCGTCAGCTT | 379 bp |
| TRPM8 | CCTGTTCCTCTTTGCGGTGTGGAT | TCCTCTGAGGTGTCGTTGGCTTT | 621 bp |
| TRPA1 | GACCACAATGGCTGGACAGCT | GTACCATTGCGTTGAGGGCTGT | 541 bp |
| murine | |||
| TRPV1 | CCCCCAAAACAGTAGCTTCA | AGCAACACCAGCCCAATTAC | 239 bp |
| TRPV2 | TGATGAAGGCTGTGCTGAAC | CACCACAGGCTCCTCTTCTC | 174 bp |
| TRPV3 | AGGCTTCTATTTTGGCGAGACAC | TCCCGAGGACGGTAGTAAGAGAC | 623 bp |
| TRPV4 | ACAACACCCGAGAGAACACC | TGAACTTGCGAGACAGATGC | 221 bp |
| TRPM8 | CGACAACTCAGAGGAGATGAGG | GGGATGGGGTAGGACTCTTTG | 525 bp |
| TRPA1 | GGAGCAGACATCAACAGCAC | GCAGGGGCGACTTCTTATC | 393 bp |
| qPCR | |||
| human | |||
| TRPV1 | CAGCAGCGAGACCCCTAA | CCTGCAGGAGTCGGTTCA | 65 bp |
| TRPV4 | CAACAACGACGGCCTCTC | GGATGATGTGCTGAAAGATCC | 73 bp |
| TRPM8 | GCCAAAGTGAAGAACGACATC | ACACTCATGAACAGCTCAACAG | 93 bp |
| Antibodies | Manufacturer | Product Number | IF | WB |
|---|---|---|---|---|
| primary | ||||
| TRPV1 | alomonelabs | ACC-030 | 1:50 | 1:200 |
| TRPV3 | antikoerper-online.de | ABIN863127 | 1:100 | 1:1000 |
| TRPV4 | alomone labs | ACC-034 | 1:150 | 1:200 |
| TRPM8 | BIOZOL | LS-B6607 | 1:50 | 1:1000 |
| GAPDH | Santa Cruz biotechnology | sc-365062 | 1:2000 | |
| secondary | ||||
| HRP | Agilent Dako | P0448 | 1:5000 | |
| Alexa Fluor 488 | Thermo Fisher Scientific | A11008 | 1:1000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keller, M.; Mergler, S.; Li, A.; Zahn, I.; Paulsen, F.; Garreis, F. Thermosensitive TRP Channels Are Functionally Expressed and Influence the Lipogenesis in Human Meibomian Gland Cells. Int. J. Mol. Sci. 2024, 25, 4043. https://doi.org/10.3390/ijms25074043
Keller M, Mergler S, Li A, Zahn I, Paulsen F, Garreis F. Thermosensitive TRP Channels Are Functionally Expressed and Influence the Lipogenesis in Human Meibomian Gland Cells. International Journal of Molecular Sciences. 2024; 25(7):4043. https://doi.org/10.3390/ijms25074043
Chicago/Turabian StyleKeller, Melina, Stefan Mergler, Aruna Li, Ingrid Zahn, Friedrich Paulsen, and Fabian Garreis. 2024. "Thermosensitive TRP Channels Are Functionally Expressed and Influence the Lipogenesis in Human Meibomian Gland Cells" International Journal of Molecular Sciences 25, no. 7: 4043. https://doi.org/10.3390/ijms25074043
APA StyleKeller, M., Mergler, S., Li, A., Zahn, I., Paulsen, F., & Garreis, F. (2024). Thermosensitive TRP Channels Are Functionally Expressed and Influence the Lipogenesis in Human Meibomian Gland Cells. International Journal of Molecular Sciences, 25(7), 4043. https://doi.org/10.3390/ijms25074043