
Antimicrobial Activities of α-Helix and β-Sheet Peptides against the Major Bovine Respiratory Disease Agent, Mannheimia haemolytica




Abstract
:1. Introduction
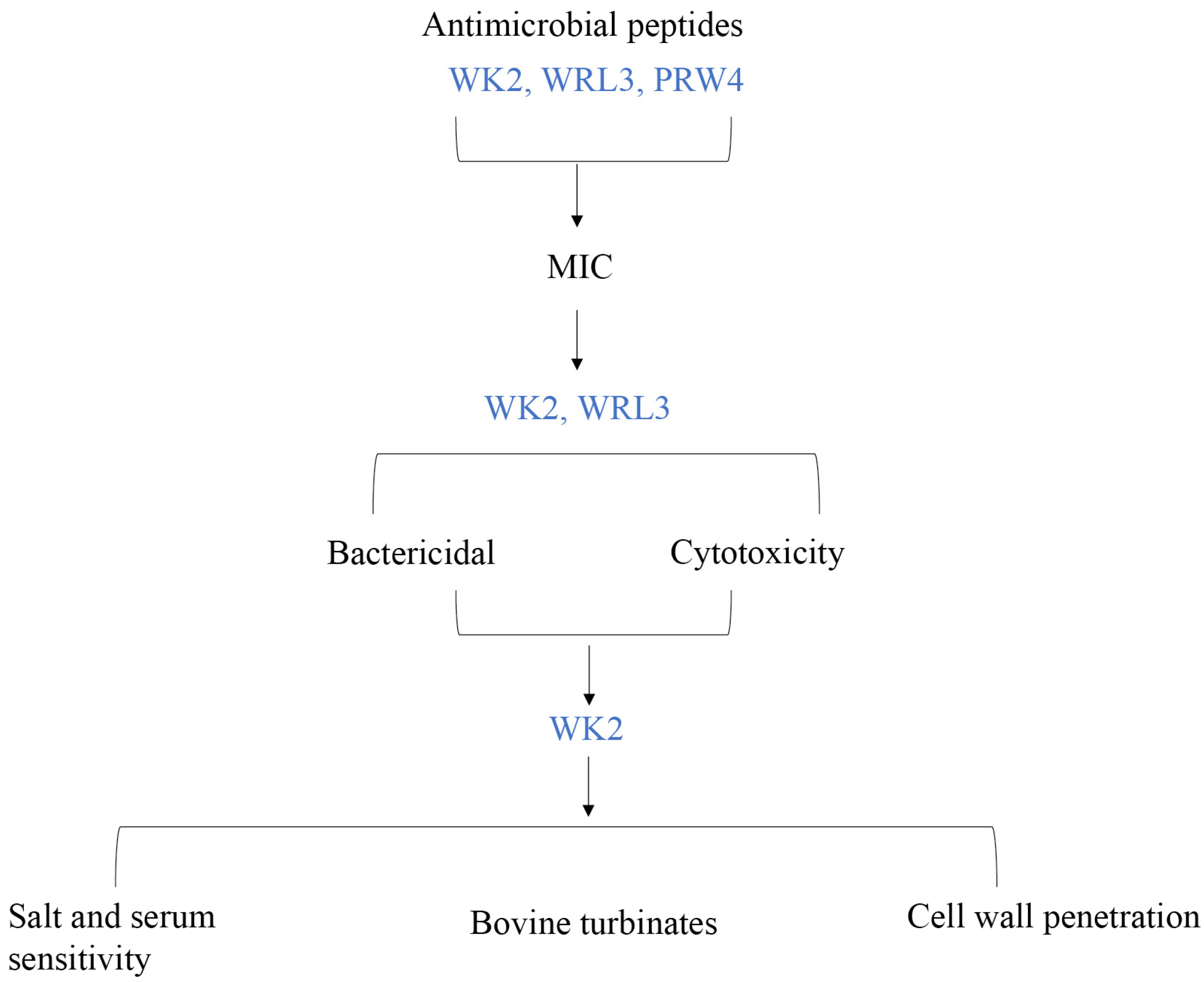
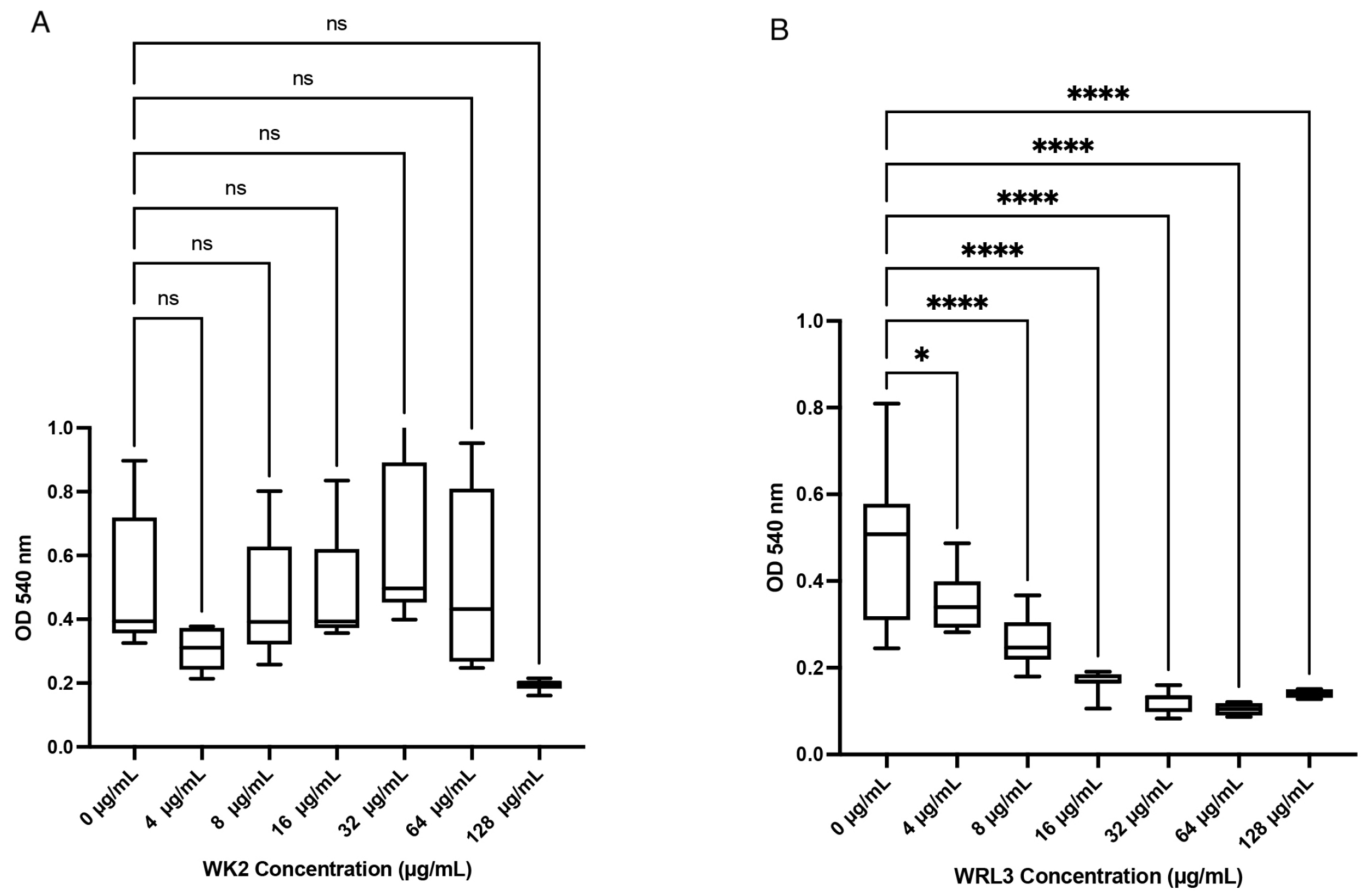
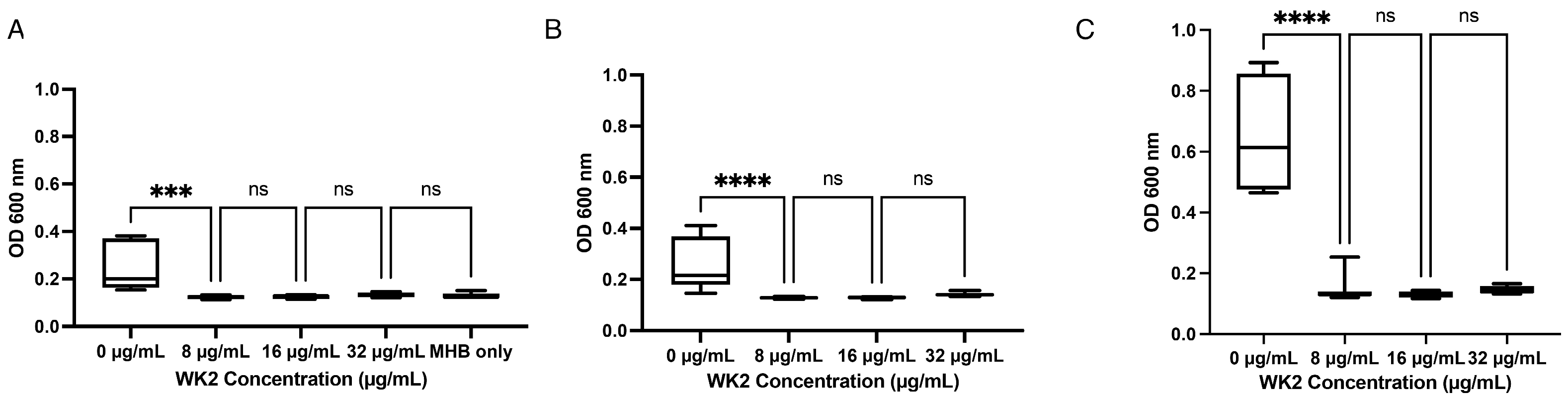
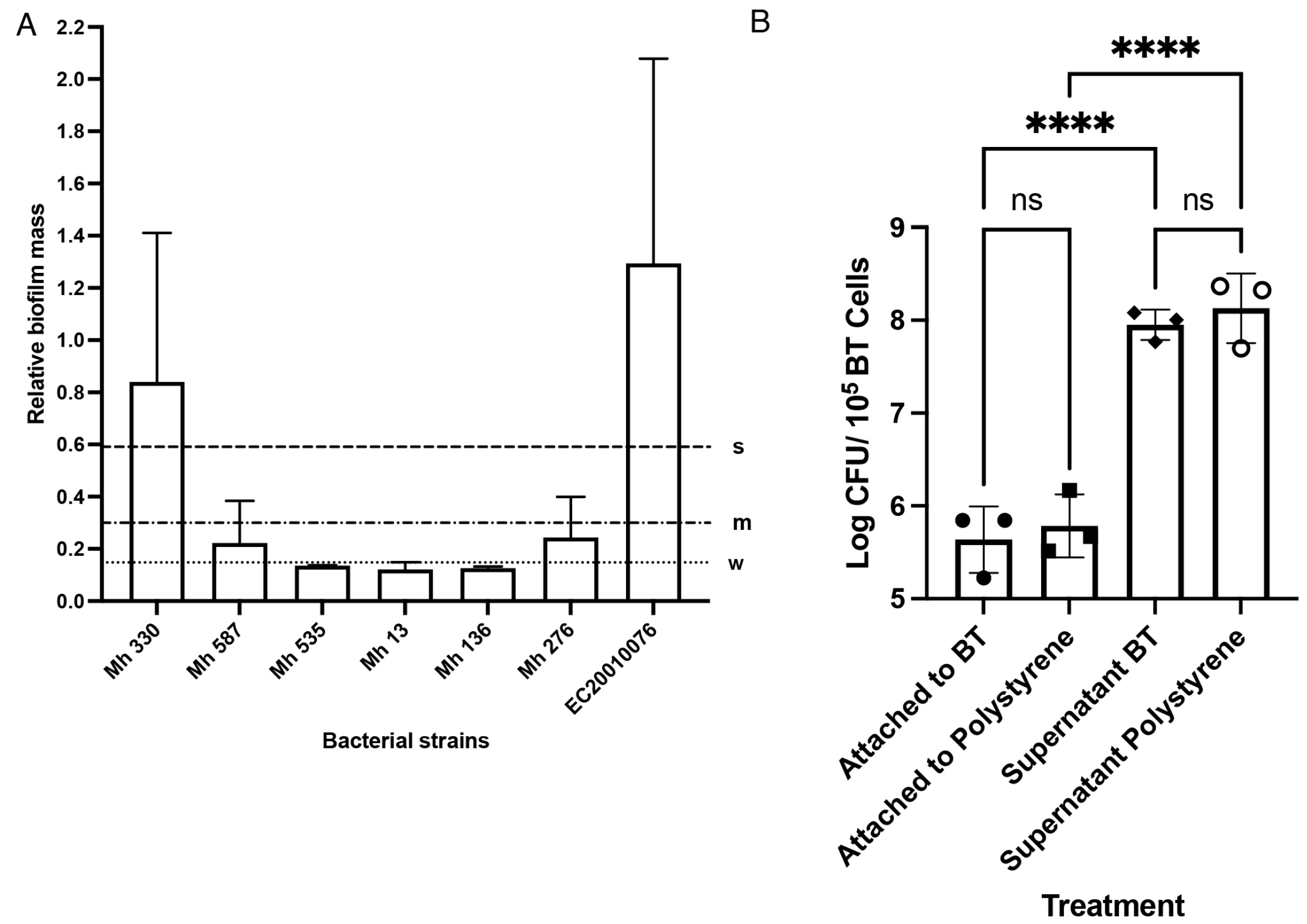
2. Results
3. Discussion
4. Materials and Methods
4.1. Synthesis of the Peptides
4.2. Bacterial Strains and Cell Culture
4.3. In Vitro Minimum Inhibitory Concentration (MIC) Assays of M. haemolytica
4.4. Assess Effect of Salt and Serum on Antibacterial Activity of Antimicrobial Peptides
4.5. Assess Cytotoxic Effects Using the MTT (3-(4,5-dimethylthiozol-2-yl)-2,5-diphenyltetrazolium Bromide) Assay
4.6. Biofilm Screening
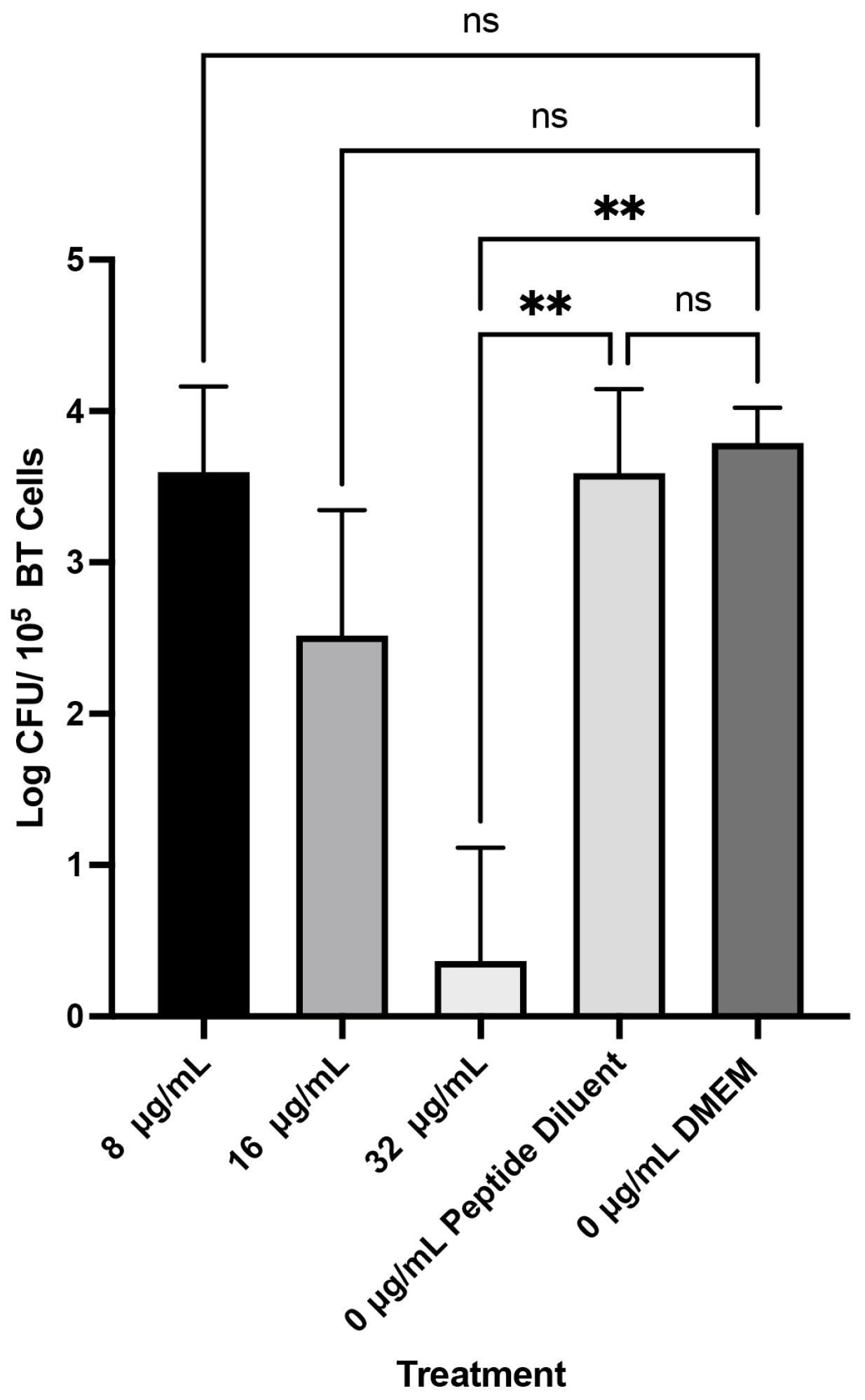
4.7. Attachment of M. haemolytica to Bovine Turbinates
4.8. Ability of WK2 to Prevent Attachment of M. haemolytica to Bovine Turbinates
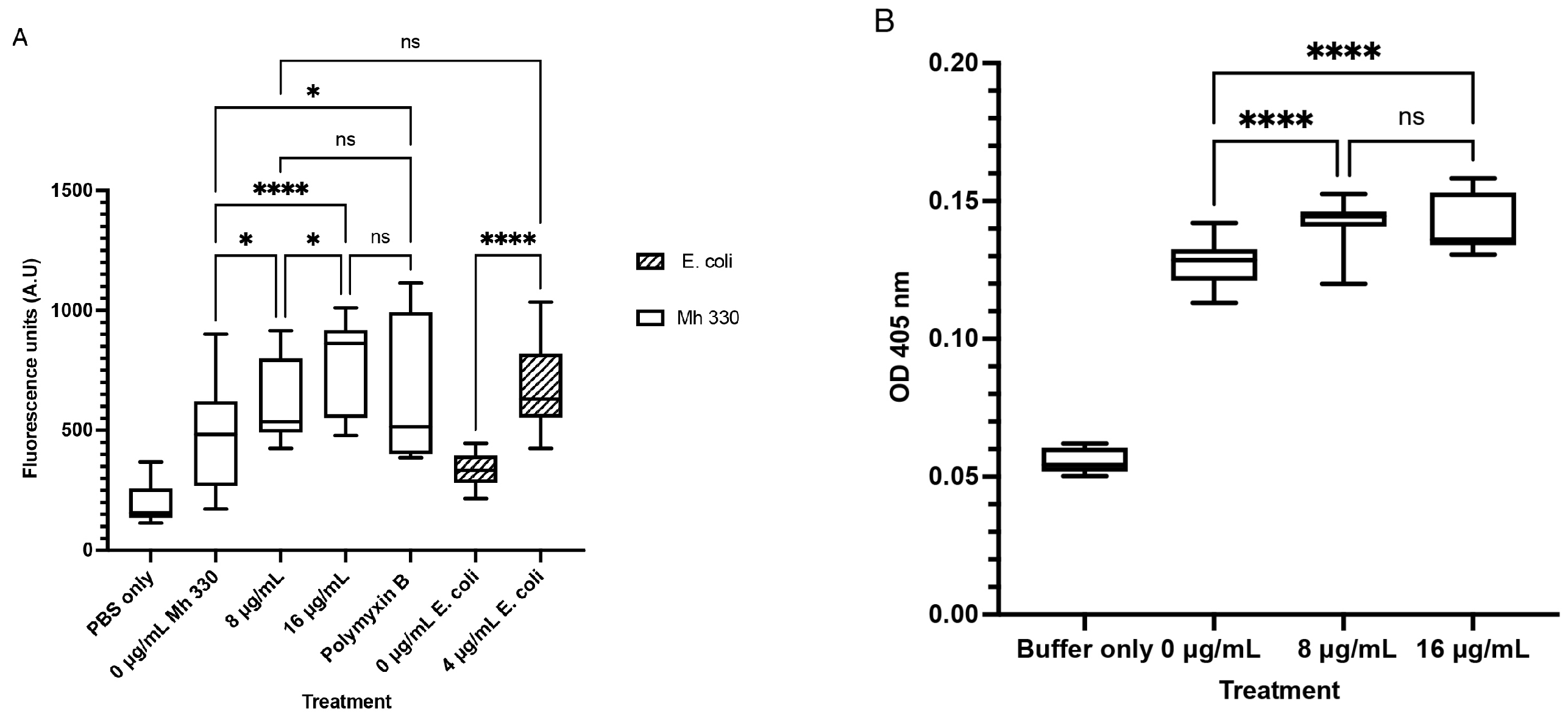
4.9. Membrane Permeability
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Andrés-Lasheras, S.; Ha, R.; Zaheer, R.; Lee, C.; Booker, C.W.; Dorin, C.; Van Donkersgoed, J.; Deardon, R.; Gow, S.; Hannon, S.J.; et al. Prevalence and Risk Factors Associated with Antimicrobial Resistance in Bacteria Related to Bovine Respiratory Disease—A Broad Cross-Sectional Study of Beef Cattle at Entry into Canadian Feedlots. Front. Vet. Sci. 2021, 8, 692646. [Google Scholar] [CrossRef] [PubMed]
- Gershwin, L.J.; Van Eenennaam, A.L.; Anderson, M.L.; McEligot, H.A.; Shao, M.X.; Toaff-Rosenstein, R.; Taylor, J.F.; Neibergs, H.L.; Womack, J. Bovine Respiratory Disease Complex Coordinated Agricultural Project Research Team Single Pathogen Challenge with Agents of the Bovine Respiratory Disease Complex. PLoS ONE 2015, 10, e0142479. [Google Scholar] [CrossRef] [PubMed]
- Mason, C.; Errington, J.; Foster, G.; Thacker, J.; Grace, O.; Baxter-Smith, K. Mannheimia Haemolytica Serovars Associated with Respiratory Disease in Cattle in Great Britain. BMC Vet. Res. 2022, 18, 5. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, A.M.; Hu, D.; Totton, S.C.; Scott, N.; Winder, C.B.; Wang, B.; Wang, C.; Glanville, J.; Wood, H.; White, B.; et al. A Systematic Review and Network Meta-Analysis of Bacterial and Viral Vaccines, Administered at or near Arrival at the Feedlot, for Control of Bovine Respiratory Disease in Beef Cattle. Anim. Health Res. Rev. 2019, 20, 143–162. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, A.M.; Coetzee, J.F.; da Silva, N.; Wang, C. A Mixed Treatment Comparison Meta-Analysis of Antibiotic Treatments for Bovine Respiratory Disease. Prev. Vet. Med. 2013, 110, 77–87. [Google Scholar] [CrossRef]
- Timsit, E.; Hallewell, J.; Booker, C.; Tison, N.; Amat, S.; Alexander, T.W. Prevalence and Antimicrobial Susceptibility of Mannheimia haemolytica, Pasteurella multocida, and Histophilus somni Isolated from the Lower Respiratory Tract of Healthy Feedlot Cattle and Those Diagnosed with Bovine Respiratory Disease. Vet. Microbiol. 2017, 208, 118–125. [Google Scholar] [CrossRef]
- Veronica, I.; Munoz, Kendall, L. Samuelson PAS, Dexter, J. Tomczak PAS, Hannah, A.; Seiver, Taylor, M. Smock PAS, John, T. Richeson Comparative Efficacy of Metaphylaxis with Tulathromycin and Pentavalent Modified-Live Virus Vaccination in High-Risk, Newly Received Feedlot Cattle. Appl. Anim. Sci. 2020, 36, 799–807. [Google Scholar]
- McAllister, H.; Halsted, S.; Parks, T.; Streeter, M.N.; Hutcheson, J.P.; Richeson, J. Effects of Intranasal or Parenteral Respiratory Vaccination Administered on Arrival or Delayed in Auction-derived Feedlot Heifers. J. Anim. Sci. 2021, 99, 38–39. [Google Scholar] [CrossRef]
- Fasina, Y.O.; Obanla, T.; Dosu, G.; Muzquiz, S. Significance of Endogenous Antimicrobial Peptides on the Health of Food Animals. Front. Vet. Sci. 2021, 8, 585266. [Google Scholar] [CrossRef]
- Skerlavaj, B.; Romeo, D.; Gennaro, R. Rapid Membrane Permeabilization and Inhibition of Vital Functions of Gram-Negative Bacteria by Bactenecins. Infect. Immun. 1990, 58, 3724–3730. [Google Scholar] [CrossRef]
- Rodrigues, G.; Souza Santos, L.; Franco, O.L. Antimicrobial Peptides Controlling Resistant Bacteria in Animal Production. Front. Microbiol. 2022, 13, 874153. [Google Scholar] [CrossRef] [PubMed]
- Fjell, C.D.; Jenssen, H.; Hilpert, K.; Cheung, W.A.; Panté, N.; Hancock, R.E.W.; Cherkasov, A. Identification of Novel Antibacterial Peptides by Chemoinformatics and Machine Learning. J. Med. Chem. 2009, 52, 2006–2015. [Google Scholar] [CrossRef]
- Dijksteel, G.S.; Ulrich, M.M.W.; Middelkoop, E.; Boekema, B.K.H.L. Review: Lessons Learned from Clinical Trials Using Antimicrobial Peptides (AMPs). Front. Microbiol. 2021, 12, 616979. [Google Scholar] [CrossRef]
- Dhingra, H.; Kaur, K.; Singh, B. Engineering and Characterization of Human β-Defensin-3 and Its Analogues and Microcin J25 Peptides against Mannheimia Haemolytica and Bovine Neutrophils. Vet. Res. 2021, 52, 83. [Google Scholar] [CrossRef] [PubMed]
- Dassanayake, R.P.; Falkenberg, S.M.; Register, K.B.; Samorodnitsky, D.; Nicholson, E.M.; Reinhardt, T.A. Antimicrobial Activity of Bovine NK-Lysin-Derived Peptides on Mycoplasma Bovis. PLoS ONE 2018, 13, e0197677. [Google Scholar] [CrossRef]
- Vulikh, K.; Bassel, L.L.; Sergejewich, L.; Kaufman, E.I.; Hewson, J.; MacInnes, J.I.; Tabatabaei, S.; Caswell, J.L. Effect of Tracheal Antimicrobial Peptide on the Development of Mannheimia Haemolytica Pneumonia in Cattle. PLoS ONE 2019, 14, e0225533. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Yang, J.; Han, J.; Gao, L.; Liu, H.; Lu, Z.; Zhao, H.; Bie, X. Insights into the Antimicrobial Activity and Cytotoxicity of Engineered α-Helical Peptide Amphiphiles. J. Med. Chem. 2016, 59, 10946–10962. [Google Scholar] [CrossRef]
- Ma, Z.; Zhang, R.; Hai, D.; Lu, Z.; Lv, F.; Zhao, H.; Zhang, C.; McAllister, T.A.; Stanford, K.; Bie, X. Antibiofilm Activity and Modes of Action of a Novel β-Sheet Peptide against Multidrug-Resistant Salmonella Enterica. Food Res. Int. 2019, 125, 108520. [Google Scholar] [CrossRef]
- Zhu, X.; Dong, N.; Wang, Z.; Ma, Z.; Zhang, L.; Ma, Q.; Shan, A. Design of Imperfectly Amphipathic α-Helical Antimicrobial Peptides with Enhanced Cell Selectivity. Acta Biomater. 2014, 10, 244–257. [Google Scholar] [CrossRef]
- Kyte, J.; Doolittle, R.F. A Simple Method for Displaying the Hydropathic Character of a Protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef]
- Scocchi, M.; Zelezetsky, I.; Benincasa, M.; Gennaro, R.; Mazzoli, A.; Tossi, A. Structural Aspects and Biological Properties of the Cathelicidin PMAP-36. FEBS J. 2005, 272, 4398–4406. [Google Scholar] [CrossRef] [PubMed]
- Cornut, I.; Büttner, K.; Dasseux, J.L.; Dufourcq, J. The Amphipathic Alpha-Helix Concept. Application to the de Novo Design of Ideally Amphipathic Leu, Lys Peptides with Hemolytic Activity Higher than that of Melittin. FEBS Lett. 1994, 349, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Klima, C.L.; Cook, S.R.; Zaheer, R.; Laing, C.; Gannon, V.P.; Xu, Y.; Rasmussen, J.; Potter, A.; Hendrick, S.; Alexander, T.W.; et al. Comparative Genomic Analysis of Mannheimia Haemolytica from Bovine Sources. PLoS ONE 2016, 11, e0149520. [Google Scholar] [CrossRef] [PubMed]
- Stanford, K.; Zaheer, R.; Klima, C.; McAllister, T.; Peters, D.; Niu, Y.D.; Ralston, B. Antimicrobial Resistance in Members of the Bacterial Bovine Respiratory Disease Complex Isolated from Lung Tissue of Cattle Mortalities Managed with or without the Use of Antimicrobials. Microorganisms 2020, 8, 288. [Google Scholar] [CrossRef] [PubMed]
- Lázár, V.; Martins, A.; Spohn, R.; Daruka, L.; Grézal, G.; Fekete, G.; Számel, M.; Jangir, P.K.; Kintses, B.; Csörgő, B.; et al. Antibiotic-Resistant Bacteria Show Widespread Collateral Sensitivity to Antimicrobial Peptides. Nat. Microbiol. 2018, 3, 718–731. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Han, J.; Chang, B.; Gao, L.; Lu, Z.; Lu, F.; Zhao, H.; Zhang, C.; Bie, X. Membrane-Active Amphipathic Peptide WRL3 with in Vitro Antibiofilm Capability and in Vivo Efficacy in Treating Methicillin-Resistant Staphylococcus Aureus Burn Wound Infections. ACS Infect. Dis. 2017, 3, 820–832. [Google Scholar] [CrossRef] [PubMed]
- Dathe, M.; Nikolenko, H.; Meyer, J.; Beyermann, M.; Bienert, M. Optimization of the Antimicrobial Activity of Magainin Peptides by Modification of Charge. FEBS Lett. 2001, 501, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Yeaman, M.R.; Yount, N.Y. Mechanisms of Antimicrobial Peptide Action and Resistance. Pharmacol. Rev. 2003, 55, 27–55. [Google Scholar] [CrossRef] [PubMed]
- Trimble, M.J.; Mlynárčik, P.; Kolář, M.; Hancock, R.E.W. Polymyxin: Alternative Mechanisms of Action and Resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a025288. [Google Scholar] [CrossRef]
- Shagaghi, N.; Palombo, E.A.; Clayton, A.H.A.; Bhave, M. Antimicrobial Peptides: Biochemical Determinants of Activity and Biophysical Techniques of Elucidating Their Functionality. World J. Microbiol. Biotechnol. 2018, 34, 62. [Google Scholar] [CrossRef]
- Boukahil, I.; Czuprynski, C.J. Mannheimia Haemolytica Biofilm Formation on Bovine Respiratory Epithelial Cells. Vet. Microbiol. 2016, 197, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Sandal, I.; Shao, J.Q.; Annadata, S.; Apicella, M.A.; Boye, M.; Jensen, T.K.; Saunders, G.K.; Inzana, T.J. Histophilus Somni Biofilm Formation in Cardiopulmonary Tissue of the Bovine Host Following Respiratory Challenge. Microbes Infect. 2009, 11, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Morck, D.W.; Olson, M.E.; Acres, S.D.; Daoust, P.Y.; Costerton, J.W. Presence of Bacterial Glycocalyx and Fimbriae on Pasteurella Haemolytica in Feedlot Cattle with Pneumonic Pasteurellosis. Can. J. Vet. Res. 1989, 53, 167–171. [Google Scholar] [PubMed]
- Olson, M.E.; Ceri, H.; Morck, D.W.; Buret, A.G.; Read, R.R. Biofilm Bacteria: Formation and Comparative Susceptibility to Antibiotics. Can. J. Vet. Res. 2002, 66, 86–92. [Google Scholar] [PubMed]
- Cozens, D.; Sutherland, E.; Lauder, M.; Taylor, G.; Berry, C.C.; Davies, R.L. Pathogenic Mannheimia Haemolytica Invades Differentiated Bovine Airway Epithelial Cells. Infect. Immun. 2019, 87, e00078-19. [Google Scholar] [CrossRef] [PubMed]
- Amat, S.; Timsit, E.; Baines, D.; Yanke, J.; Alexander, T.W. Development of Bacterial Therapeutics against the Bovine Respiratory Pathogen Mannheimia Haemolytica. Appl. Environ. Microbiol. 2019, 85, e01359-19. [Google Scholar] [CrossRef] [PubMed]
- Amat, S.; Alexander, T.W.; Holman, D.B.; Schwinghamer, T.; Timsit, E. Intranasal Bacterial Therapeutics Reduce Colonization by the Respiratory Pathogen Mannheimia Haemolytica in Dairy Calves. mSystems 2020, 5, 988. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Stanford, K.; McAllister, T.A.; Johnson, R.P.; Chen, J.; Hou, H.; Zhang, G.; Niu, Y.D. Biofilm Formation, Virulence Gene Profiles, and Antimicrobial Resistance of Nine Serogroups of Non-O157 Shiga Toxin-Producing Escherichia coli. Foodborne Pathog. Dis. 2016, 13, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Loh, B.; Grant, C.; Hancock, R.E. Use of the Fluorescent Probe 1-N-Phenylnaphthylamine to Study the Interactions of Aminoglycoside Antibiotics with the Outer Membrane of Pseudomonas Aeruginosa. Antimicrob. Agents Chemother. 1984, 26, 546–551. [Google Scholar] [CrossRef]
- Schaffner, D.W. Predictive Food Microbiology Gedanken Experiment: Why Do Microbial Growth Data Require a Transformation? Food Microbiol. 1998, 15, 185–189. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Sequence | Structure | Molecular Weight | Net Charge | MIC E. coli ATCC 25,922 µg/mL | Parent |
|---|---|---|---|---|---|---|
| PRW4 | RFRRLRWKTRWRLKKI-NH2 | α-helical | 2298.89 | +10 | 9.2 | PMAP-36 |
| WRL3 | WLRAFRRLVRRLARGLRR-NH2 | α-helical | 2350.92 | +9 | 4.0 | Leucocin A |
| WK2 | WKWKCTKSGCKWKW-NH2 | β-sheet | 1854.28 | +6 | 4 | Leucocin A |
| Strains | Serotype | Sensititre Resistance Profile |
|---|---|---|
| Mh 276 | 1 | SDM, SXT, CLI, TYLT, SPE |
| Mh 330 | 1 | SDM, CLI, TYLT |
| Mh 13 | 1 | OXY, SDM, SXT, TIL, TUL, CLI, NEO, TYLT |
| Mh 535 | 1 | Susceptible to all antibiotics tested |
| Mh 587 | 2 | Susceptible to all antibiotics tested |
| Mh 136 | 1 | AMP, CTET, FFN, GEN, OXY, SDM, TIL, TUL, CLI, DANO, ENRO, NEO, PEN, TYLT |
| E. coli ATCC25922 | O6 biotype 1 | N/A |
| Strain | PRW4 MIC (µg/mL) | WRL3 MIC (µg/mL) | WRL3 MBC (µg/mL) | WK2 MIC (µg/mL) | WK2 MBC (µg/mL) |
|---|---|---|---|---|---|
| Mh 330 | >256 | 16–32 | 32 | 8–16 | 8–16 |
| Mh 587 | 64 | 2 | 16 | 2–4 | 4 |
| Mh 276 | 256 | 16–32 | 32 | 16 | 16 |
| Mh 136 | >256 | 16 | 16 | 32 | 32 |
| Mh 13 | >256 | 16–32 | 32 | 16–32 | >128 |
| Mh 535 | >256 | 4–8 | 8 | 16 | 16 |
| E. coli ATCC 25922 | 8 | 32 | 32 | 4 | 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bao, R.; Ma, Z.; Stanford, K.; McAllister, T.A.; Niu, Y.D. Antimicrobial Activities of α-Helix and β-Sheet Peptides against the Major Bovine Respiratory Disease Agent, Mannheimia haemolytica. Int. J. Mol. Sci. 2024, 25, 4164. https://doi.org/10.3390/ijms25084164
Bao R, Ma Z, Stanford K, McAllister TA, Niu YD. Antimicrobial Activities of α-Helix and β-Sheet Peptides against the Major Bovine Respiratory Disease Agent, Mannheimia haemolytica. International Journal of Molecular Sciences. 2024; 25(8):4164. https://doi.org/10.3390/ijms25084164
Chicago/Turabian StyleBao, Ruina, Zhi Ma, Kim Stanford, Tim A. McAllister, and Yan D. Niu. 2024. "Antimicrobial Activities of α-Helix and β-Sheet Peptides against the Major Bovine Respiratory Disease Agent, Mannheimia haemolytica" International Journal of Molecular Sciences 25, no. 8: 4164. https://doi.org/10.3390/ijms25084164
APA StyleBao, R., Ma, Z., Stanford, K., McAllister, T. A., & Niu, Y. D. (2024). Antimicrobial Activities of α-Helix and β-Sheet Peptides against the Major Bovine Respiratory Disease Agent, Mannheimia haemolytica. International Journal of Molecular Sciences, 25(8), 4164. https://doi.org/10.3390/ijms25084164