
Influence of Abiotic and Biotic Elicitors on Organogenesis, Biomass Accumulation, and Production of Key Secondary Metabolites in Asteraceae Plants


Abstract
:1. Medicinal Plants from Asteraceae Family: Chemical Constituents and Applications
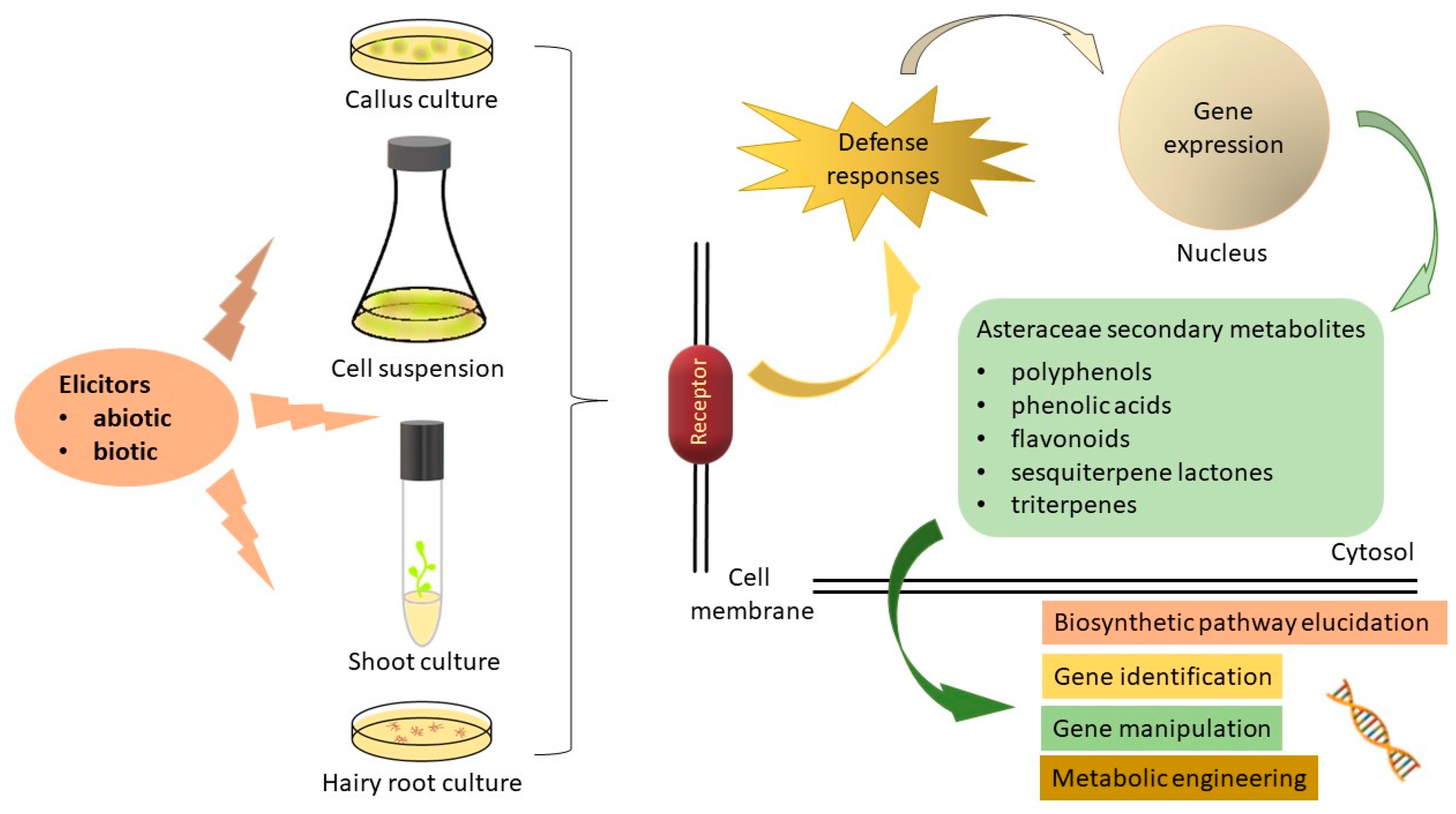
2. Elicitors
2.1. Abiotic Elicitors
2.1.1. Physical Elicitors
Osmotic Stress
Drought Stress
Salinity

{kind=link}
| Plant Species | Physical Elicitors | Type of In Vitro Culture | Effects | References |
|---|---|---|---|---|
| UV radiation | ||||
| Artemisia annua L. (sweet wormwood) | Light irradiation (3000 Lux for 16 h cool-white fluorescent lamps and then darkness for 8 h) | HR | Hairy roots’ growth and artemisinin accumulation increased. | [36] |
| Artemisia absinthium L. (common wormwood) | Different spectral lights | CC | Red spectrum enhanced peroxidase activity, protease activity, total protein content, and chlorophyll a/b ratio; green spectrum stimulated TPC, TFC, and AA; yellow light enhanced MDA content, while white and green light improved total chlorophyll content and carotenoid content. | [39] |
| Artemisia absinthium L. (common wormwood) | Light | SuspC | Maximum increase in dry biomass; high AA; enhanced levels of TPC and total secondary metabolites in light-grown suspension cultures during log phase of growth. | [38] |
| Stevia rebaudiana Bert. (stevia) | Different spectral lights | CC | Control white light improved callogenic frequency and biomass accumulation; blue light enhanced TPC and TFC; green and red light improved AA. | [37] |
| Stevia rabaudiana Bert. (stevia) | Light-emitting diodes’ (LEDs’) spectra | ShC | Blue LED light increased seed germination. Blue LED light stimulated the growth of roots and leaves of in vitro plantlets; and the number and opening of stomata. Red LED light increased stems and roots, but it had the least positive impact on the production of carotenoids and chlorophylls. Red and blue LED light have opposite effect on the activity of antioxidant enzymes (catalase (CAT), peroxidase (POD), and superoxide dismutase (SOD)). | [44] |
| Stevia rabaudiana Bert. (stevia) | Light-emitting diodes’ (LEDs’) spectra | ShC | Red LEDs enhanced proliferation rate; Blue/Red LED promoted shoot elongation. | [45] |
| Stevia rabaudiana Bert. (stevia) | Red LED | ShC | S. rebaudiana microshoots grown on MS media with 1.0 mg L−1 6-benzylaminopurine (BAP) and 0.5 mg L−1 Indole acetic acid (IAA) under red monochromatic light treatments showed a 30% increase in multiplication coefficient compared to control (white light, media without PGRs). | [46] |
| Stevia rabaudiana Bert. (stevia) | Light-emitting diodes’ (LEDs’) spectra | ShC | Blue LED light promoted shoot elongation, leaves number/shoot, leaf fresh weight, leaf dry weight, and photosynthetic pigment production; red LED treatment resulted in high shoot production; fluorescent light induced 100% root induction. | [47] |
| Stevia rabaudiana Bert. (stevia) | Spectral lights | AdvRC | Violet light showed maximum FW and highest DPPH inhibition, while blue light enhanced phenolic content and total flavonoid production. | [48] |
| Stevia rabaudiana Bert. (stevia) | LED treatment | ShC | LED source with an intensity of 75 and 230 µmol/(m2 s) enhanced FW accumulation of aerial parts and roots and improved ex vitro adaptation of plants. | [49] |
| Stevia rabaudiana Bert. (stevia) | Far-red LED induction system | ShC | Favorable impact on the growth of shoots and production of metabolites (stevioside and rebaudioside-A) by up to 37.15% and 22.99%, enhancing gene expression related to steviol glycosides. | [50] |
| Cichorium intybus L. (common chicory) | Artificial Light | CC | Radiation levels in the region of FR—far red > R—red promote callus fresh weight and inulin content in callus cells. | [51] |
| Silybum marianum L. (milk thistle) | Monochromatic lights | CC | High levels of silychristin, isosilychristin, silydianin, silybin A, and silybin B under red light; maximum content of isosilybin A and isosilybin B under green light; high amount of taxifolin under continuous white light. | [40] |
| Echinacea purpurea L. (echinacea) | UV-B treatments | CC; SuspC | Enhanced caffeic acid and growth parameters, TPC, and AA of cell suspensions. | [41] |
| Echinacea angustifolia DC. (blacksamson echinacea) | Continuous light | SuspC | Continuous light for 14 days significantly increased levels of certain caffeic acid derivatives and reduced level of hydroxytyrosol derivatives with rhamnose residues. | [42] |
| Echinacea purpurea L. (echinacea) | Light | HR | Increased levels of anthocyanins and caffeic acid derivative biosynthesis, stimulated the activity of phenylalanine ammonium lyase. | [43] |
| Osmotic stress | ||||
| Stevia rebaudiana Bert. (stevia) | Proline and PEG | CC;SC | Increased steviol glycoside synthesis. | [55] |
| Arnica montana L. (mountain arnica) | Sucrose Maltose Glucose | HR | Enhanced growth and biomass accumulation of Arnica montana hairy roots on MS nutrient medium containing 3% or 5% sucrose; sugars and sugar alcohol content were influenced by the concentration of respective carbon sources in the nutrient medium. | [57] |
| Arnica montana L. (mountain arnica) | Sorbitol and mannitol | ShC | Reduced plant growth; maintain plant quality for up to 6 months. | [59] |
| Artemisia annua L. (sweet wormwood) | Sorbitol and coronatine | SuspC | Sorbitol treatment enhanced malondialdehyde and hydrogen peroxide content; application of coronatine and sorbitol together increased the expression of artemisinin biosynthetic genes and artemisinin production at all tested concentrations. | [53] |
| Arnica montana L. (mountain arnica) | Medium salt content | ShC | The highest phenolic content and antioxidant activity are obtained in shoots grown on ¼ strength of MS medium. | [60] |
| Drought stress | ||||
| Stevia rebaudiana (stevia) | Agar-induced drought stress | ShC | The highest accumulation of proline and malondialdehyde are detected in the highest concentration of agar, but enhanced antioxidant enzyme activities are obtained on 8 g/L agar containing cultural medium. | [64] |
| Silybum marianum (milk thistle) | Mannitol- induced drought stress | ShC | Inhibited root induction; retarded plant growth; enhanced accumulation of TPC, TFC, and total protein content along with several antioxidative enzymes. | [65] |
| Salinity | ||||
| Stevia rebaudiana Bert. (stevia) | NaCl | ShC | Increased AA, hydroxycinnamic acid, and total soluble sugar content; induced changes in expression of steviol glycoside-related biosynthetic genes. | [77] |
| Stevia rebaudiana Bert. (stevia) | NaCl | CC, ShC | Steviol glycosides rebaudioside A and stevioside, as well as TPC, TFC, and AA, showed significant improvement. | [78] |
| Stevia rebaudiana Bert. (stevia) | NaCl concentrations: 500, 1000, 2000, and 3000 mg L−1 | CC, ShC | Salinity has a negative effect on shoot number, shoot length, root number, root length, leaf number, and total chlorophyll content; biochemical markers peroxidase (PDO), polyphenol oxidase (PPO), and malate dehydrogenase (MDH) were associated with salt tolerance. | [79] |
| Artemisia arborescens L. (wormwood) | NaCl or KCl | ShC | Improvement in TPC, TFC, condensed tannins, and volatile compounds accumulation. | [80] |
2.1.2. Plant Growth Regulators
Jasmonic Acid and Methyl Jasmonate
Salicylic Acid
| Plant Species | Elicitors | Type of In Vitro Culture | Effects | References |
|---|---|---|---|---|
| Plant growth regulators | ||||
| Artemisia annua L. (sweet wormwood) | MeJA and mevalonic acid lactone as a precursor | SuspC | 5.93 times higher artemisinin productivity compared to control cultures | [88] |
| Artemisia annua L. (sweet wormwood) | MeJA (40 μg/L) and the precursors casein acid hydrolysate (50 μg/L) and sodium acetate (500 μg/L) | HR | A maximum artemisinin content of 3.45 mg/g on 15 days after treatment | [92] |
| Artemisia annua L. (sweet wormwood) | β-cyclodextrin (50 mM) and MeJA (100 μM) | SuspC | About 300 times more artemisinin than the control | [93] |
| Artemisia annua L. (sweet wormwood) | MeJA (100 μM) and cell homogenate of Piriformospora indica (3% v/v) | HR | The enhancement of 2.44 times in artemisinin concentration in the hairy roots; positive correlation with regulatory genes of the MVA, MEP, and artemisinin biosynthetic pathways, viz. hmgr, ads, cyp71av1, aldh1, dxs, dxr, and dbr2 | [95] |
| Artemisia annua L. (sweet wormwood) | Twenty-two micromolar MeJA | SuspC | Three-fold increase in artemisinin production in around 30 min; the MeJA-induced upregulation of CYP71AV1 | [91] |
| Silybum marianum L. (milk thistle) | MeJA and methyl B cyclodextrin | SuspC | Silymarin accumulation increased | [99] |
| Silybum marianum L. (milk thistle) | MeJA | CC, HR | Increase the total silymarin | [98] |
| Silybum marianum L. (milk thistle) | MeJA | CC | MeJA promoted the accumulation of silymarin and enhanced chalcone synthase (CHS) activity | [96] |
| Silybum marianum L. (milk thistle) | MeJA (100 μM), YE (50 μg.mL−1), and Phe (0.1 mM) | CC | The enhancement of the silymarin production - 8.6 times higher than the control | [97] |
| Echinacea purpurea L. (echinacea) | MeJA 7.5 mg/L | SuspC | Chicoric acid accumulation; C3H expression appeared correlated with the enhanced biosynthesis of chicoric acid in E. purpurea cell suspension and significantly boosted by MeJA elicitation | [100] |
| Echinacea purpurea L. (echinacea) | MeJA 100 μM | ShC | Raised the caffeic acid derivatives in both shoots and roots; growth parameters of the shoot and roots declined in line with the rising level of MeJA treatments | [101] |
| Echinacea purpurea L. (echinacea) | MeJA | CC; ShC; RC | Rise in TPC and CAD accumulation, particularly at concentrations of 100 and 150 µM MeJA; inhibited biomass and growth indexes in all callus, shoot, and root cultures | [103] |
| Echinacea purpurea L. (echinacea) and E. pallida Nutt. (pale purple coneflower) | 25 μM MeJA | AdvRC | The maximum production of phenolics, flavonoids, and caffeic acid derivatives (cichoric acid and echinacoside); maximum levels of activities of antioxidant enzymes (superoxide dismutase, peroxidase, ascorbate peroxidase, and catalase) with 25 μM MeJA | [104] |
| Cichorium intybus L. (common chicory) | MeJA 100 μM | HR | High sesquiterpene lactone accumulation | [105] |
| Cichorium intybus L. (common chicory) | MeJA | HR | High 3,5-dicaffeoylquinic acid; the biomass of hairy roots and rates of CQAs are higher than in plants and other hairy root cultures | [106] |
| Artemisia absinthium L. (common wormwood) | MeJA, JA and gibberellic acid (GA) | SuspC | Biomass accumulation was inhibited by the application of elicitors but enhanced the accumulation of TPC, TFC, and high AA | [107] |
| Artemisia amygdalina Decne. | Methyl jasmonate | AdvRC | The higher production of TPC, TFC, and phenylalanine ammonia lyase activity | [108] |
| Tanacetum parthenium L. (feverfew) | 2.5 mg/L YE and 100 µM MeJA | HR | Enhanced parthenolide production and the expression of the parthenolide synthase (TpPTS) gene | [109] |
| Tanacetum parthenium L. (feverfew) | 2.5 mg/L YE and 0.5 mg/L MeJA | SuspC | The highest parthenolide accumulation is achieved in the cell suspension containing 0.5 mg/L 2,4-D and 0.1 mg/L TDZ treated with YE + MeJA elicitor for 48 h | [110] |
| Stevia rebaudiana Bert. (stevia) | Salicylic acid (SA) and MeJA | ShC | The maximum shoot growth, root regeneration, and FW and DW accumulation on WPM containing 50 μM SA; WPM containing MeJA and SA increased SOD2 and POX1 activity, TPC, and AA and decreased CAT activity; the highest levels of enzymatic and non-enzymatic antioxidants were observed in the WPM containing 100 μM SA; 50 μM MeJA and 100 μM SA enhanced stevioside production | [113] |
| Stevia rebaudiana Bert. (stevia) | SA | CC | SA elicitation, (0.75 mg/L) promoted callus growth rate, callus diameter, and relative callus FW; the addition of 0.25 (mg/L) of SA to the MS medium led to the production of the highest amount of rebaudioside A | [114] |
| Artemisia aucheri Boiss. | SA | ShC | SA pretreatment decreased the effects of PEG-simulated drought stress under in vitro conditions by improving the activity of antioxidant enzymes | [116] |
| Echinacea purpurea L. (echinacea) | SA | ShC | The highest average number of branches formed (33.6 branches/plant part), leaves (33.6 leaves/vegetable part), fresh 1.067 g and dry 0.058 g weights, and a 291.3427 μg/g DW concentration of echinacoside | [117] |
2.1.3. Nanoparticles (NP)
2.2. Biotic Elicitors
3. Conclusions and Future Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Devkota, H.P. An overview of medicinal plants of the Asteraceae family and their role in human health. In Medicinal Plants of the Asteraceae Family: Traditional Uses, Phytochemistry and Pharmacological Activities; Devkota, H.P., Aftab, T., Eds.; Springer: Singapore, 2022; pp. 1–15. [Google Scholar] [CrossRef]
- Bessada, S.M.; Barreira, J.C.; Oliveira, M.B.P. Asteraceae species with most prominent bioactivity and their potential applications: A review. Ind. Crops Prod. 2015, 76, 604–615. [Google Scholar] [CrossRef]
- Rolnik, A.; Olas, B. The Plants of the Asteraceae Family as agents in the protection of human health. Int. J. Mol. Sci. 2021, 22, 3009. [Google Scholar] [CrossRef] [PubMed]
- Septembre-Malaterre, A.; Lalarizo Rakoto, M.; Marodon, C.; Bedoui, Y.; Nakab, J.; Simon, E.; Hoarau, L.; Savriama, S.; Strasberg, D.; Guiraud, P.; et al. Artemisia annua, a traditional plant brought to light. Int. J. Mol. Sci. 2020, 21, 4986. [Google Scholar] [CrossRef] [PubMed]
- Kriplani, P.; Guarve, K.; Baghael, U.S. Arnica montana L.—A plant of healing. J. Pharm. Pharmacol. 2017, 69, 925–945. [Google Scholar] [CrossRef] [PubMed]
- Šutovská, M.; Capek, P.; Kočmalová, M.; Pawlaczyk, I.; Zaczyńska, E.; Czarny, A.; Uhliariková, I.; Gancarz, R.; Fraňová, S. Characterization and pharmacodynamic properties of Arnica montana complex. Int. J. Biol. Macromol. 2014, 69, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.D.; Rao, G.P. Stevia: The herbal sugar of 21st century. Sugar Tech 2005, 7, 17–24. [Google Scholar] [CrossRef]
- Tavarini, S.; Angelini, L.G. Stevia rebaudiana Bertoni as a source of bioactive compounds: The effect of harvest time, experimental site and crop age on steviol glycoside content and antioxidant properties. J. Sci. Food Agric. 2013, 93, 2121–2129. [Google Scholar] [CrossRef]
- Peteliuk, V.; Rybchuk, L.; Bayliak, M.; Storey, K.B.; Lushchak, O. Natural sweetener Stevia rebaudiana: Functionalities, health benefits and potential risks. EXCLI J. 2021, 20, 1412. [Google Scholar] [CrossRef] [PubMed]
- Manayi, A.; Vazirian, M.; Saeidnia, S. Echinacea purpurea: Pharmacology, phytochemistry and analysis methods. Pharmacogn Rev. 2015, 9, 63–72. [Google Scholar] [CrossRef]
- Burlou-Nagy, C.; Bănică, F.; Jurca, T.; Vicaș, L.G.; Marian, E.; Muresan, M.E.; Bácskay, I.; Kiss, R.; Fehér, P.; Pallag, A. Echinacea purpurea (L.) Moench: Biological and pharmacological properties. A review. Plants 2022, 11, 1244. [Google Scholar] [CrossRef]
- Al-Snafi, A.E. Medical importance of Cichorium intybus–A review. IOSR J. Pharm. 2016, 6, 41–56. [Google Scholar]
- Perović, J.; Tumbas Šaponjac, V.; Kojić, J.; Krulj, J.; Moreno, D.A.; García-Viguera, C.; Bodroža-Solarov, M.; Ilić, N. Chicory (Cichorium intybus L.) as a food ingredient—Nutritional composition, bioactivity, safety, and health claims: A review. Food Chem. 2021, 336, 127676. [Google Scholar] [CrossRef] [PubMed]
- Karkanis, A.; Bilalis, D.; Efthimiadou, A. Cultivation of milk thistle (Silybum marianum L. Gaertn.), a medicinal weed. Ind. Crops Prod. 2011, 34, 825–830. [Google Scholar] [CrossRef]
- Vaknin, Y.; Hadas, R.; Schafferman, D.; Murkhovsky, L.; Bashan, N. The potential of milk thistle (Silybum marianum L.), an Israeli native, as a source of edible sprouts rich in antioxidants. Int. J. Food Sci. Nutr. 2008, 59, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Pareek, A.; Suthar, M.; Rathore, G.S.; Bansal, V. Feverfew (Tanacetum parthenium L.): A systematic review. Pharmacogn. Rev. 2011, 5, 103. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.R.; Ravishankar, G.A. Plant cell cultures: Chemical factories of secondary metabolites. Biotechnol. Adv. 2002, 20, 101–153. [Google Scholar] [CrossRef]
- Thakur, G.S.; Sharma, R.; Sanodiya, B.S.; Baghel, R.; Thakur, R.; Singh, B.N.; Savita, A.; Dubey, A.; Sikarwar, L.; Jaiswal, P.; et al. In vitro induction of tuber formation for the synthesis of secondary metabolites in Chlorophytum borivilianum Sant. et Fernand. Afr. J. Biotechnol. 2013, 12, 2900–2907. [Google Scholar]
- Rea, G.; Antonacci, A.; Lambreva, M.; Pastorelli, S.; Tibuzzi, A.; Ferrari, S.; Fischer, D.; Johanningmeier, U.; Oleszek, W.; Doroszewska, T. Integrated plant biotechnologies applied to safer and healthier food production: The Nutra-Snack manufacturing chain. Trends Food Sci. Technol. 2011, 22, 353–366. [Google Scholar] [CrossRef]
- DiCosmo, F.; Misawa, M. Eliciting secondary metabolism in plant cell cultures. Trends Biotech. 1985, 3, 318–322. [Google Scholar] [CrossRef]
- Singh, A.; Dwivedi, P. Methyl-jasmonate and salicylic acid as potent elicitors for secondary metabolite production in medicinal plants: A review. J. Pharmacogn. Phytochem. 2018, 7, 750–757. [Google Scholar]
- Zhao, J.; Davis, L.C.; Verpoorte, R. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 2005, 23, 283–333. [Google Scholar] [CrossRef]
- Thiruvengadam, M.; Praveen, N.; Kim, E.H.; Kim, S.H.; Chung, I.M. Production of anthraquinones, phenolic compounds and biological activities from hairy root cultures of Polygonum multiflorum Thunb. Protoplasma 2014, 251, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.J.; Park, S.Y.; Paek, K.Y. Enhancement strategies of bioactive compound production in adventitious root cultures of Eleutherococcus koreanum Nakai subjected to methyl jasmonate and salicylic acid elicitation through airlift bioreactors. Plant Cell Tissue Organ. Cult. 2015, 120, 1–10. [Google Scholar] [CrossRef]
- Bhaskar, R.; Xavier, L.S.E.; Udayakumaran, G.; Kumar, D.S.; Venkatesh, R.; Nagella, P. Biotic elicitors: A boon for the in-vitro production of plant secondary metabolites. Plant Cell Tiss. Organ. Cult. 2022, 149, 7–24. [Google Scholar] [CrossRef]
- Narayani, M.; Shrivastava, S. Elicitation: A stimulation of stress in in vitro plant cell/tissue cultures for enhancement of secondary metabolite production. Phytochemical. Rev. 2017, 16, 1227–1252. [Google Scholar] [CrossRef]
- Thakur, M.; Bhattacharya, S.; Kumar Khosla, P.; Puri, S. Improving production of plant secondary metabolites through biotic and abiotic elicitation. J. Appl. Res. Med. Aromat. Plants 2020, 12, 1–12. [Google Scholar] [CrossRef]
- Naik, P.M.; Al-Khayri, J.M. Abiotic and biotic elicitors–role in secondary metabolites production through in vitro culture of medicinal plants. In Abiotic and Biotic Stress in Plants-Recent Advances and Future Perspectives; Books on Demand: Norderstedt, Germany; Rijeka, Croatia, 2016; pp. 247–277. [Google Scholar]
- Cirak, C.; Radušienė, J.; Kurtarc, E.S.; Marksa, M.; Ivanauskas, L. In vitro plant regeneration and jasmonic acid induced bioactive chemical accumulations in two Hypericum species from Turkey. S. Afr. J. Bot. 2020, 128, 312–318. [Google Scholar] [CrossRef]
- Sirhindi, G.; Mushtaq, R.; Gill, S.S.; Sharma, P.; Allah, E.F.A.; Ahmad, P. Jasmonic acid and methyl jasmonate modulate growth, photosynthetic activity and expression of photosystem II subunit genes in Brassica oleracea L. Sci. Rep. 2020, 10, 9322. [Google Scholar] [CrossRef]
- Murthy, H.N.; Lee, E.J.; Paek, K.Y. Production of secondary metabolites from cell and organ cultures: Strategies and approaches for biomass improvement and metabolite accumulation. Plant Cell Tissue Organ Cult. 2014, 118, 1–16. [Google Scholar] [CrossRef]
- Kundu, K.; Roy, A.; Saxena, G.; Kumar, L.; Bharadvaja, N. Effect of different carbon sources and elicitors on shoot multiplication in accessions of Centella asiatica. Med. Aromat. Plants 2016, 5, 1000251. [Google Scholar] [CrossRef]
- Putalun, W.; Udomsin, O.; Yusakul, G.; Juengwatanatrakul, T.; Sakamoto, S.; Tanaka, H. Enhanced plumbagin production from in vitro cultures of Drosera burmanii using elicitation. Biotechnol. Lett. 2010, 32, 721–724. [Google Scholar] [CrossRef] [PubMed]
- Sivanandhan, G.; Rajesh, M.; Arun, M.; Jeyaraj, M.; Kapil Dev, G.; Arjunan, A.; Manickavasagam, M.; Muthuselvam, M.; Selvaraj, N.; Ganapathi, A. Effect of culture conditions, cytokinins, methyl jasmonate and salicylic acid on the biomass accumulation and production of withanolides in multiple shoot culture of Withania somnifera (L.) dunal using liquid culture. Acta Physiol. Plant. 2013, 35, 715–728. [Google Scholar] [CrossRef]
- Hashim, M.; Ahmad, B.; Drouet, S.; Hano, C.; Abbasi, B.H.; Anjum, S. Comparative effects of different light sources on the production of key secondary metabolites in plants in vitro cultures. Plants 2021, 10, 1521. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.Z.; Guo, C.; Wang, Y.C.; Ouynag, F. Effect of light irradiation on hairy root growth and artemisinin biosynthesis of Artemisia annua L. Process. Biochem. 2002, 38, 581–585. [Google Scholar] [CrossRef]
- Ahmad, N.; Rab, A.; Ahmad, N. Light-induced biochemical variations in secondary metabolite production and antioxidant activity in callus cultures of Stevia rebaudiana (Bert). J. Photochem. Photobiol. B Biol. 2016, 154, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Abbasi, B.H. Light-induced fluctuations in biomass accumulation, secondary metabolites production and antioxidant activity in cell suspension cultures of Artemisia absinthium L. J. Photochem. Photobiol. B Biol. 2014, 140, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Tariq, U.; Ali, M.; Abbasi, B.H. Morphogenic and biochemical variations under different spectral lights in callus cultures of Artemisia absinthium L. J. Photochem. Photobiol. B Biol. 2014, 130, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Younas, M.; Drouet, S.; Nadeem, M.; Giglioli-Guivarc’h, N.; Hano, C.; Abbasi, B.H. Differential accumulation of silymarin induced by exposure of Silybum marianum L. callus cultures to several spectres of monochromatic lights. J. Photochem. Photobiol. B Biol. 2018, 184, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Manaf, H.H.; Rabie, K.A.E.; Abd El-Aal, M.S. Impact of UV-B radiation on some biochemical changes and growth parameters in Echinacea purpurea callus and suspension culture. Ann. Agri. Sci. 2016, 61, 207–216. [Google Scholar] [CrossRef]
- Guarnerio, C.F.; Fraccaroli, M.; Gonzo, I.; Pressi, G.; Dal Toso, R.; Guzzo, F.; Levi, M. Metabolomic analysis reveals that the accumulation of specific secondary metabolites in Echinacea angustifolia cells cultured in vitro can be controlled by light. Plant Cell Rep. 2012, 31, 361–367. [Google Scholar] [CrossRef]
- Abbasi, B.H.; Tian, C.-L.; Murch, S.J.; Saxena, P.K.; Liu, C.-Z. Light-enhanced caffeic acid derivatives biosynthesis in hairy root cultures of Echinacea purpurea. Plant Cell Rep. 2007, 26, 1367–1372. [Google Scholar] [CrossRef] [PubMed]
- Simlat, M.; Ślęzak, P.; Moś, M.; Warchoł, M.; Skrzypek, E.; Ptak, A. The effect of light quality on seed germination, seedling growth and selected biochemical properties of Stevia rebaudiana Bertoni. Sci. Hortic. 2016, 211, 295–304. [Google Scholar] [CrossRef]
- Ramírez-Mosqueda, M.A.; Iglesias-Andreu, L.G.; Bautista-Aguilar, J.R. The effect of light quality on growth and development of in vitro plantlet of Stevia rebaudiana Bertoni. Sugar Tech 2017, 19, 331–336. [Google Scholar] [CrossRef]
- Shulgina, A.A.; Kalashnikova, E.A.; Tarakanov, I.G.; Kirakosyan, R.N.; Cherednichenko, M.Y.; Polivanova, O.B.; Baranova, E.N.; Khaliluev, M.R. Influence of light conditions and medium composition on morphophysiological characteristics of Stevia rebaudiana Bertoni in vitro and in vivo. Horticulturae 2021, 7, 195. [Google Scholar] [CrossRef]
- Attaya, A.S. LED Light technology as a source of illumination and a promising method for Stevia rebaudiana elite propagation. Egypt J. Agron. 2021, 43, 123–132. [Google Scholar] [CrossRef]
- Idrees, M.; Sania, B.; Hafsa, B.; Kumari, S.; Khan, H.; Fazal, H.; Ahmad, I.; Akbar, F.; Ahmad, N.; Ali, S.; et al. Spectral lights trigger biomass accumulation and production of antioxidant secondary metabolites in adventitious root cultures of Stevia rebaudiana (Bert.). Comptes Rendus Boil. 2018, 341, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Nakonechnaya, O.V.; Gafitskaya, I.V.; Burkovskaya, E.V.; Khrolenko, Y.A.; Grishchenko, O.V.; Zhuravlev, Y.N.; Subbotin, E.P.O.; Kulchin, Y.N. Effect of light intensity on the morphogenesis of Stevia rebaudiana under in vitro conditions. Russ. J. Plant Physiol. 2019, 66, 656–663. [Google Scholar] [CrossRef]
- Melviana, A.C.; Esyanti, R.R.; Setyobudi, R.H.; Mel, M.; Adinurani, P.G.; Burlakovs, J. Gene expression related to steviol glycoside synthesis produced in Stevia rebaudiana (Bert.) shoot culture induced with high far-red LED light in TIS RITA® bioreactor system. Sarhad J. Agric. 2021, 37, 1–8. [Google Scholar] [CrossRef]
- Kalashnikova, E.A.; Kirakosyan, R.N.; Trukhachev, V.I.; Pankova, M.G.; Sumin, A.V. Features of growth and inulin content in callus cultures Cichorium intybus L. in vitro. Russ. J. Plant Physiol. 2023, 70, 78. [Google Scholar] [CrossRef]
- Yoneda, Y.; Nakashima, H.; Miyasaka, J.; Ohdoi, K.; Shimizu, H. Impact of blue, red, and far-red light treatments on gene expression and steviol glycoside accumulation in Stevia rebaudiana. Phytochemistry 2017, 137, 57–65. [Google Scholar] [CrossRef]
- Salehi, M.; Karimzadeh, G.; Naghavi, M.R. Synergistic effect of coronatine and sorbitol on artemisinin production in cell suspension culture of Artemisia annua L. cv. Anamed. Plant Cell Tissue Organ Cult. 2019, 137, 587–597. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: A review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Sharma, S.; Saxena, S. Biomass yield and steviol glycoside production in callus and suspension culture of Stevia rebaudiana treated with proline and polyethylene glycol. Appl. Biochem. Biotechnol. 2015, 176, 863–874. [Google Scholar] [CrossRef]
- Kumar, P.; Subiramani, S.; Govindarajan, S.; Sadasivam, V.; Manickam, V.; Mogilicherla, K.; Thiruppathi, S.; Narayanasamy, J. Evaluation of different carbon sources for high frequency callus culture with reduced phenolic secretion in cotton (Gossypium hirsutum L.) cv. SVPR-2 G. Biotechnol. Rep. 2015, 7, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Petrova, M.; Zayova, E.; Dincheva, I.; Badjakov, I.; Vlahova, M. Influence of carbon sources on growth and GC-MS based metabolite profiling of Arnica montana L. hairy roots. Turk. J. Biol. 2015, 39, 469–478. [Google Scholar] [CrossRef]
- Sharaf, S.A.; Shibli, R.A.; Kasrawi, M.A.; Baghdadi, S.H. Slow-growth preservation of wild shih (Artemisia herba-alba Asso.) microshoots from Jordan. J. Food Agric. Environ. 2012, 10, 1359–1364. [Google Scholar]
- Petrova, M.; Zayova, E.; Geneva, M.; Dimitrova, L.; Vitkova, A.; Stanilova, M. Multiplication and conservation of threatened medicinal plant Arnica montana L. by in vitro techniques. Agric. Consp. Sci. 2021, 86, 57–65. [Google Scholar]
- Zayova, E.; Petrova, M.; Nikolova, M.; Dimitrova, L. Effect of medium salt strength on the micropropagation, phenolic content and antioxidant activity of Arnica montana L., threatened plant species. Bio. Bulletin. 2016, 2, 6–13. [Google Scholar]
- Gonzalez-Dugo, V.; Durand, J.L.; Gastal, F.; Bariac, T.; Poincheval, J. Restricted root-to-shoot translocation and decreased sink size are responsible for limited nitrogen uptake in three grass species under water deficit. Environ. Exp. Bot. 2012, 75, 258–267. [Google Scholar] [CrossRef]
- Srivastava, S.; Srivastava, M. Morphological changes and antioxidant activity of Stevia rebaudiana under water stress. Am. J. Plant Sci. 2014, 5, 3417. [Google Scholar] [CrossRef]
- Maurino, V.G.; Flügge, U.I. Experimental systems to assess the effects of reactive oxygen species in plant tissues. Plant Signal. Behav. 2008, 3, 923–928. [Google Scholar] [CrossRef] [PubMed]
- Lahijanian, S.; Eskandari, M.; Akhbarfar, G.; Azizi, I.; Afazel, M.; Ghobadi, C. Morphological, physiological and antioxidant response of Stevia rebaudiana under in vitro agar induced drought stress. J. Agric. Food Res. 2023, 11, 100495. [Google Scholar] [CrossRef]
- Zahir, A.; Abbasi, B.H.; Adil, M.; Anjum, S.; Zia, M. Synergistic effects of drought stress and photoperiods on phenology and secondary metabolism of Silybum marianum. Appl. Biochem. Biotechnol. 2014, 174, 693–707. [Google Scholar] [CrossRef] [PubMed]
- ElSayed, A.I.; El-hamahmy, M.A.M.; Rafudeen, M.S.; Mohamed, A.H.; Omar, A.A. The Impact of drought stress on antioxidant responses and accumulation of flavonolignans in milk thistle (Silybum marianum (L.) Gaertn). Plants 2019, 8, 611. [Google Scholar] [CrossRef] [PubMed]
- De Caroli, M.; Rampino, P.; Curci, L.M.; Pecatelli, G.; Carrozzo, S.; Piro, G. CiXTH29 and CiLEA4 Role in Water Stress Tolerance in Cichorium intybus Varieties. Biology 2023, 12, 444. [Google Scholar] [CrossRef] [PubMed]
- Vandoorne, B.; Mathieu, A.S.; Van den Ende, W.; Vergauwen, R.; Périlleux, C.; Javaux, M.; Lutts, S. Water stress drastically reduces root growth and inulin yield in Cichorium intybus (var. sativum) independently of photosynthesis. J. Exp. Bot. 2012, 63, 4359–4373. [Google Scholar] [CrossRef] [PubMed]
- Soni, P.; Abdin, M.Z. Water deficit-induced oxidative stress affects artemisinin content and expression of proline metabolic genes in Artemisia annua L. FEBS Open Bio. 2017, 7, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Motos, J.R.; Diaz-Vivancos, P.; Alvarez, S.; Fernandez-Garcia, N.; Sanchez-Blanco, M.J.; Hernandez, J.A. Physiological and biochemical mechanisms of the ornamental Eugenia myrtifolia L. plants for coping with NaCl stress and recovery. Planta 2015, 242, 829–846. [Google Scholar] [CrossRef]
- Wu, H. Plant salt tolerance and Na+ sensing and transport. Crop J. 2018, 6, 215–225. [Google Scholar] [CrossRef]
- Hernández, J.A.; Jiménez, A.; Mullineaux, P.M.; Sevilla, F. Tolerance of pea (Pisum sativum L.) to long-term salt stress is associated with induction of antioxidant defenses. Plant Cell Environ. 2000, 23, 853–862. [Google Scholar] [CrossRef]
- Winkel-Shirley, B. Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant Biol. 2002, 5, 218–223. [Google Scholar] [CrossRef]
- Selmar, D. Potential of salt and drought stress to increase pharmaceutical significant secondary compounds in plants. Landbauforsch. Volkenrode 2008, 58, 139. [Google Scholar]
- Haghighi, Z.; Karimi, N.; Modarresi, M.; Mollayi, S. Enhancement of compatible solute and secondary metabolites production in Plantago ovata Forsk. by salinity stress. J. Med. Plants Res. 2012, 6, 3495–3500. [Google Scholar]
- Lucho, S.R.; do Amaral, M.N.; Auler, P.A.; Bianchi, V.J.; Ferrer, M.A.; Calderón, A.A.; Braga, E.J.B. Salt stress-induced changes in in vitro cultured Stevia rebaudiana Bertoni: Effect on metabolite contents, antioxidant capacity and expression of steviol glycosides-related biosynthetic genes. J. Plant Growth Regul. 2019, 38, 1341–1353. [Google Scholar] [CrossRef]
- Javed, R.; Gürel, E. Salt stress by NaCl alters the physiology and biochemistry of tissue culture-grown Stevia rebaudiana Bertoni. Turk. J. Agric. For. 2019, 43, 11–20. [Google Scholar] [CrossRef]
- Azzam, C.R.; Al-Taweel, S.K.; Abdel-Aziz, R.M.; Rabea, K.M.; Abou-Sreea, A.I.B.; Rady, M.M.; Ali, E.F. Salinity effects on gene expression, morphological, and physio-biochemical responses of Stevia rebaudiana Bertoni in vitro. Plants 2021, 10, 820. [Google Scholar] [CrossRef] [PubMed]
- Ben Rejeb, F.; Chograni, H.; Ben Romdhane, M.; Riahi, L. Effects of two abiotic elicitors on secondary metabolites accumulation and bioactivity in tree wormwood in vitro shoot cultures. Nat. Resour. Human Health 2023, 3, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Zanella, L.; Gismondi, A.; Di Marco, G.; Braglia, R.; Scuderi, F.; Redi, E.L.; Galgani, A.; Canini, A. Induction of antioxidant metabolites in Moringa oleifera callus by abiotic stresses. J. Nat. Prod. 2019, 82, 2379–2386. [Google Scholar] [CrossRef]
- Anusha, T.S.; Joseph, M.V.; Elyas, K.K. Callus induction and elicitation of total phenolics in callus cell suspension culture of Celastrus paniculatus–willd, an endangered medicinal plant in India. Pharmacogn. J. 2016, 8, 471–475. [Google Scholar]
- Gundlach, H.; Müller, M.J.; Kutchan, T.M.; Zenk, M.H. Jasmonic acid is a signal transducer in elicitor-induced plant cell cultures. Proc. Natl. Acad. Sci. USA 1992, 89, 2389–2393. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Mostafa, S.; Zeng, W.; Jin, B. Function and mechanism of jasmonic acid in plant responses to abiotic and biotic stresses. Int. J. Mol. Sci. 2021, 22, 8568. [Google Scholar] [CrossRef] [PubMed]
- Giri, C.C.; Zaheer, M. Chemical elicitors versus secondary metabolite production in vitro using plant cell, tissue and organ cultures: Recent trends and a sky eye view appraisal. Plant Cell Tiss. Organ Cult. 2016, 126, 1–18. [Google Scholar] [CrossRef]
- Ho, T.-T.; Murthy, H.N.; Park, S.-Y. Methyl jasmonate induced oxidative stress and accumulation of secondary metabolites in plant cell and organ cultures. Int. J. Mol. Sci. 2020, 21, 716. [Google Scholar] [CrossRef] [PubMed]
- Pauwels, L.; Morreel, K.; De Witte, E.; Lammertyn, F.; Van Montagu, M.; Boerjan, W.; Inze, D.; Goossens, A. Mapping methyl jasmonate-mediated transcriptional reprogramming of metabolism and cell cycle progression in cultured Arabidopsis cells. Proc. Natl. Acad. Sci. USA 2008, 105, 1380–1385. [Google Scholar] [CrossRef] [PubMed]
- Baldi, A.; Dixit, V.K. Yield enhancement strategies for artemisinin production by suspension cultures of Artemisia annua. Explor. Horiz. Biotechnol. Glob. Ventur. 2008, 99, 4609–4614. [Google Scholar] [CrossRef] [PubMed]
- Al-Khayri, J.M.; Sudheer, W.N.; Lakshmaiah, V.V.; Mukherjee, E.; Nizam, A.; Thiruvengadam, M.; Nagella, P.; Alessa, F.M.; Al-Mssallem, M.Q.; Rezk, A.A.; et al. Biotechnological approaches for production of artemisinin, an anti-malarial drug from Artemisia annua L. Molecules 2022, 27, 3040. [Google Scholar] [CrossRef] [PubMed]
- Pu, G.B.; Ma, D.M.; Chen, J.L.; Ma, L.Q.; Wang, H.; Li, G.F.; Ye, H.C.; Liu, B.Y. Salicylic acid activates artemisinin biosynthesis in Artemisia annua L. Plant Cell Rep. 2009, 28, 1127–1135. [Google Scholar] [CrossRef]
- Caretto, S.; Quarta, A.; Durante, M.; Nisi, R.; De Paolis, A.; Blando, F.; Mita, G. Methyl jasmonate and miconazole differently affect artemisinin production and gene expression in Artemisia annua suspension cultures. Plant Biol. 2011, 13, 51–58. [Google Scholar] [CrossRef]
- Patra, N.; Srivastava, A.K.; Sharma, S. Study of various factors for enhancement of artemisinin in Artemisia annua hairy roots. Int. J. Chem. Eng. Appl. 2013, 4, 157–160. [Google Scholar] [CrossRef]
- Durante, M.; Caretto, S.; Quarta, A.; De Paolis, A.; Nisi, R.; Mita, G. b-Cyclodextrins enhance artemisinin production in Artemisia annua suspension cell cultures. Appl. Microbiol. Biotechnol. 2011, 90, 1905–1913. [Google Scholar] [CrossRef]
- Putalun, W.; Luealon, W.; De-Eknamkul, W.; Tanaka, H.; Shoyama, Y. Improvement of artemisinin production by chitosan in hairy root cultures of Artemisia annua L. Biotechnol. Lett. 2007, 29, 1143–1146. [Google Scholar] [CrossRef] [PubMed]
- Ahlawat, S.; Saxena, P.; Alam, P.; Wajid, S.; Abdin, M.Z. Modulation of artemisinin biosynthesis by elicitors, inhibitor, and precursor in hairy root cultures of Artemisia annua L. J. Plant Interact. 2014, 9, 811–824. [Google Scholar] [CrossRef]
- Sánchez-Sampedro, M.A.; Fernández-Tárrago, J.; Corchete, P. Yeast extract and methyl jasmonate-induced silymarin production in cell cultures of Silybum marianum (L.) Gaertn. J. Biotechnol. 2005, 119, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Firouzi, A.; Mohammadi, S.A.; Khosrowchahli, M.; Movafeghi, A.; Hasanloo, T. Enhancement of silymarin production in cell culture of Silybum marianum (L) Gaertn by elicitation and precursor feeding. J. Herbs Spices Med. Plants 2013, 19, 262–274. [Google Scholar] [CrossRef]
- Gabr, A.M.M.; Ghareeb, H.; El Shabrawi, H.M.; Smetanska, I.; Bekheet, S.A. Enhancement of silymarin and phenolic compound accumulation in tissue culture of Milk thistle using elicitor feeding and hairy root cultures. J. Genet. Eng. Biotechnol. 2016, 14, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Corchete, P.; Bru, R. Proteome alterations monitored by DIGE analysis in Silybum marianum cell cultures elicited with methyl jasmonate and methyl B cyclodextrin. J. Proteom. 2013, 85, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Ravazzolo, L.; Ruperti, B.; Frigo, M.; Bertaiola, O.; Pressi, G.; Malagoli, M.; Quaggiotti, S. C3H expression is crucial for methyl jasmonate induction of chicoric acid production by Echinacea purpurea (L.) Moench cell suspension cultures. Int. J. Mol. Sci. 2022, 23, 11179. [Google Scholar] [CrossRef]
- Demirci, T.; Özmen, S.; Yılmaz, E.G.; Aşcı, Ö.A.; Baydar, N.G. The influence of methyl jasmonate on growth and caffeic acid derivative contents of in vitro shoot and roots in echinacea (Echinacea purpurea). Indian J. Pharm. Educ. Res. 2017, 51, 513–517. [Google Scholar] [CrossRef]
- Cui, H.Y.; Baque, M.A.; Lee, E.J.; Paek, K.Y. Scale-up of adventitious root cultures of Echinacea angustifolia in a pilot-scale bioreactor for the production of biomass and caffeic acid derivatives. Plant. Biotechnol. Rep. 2013, 7, 297–308. [Google Scholar] [CrossRef]
- Demirci, T. Determination of secondary metabolite production efficiency in Echinacea purpurea callus, shoot, and root in vitro cultures with methyl jasmonate applications. Acta Physiol. Plant 2022, 44, 128. [Google Scholar] [CrossRef]
- An, D.; Wu, C.H.; Wang, M.; Wang, M.; Chang, G.N.; Chang, X.J.; Lian, M.L. Methyl jasmonate elicits enhancement of bioactive compound synthesis in adventitious root co-culture of Echinacea purpurea and Echinacea pallida. In Vitro Cell. Dev. Biol. Plant 2022, 58, 181–187. [Google Scholar] [CrossRef]
- Malarz, J.; Stojakowska, A.; Kisiel, W. Effect of methyl jasmonate and salicylic acid on sesquiterpene lactone accumulation in hairy roots of Cichorium intybus. Acta Physiol. Plant. 2007, 29, 127–132. [Google Scholar] [CrossRef]
- Bernard, G.; Alves Dos Santos, H.; Etienne, A.; Samaillie, J.; Neut, C.; Sahpaz, S.; Hilbert, J.-L.; Gagneul, D.; Jullian, N.; Tahrioui, A.; et al. MeJA elicitation of chicory hairy roots promotes efficient increase of 3,5-diCQA accumulation, a potent antioxidant and antibacterial molecule. Antibiotics 2020, 9, 659. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Abbasi, B.H.; Ali, G.S. Elicitation of antioxidant secondary metabolites with jasmonates and gibberellic acid in cell suspension cultures of Artemisia absinthium L. Plant. Cell Tissue Organ. Cult. 2015, 120, 1099–1106. [Google Scholar] [CrossRef]
- Taj, F.; Khan, M.A.; Ali, H.; Khan, R.S. Improved production of industrially important essential oils through elicitation in the adventitious roots of Artemisia amygdalina. Plants 2019, 8, 430. [Google Scholar] [CrossRef] [PubMed]
- Pourianezhad, F.; Rahnama, H.; Mousavi, A.; Khosrowshahli, M.; Mafakheri, S. Effects of combined elicitors on parthenolide production and expression of parthenolide synthase (TpPTS) in Tanacetum parthenium hairy root culture. Plant Biotechnol. Rep. 2019, 13, 211–218. [Google Scholar] [CrossRef]
- Pourianezhad, F.; Rahnama, H.; Mousavi, A.; Khosrowshahli, M.; Mafakheri, S. Parthenolide production in cell suspension culture of feverfew. Bioresour. Bioprocess. 2019, 6, 23. [Google Scholar] [CrossRef]
- Lucho, S.R.; do Amaral, M.N.; Milech, C.; Ferrer, M.Á.; Calderón, A.A.; Bianchi, V.J.; Braga, E.J. Elicitor-induced transcriptional changes of genes of the steviol glycoside biosynthesis pathway in Stevia rebaudiana Bertoni. J. Plant Growth Regul. 2018, 37, 971–985. [Google Scholar] [CrossRef]
- Xu, L.L.; Fan, Z.Y.; Dong, Y.J.; Kong, J.; Bai, X.Y. Effects of exogenous salicylic acid and nitric oxide on physiological characteristics of two peanut cultivars under cadmium stress. Biol. Plant 2015, 59, 171–182. [Google Scholar] [CrossRef]
- Moharramnejad, S.; Azam, A.T.; Panahandeh, J.; Dehghanian, Z.; Ashraf, M. Effect of methyl jasmonate and salicylic acid on in vitro growth, stevioside production, and oxidative defense system in Stevia rebaudiana. Sugar Tech 2019, 21, 1031–1038. [Google Scholar] [CrossRef]
- Golkar, P.; Moradi, M.; Garousi, G.A. Elicitation of stevia glycosides using salicylic acid and silver nanoparticles under callus culture. Sugar Tech 2019, 21, 569–577. [Google Scholar] [CrossRef]
- Aftab, T.; Masroor, M.; Khan, A.Z.; Idrees, M.; Naeem, M.; Moinuddin. Salicylic acid acts as potent enhancer of growth, photosynthesis and artemisinin production in Artemisia annua L. J. Crop Sci. Biotechnol. 2010, 13, 183–188. [Google Scholar] [CrossRef]
- Abbaspour, J.; Ehsanpour, A.A. Physiological targets of salicylic acid on Artemisia aucheri BOISS as a medicinal and aromatic plant grown under in vitro drought stress. Bot. Stud. 2016, 57, 39. [Google Scholar] [CrossRef] [PubMed]
- Al-Amery, L.K.J.; Abdul-Qader, Z.M.; Husni, H.S. Improving propagation of Echinacea purpurea and its content of some active compounds by using Tyrosine and Salicylic acid in vitro. Baghdad Sci. J. 2023, 20, 919–927. [Google Scholar] [CrossRef]
- Kim, D.H.; Gopal, J.; Sivanesan, I. Nanomaterials in plant tissue culture: The disclosed and undisclosed. RSC Adv. 2017, 7, 36492–36505. [Google Scholar] [CrossRef]
- Javed, R.; Usman, M.; Yucesan, B.; Zia, M.; Gurel, E. Effect of zinc oxide (ZnO) nanoparticles on physiology and steviol glycosides production in micropropagated shoots of Stevia rebaudiana Bertoni. Plant Physiol. Biochem. 2017, 110, 94–99. [Google Scholar] [CrossRef]
- Javed, R.; Mohamed, A.; Yücesan, B.; Gürel, E.; Kausar, R.; Zia, M. CuO nanoparticles significantly influence in vitro culture, steviol glycosides, and antioxidant activities of Stevia rebaudiana Bertoni. Plant Cell Tissue Organ Cult. 2017, 131, 611–620. [Google Scholar] [CrossRef]
- Ahmad, M.A.; Javed, R.; Adeel, M.; Rizwan, M.; Ao, Q.; Yang, Y. Engineered ZnO and CuO nanoparticles ameliorate morphological and biochemical response in tissue culture regenerants of candyleaf (Stevia rebaudiana). Molecules 2020, 25, 1356. [Google Scholar] [CrossRef]
- Ghazal, B.; Saif, S.; Farid, K.; Khan, A.; Rehman, S.; Reshma, A.; Fazal, H.; Ali, M.; Ahmad, A.; Rahman, L.; et al. Stimulation of secondary metabolites by copper and gold nanoparticles in submerge adventitious root cultures of Stevia rebaudiana (Bert.). IET Nanobiotechnol. 2018, 12, 569–573. [Google Scholar] [CrossRef]
- Khan, M.A.; Ali, A.; Mohammad, S.; Ali, H.; Khan, T.; Mashwani, Z.; Jan, A.; Ahmad, P. Iron nano modulated growth and biosynthesis of steviol glycosides in Stevia rebaudiana. Plant Cell Tissue Organ Cult. 2020, 143, 121–130. [Google Scholar] [CrossRef]
- Ramezani, M.; Asghari, S.; Gerami, M.; Ramezani, F.; Abdolmaleki, M.K. Effect of silver nanoparticle treatment on the expression of key genes involved in glycosides biosynthetic pathway in Stevia rebaudiana B. plant. Sugar Tech 2020, 22, 518–527. [Google Scholar] [CrossRef]
- Castro-González, C.G.; Sánchez-Segura, L.; Gómez-Merino, F.C.; Bello-Bello, J.J. Exposure of stevia (Stevia rebaudiana B.) to silver nanoparticles in vitro: Transport and accumulation. Sci. Rep. 2019, 9, 10372. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Gupta, S.; Jain, R.; Kothari, S.L.; Kachhwaha, S. SiO2 nanoparticles as elicitor for increased rebaudioside-A in Stevia rebaudiana micropropagated in solid and liquid cultures: A comparative study. Plant Cell Tissue Organ Cult. (PCTOC) 2023, 155, 541–552. [Google Scholar] [CrossRef]
- Desai, C.V.; Desai, H.B.; Suthar, K.P.; Singh, D.; Patel, R.M.; Taslim, A. Phytotoxicity of zinc-nanoparticles and its influence on stevioside production in Stevia rebaudiana Bertoni. Appl. Biol. Res. 2015, 17, 1–7. [Google Scholar] [CrossRef]
- Desai Heta, B.; Desai Charmi, V.; Desai Charmi, P.; Singh, D.; Suthar, H.G. Effect of magnesium nanoparticles on physiology and stevioside in Stevia rebaudiana Bertoni. Eur. J. Biomed. Pharm. Sci. 2017, 4, 642–646. [Google Scholar]
- Zhang, B.; Zheng, L.P.; Yi Li, W.; Wen Wang, J. Stimulation of artemisinin production in Artemisia annua hairy roots by Ag-SiO2 core-shell nanoparticles. Curr. Nanosci. 2013, 9, 363–370. [Google Scholar] [CrossRef]
- Bami, S.S.; Khavari-Nejad, R.A.; Ahadi, A.M.; Rezayatmand, Z. TiO2 nanoparticles and salinity stress in relation to artemisinin production and ADS and DBR2 expression in Artemisia absinthium L. Braz. J. Biol. 2021, 82, e237214. [Google Scholar] [CrossRef]
- Ghassemi, B.; Nayeri, F.D.; Hosseini, R. The effects of chitosan nanoparticles on genes expression of artemisinin synthase in suspension culture of Artemisia annua L.: A comparative study. Int. J. Adv. Biol. Biomed. Res. 2021, 9, 190–203. [Google Scholar]
- Ghassemi, B.; Hosseini, R.; Dehghan Nayeri, F. Effects of cobalt nano particles on artemisinin production and gene expression in Artemisia annua. Turk. J. Bot. 2015, 39, 769–777. [Google Scholar] [CrossRef]
- Sichanova, M.; Geneva, M.; Petrova, M.; Miladinova-Georgieva, K.; Kirova, E.; Nedev, T.; Tsekova, D.; Iwanov, I.; Dochev, K.; Ivanova, V.; et al. Improvement of Stevia rebaudiana Bertoni in vitro propagation and steviol glycoside content using aminoacid silver nanofibers. Plants 2022, 11, 2468. [Google Scholar] [CrossRef]
- Sichanova, M.; Geneva, M.; Petrova, M.; Miladinova-Georgieva, K.; Kirova, E.; Nedev, T.; Tsekova, D.; Ivanova, V.; Trendafilova, A. Influence of the abiotic elicitors Ag salts of aspartic acid derivatives, self-organized in nanofibers with monomeric and dimeric molecular structures, on the antioxidant activity and stevioside content in micropropagated Stevia rebaudiana Bert. Plants 2023, 12, 3574. [Google Scholar] [CrossRef] [PubMed]
- Mosavat, N.; Golkar, P.; Yousefifard, M.; Javed, R. Modulation of callus growth and secondary metabolites in different Thymus species and Zataria multiflora micropropagated under ZnO nanoparticles stress. Biotech. Appl. Biochem. 2019, 66, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Jianwen, W.; Zhonghao, X.; Renxiang, T. Elicitation on artemisinin biosynthesis in Artemisia annua hairy roots by the oligosaccharide extract from the endophytic Colletotrichum sp. B501. Acta Bot. Sin. 2002, 44, 1233–1238. [Google Scholar]
- Wang, J.-W.; Xiang, R. Stimulation of artemisinin production in Artemisia annua hairy roots by the elicitor from the endophytic Colleotrichum sp. Biotechnol. Lett. 2001, 23, 857–860. [Google Scholar] [CrossRef]
- Wang, J.W.; Zheng, L.P.; Zhang, B.; Zou, T. Stimulation of artemisinin synthesis by combined cerebroside and nitric oxide elicitation in Artemisia annua hairy roots. Appl. Microbiol. Biotechnol. 2009, 85, 285–292. [Google Scholar] [CrossRef]
- Zheng, L.-P.; Guo, Y.-T.; Wang, J.-W.; Tan, R.-X. Nitric Oxide potentiates Oligosaccharide-induced artemisinin production in Artemisia annua hairy roots. J. Integr. Plant Biol. 2008, 50, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.P.; Tian, H.; Yuan, Y.F.; Wang, J.W. The influence of endophytic Penicillium oxalicum B4 on growth and artemisinin biosynthesis of in vitro propagated plantlets of Artemisia annua L. Plant Growth Regul. 2016, 80, 93–102. [Google Scholar] [CrossRef]
- Sharma, G.; Agrawal, V. Marked enhancement in the artemisinin content and biomass productivity in Artemisia annua L. shoots co-cultivated with Piriformospora indica. World J. Microbiol. Biotechnol. 2013, 29, 1133–1138. [Google Scholar] [CrossRef]
- Bayraktar, M.; Naziri, E.; Akgun, I.H.; Karabey, F.; Ilhan, E.; Akyol, B.; Gurel, A. Elicitor induced stevioside production, in vitro shoot growth, and biomass accumulation in micropropagated Stevia rebaudiana. Plant Cell Tissue Org. Cult. 2016, 127, 289–300. [Google Scholar] [CrossRef]
- Elshahawy, O.A.; Zeawail, M.E.-F.; Hamza, M.A.; Elateeq, A.A.; Omar, M.A. Improving the production of total phenolics and flavonoids and the antioxidant capacity of Echinacea purpurea callus through biotic elicitation. Egypt. J. Chem. 2022, 65, 137–149. [Google Scholar] [CrossRef]
- Shah, M.; Jan, H.; Drouet, S.; Tungmunnithum, D.; Shirazi, J.H.; Hano, C.; Abbasi, B.H. Chitosan elicitation impacts flavonolignan biosynthesis in Silybum marianum (L.) Gaertn cell suspension and enhances antioxidant and anti-inflammatory activities of cell extracts. Molecules 2021, 26, 791. [Google Scholar] [CrossRef] [PubMed]
- Habib, M.S.; Salim, S.A. Effect of yeast extract and gibberellic acid on the Artemisia herba alba callus antioxidant activity and its contents of pharmaceutical compounds. NeuroQuantology 2022, 20, 7312. [Google Scholar]

| Plant Species | Elicitors | Type of In Vitro Culture | Effects | References |
|---|---|---|---|---|
| Nanoparticles | ||||
| Stevia rebaudiana Bert. (stevia) | ZnO | ShC | Higher steviol glycoside content | [119] |
| Stevia rebaudiana Bert. (stevia) | ZnO and CuO | ShC | Higher rooting, steviol glycoside, TPC, TFC, and DPPH free radical scavenging activity | [121] |
| Stevia rebaudiana Bert. (stevia) | CuO | ShC | Positive effect on shoot organogenesis, steviol glycoside content, and AA | [120] |
| Stevia rebaudiana Bert. (stevia) | AuCu | AdvRC | Positive impact on biomass and secondary metabolite production | [122] |
| Stevia rebaudiana Bert. (stevia) | FeNPs | ShC | Improved growth parameters, higher levels of TPC, TFC and steviol glycosides, and AA | [123] |
| Stevia rebaudiana Bert. (stevia) | Ag | ShC | Positive effect on the shoot production and length, as a microbicidal agent, as an inhibitor in ethylene synthesis, in photosynthetic pigment synthesis, nutrient accumulation, AA, and ROS generation | [125] |
| Stevia rebaudiana Bert. (stevia) | SiO2 | ShC | Enhanced secondary metabolite biosynthesis | [126] |
| Stevia rebaudiana Bert. (stevia) | Zn | ShC | Potential phytotoxicity | [127] |
| Stevia rebaudiana Bert. (stevia) | Mg | ShC | Normal plant physiology and stevioside production | [128] |
| Stevia rebaudiana Bert. (stevia) | Valine derivative nanofibers, carriers of silver atoms (NF-1%Ag and NF-2%Ag) | ShC | The stimulation of plant growth at low concentrations; the highest amount of stevioside at the highest NF-1%Ag concentration | [133] |
| Stevia rebaudiana Bert. (stevia) | L-aspartic acid derivative nanofibers with a dimeric molecular structure, carriers of Ag (NF2-Ag salt) | ShC | Increased soluble sugars and total dicaffeoylquinic (DCQA) acid content, enhanced CQA/DCQA ratio and micropropagation rate | [134] |
| Artemisia annua L. (sweet wormwood) | Ag-SiO2 | HR | Induced oxidative stress (higher H2O2 and MDA content), enhanced catalase activity, and increased artemisinin content | [129] |
| Artemisia annua L. (sweet wormwood) | Co | SuspC | Higher artemisinin yield; an upregulated expression of ADS, CYP, CPR, DBR2, and ALDH genes; a decreased expression of SQS and DBR2 genes | [131,132] |
| Biotic Elicitor | ||||
|---|---|---|---|---|
| Plant Species | Biotic Elicitors | Type of In Vitro Culture | Effects | References |
| Artemisia annua L. (sweet wormwood) | Cell wall’s oligosaccharide from Colletotrichum sp. B501 | HR | Increased artemisinin yield by 68.29% | [136] |
| Artemisia annua L. (sweet wormwood) | Cerebroside from fungal source | HR | Oxidative bursts that released nitric oxide (NO); increased artemisinin yield 2.3 folds | [138] |
| Artemisia annua L. (sweet wormwood) | Oligosaccharide from Fusarium oxysporum mycelium | HR | Increased artemisinin content from 0.7 mg/g DW to 1.3 mg/g DW | [139] |
| Artemisia annua L. (sweet wormwood) | Penicillium oxalicum B4 | In vitro-grown rooted plantlets | Increased artemisinin content by 43.5% | [140] |
| Stevia rebaudiana Bert. (stevia) | Alginate, casein hydrolysate, pectin, yeast extract, and chitosan | ShC | Biomass accumulation is the highest when 100 µM chitosan is used. The in vitro plantlets treated with either 2.0 g/L YE or 0.5 g/L ALG produced the highest amount of stevioside (14.69 and 14.54 mg/g DW, respectively). | [142] |
| Artemisia annua L. (sweet wormwood) | Chitosan (150 mg/L) | HR | Enhanced the accumulation of artemisinin | [94] |
| Echinacea purpurea L. (echinacea) | Fungal elicitors yeast extract, Aspergillus niger and Fusarium oxysporum | CC | Total phenolics is recorded with 4 g/L YE, 1 g/L A. niger or 0.25 g/L F. Oxysporum | [143] |
| Silybum marianum L. (milk thistle) | Chitosan (0.5–50.0 mg/L) | SuspC | Chitosan (5.0 mg/L) improved both biomass production and the accumulation of silymarin | [144] |
| Artemisia alba Turra. (white wormwood) | Yeast extract at a concentration of 100 or 200 mg/L | CC | An enhanced accumulation of santonin and artemisinin | [145] |
| Artemisia annua L. (sweet wormwood) | Yeast extract Chitosan | SuspC | Increased artemisinin accumulation | [88] |
| Artemisia annua L. (sweet wormwood) | Piriformospora indica homogenates and MeJA | HR | An increase in artemisinin accumulation; the effects of Piriformospora indica on artemisinin synthesis, positively correlated with regulatory genes of MVA, MEP, and artemisinin biosynthetic genes hmgr, ads, cyp71av1, aldh1, dxs, dxr, and dbr2 | [95] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrova, M.; Miladinova-Georgieva, K.; Geneva, M. Influence of Abiotic and Biotic Elicitors on Organogenesis, Biomass Accumulation, and Production of Key Secondary Metabolites in Asteraceae Plants. Int. J. Mol. Sci. 2024, 25, 4197. https://doi.org/10.3390/ijms25084197
Petrova M, Miladinova-Georgieva K, Geneva M. Influence of Abiotic and Biotic Elicitors on Organogenesis, Biomass Accumulation, and Production of Key Secondary Metabolites in Asteraceae Plants. International Journal of Molecular Sciences. 2024; 25(8):4197. https://doi.org/10.3390/ijms25084197
Chicago/Turabian StylePetrova, Maria, Kamelia Miladinova-Georgieva, and Maria Geneva. 2024. "Influence of Abiotic and Biotic Elicitors on Organogenesis, Biomass Accumulation, and Production of Key Secondary Metabolites in Asteraceae Plants" International Journal of Molecular Sciences 25, no. 8: 4197. https://doi.org/10.3390/ijms25084197
APA StylePetrova, M., Miladinova-Georgieva, K., & Geneva, M. (2024). Influence of Abiotic and Biotic Elicitors on Organogenesis, Biomass Accumulation, and Production of Key Secondary Metabolites in Asteraceae Plants. International Journal of Molecular Sciences, 25(8), 4197. https://doi.org/10.3390/ijms25084197