
The SARS-CoV-2 Spike Protein Receptor-Binding Domain Expressed in Rice Callus Features a Homogeneous Mix of Complex-Type Glycans

 , , , and
, , , and
Abstract
:1. Introduction
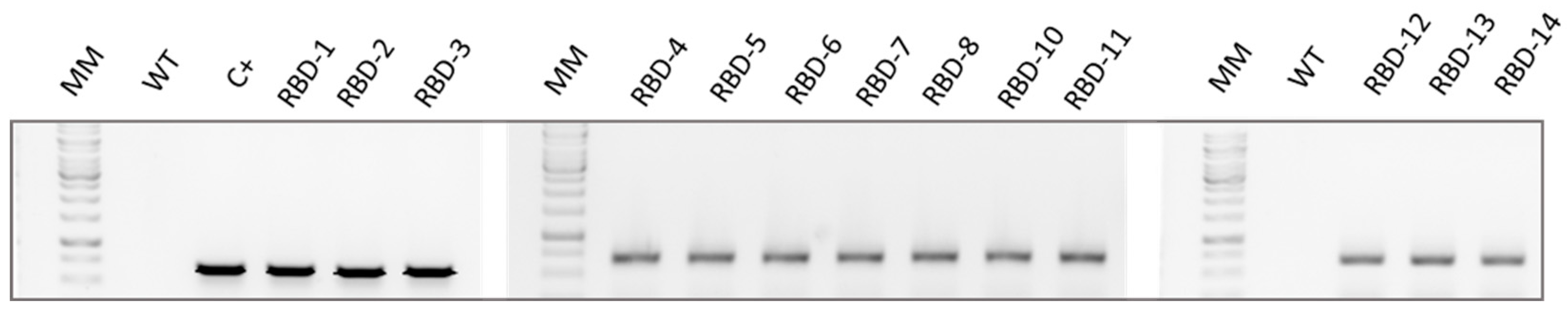
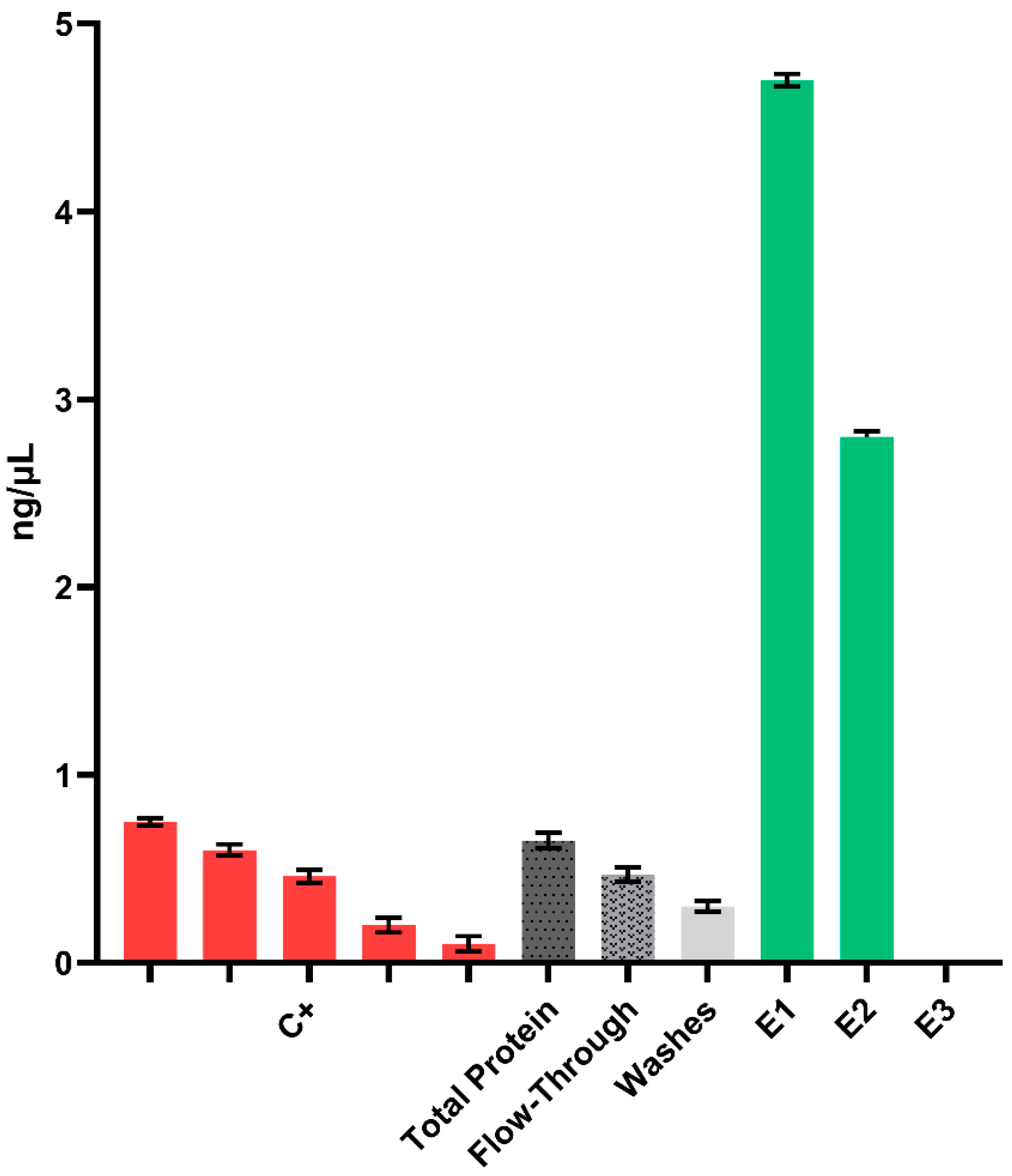
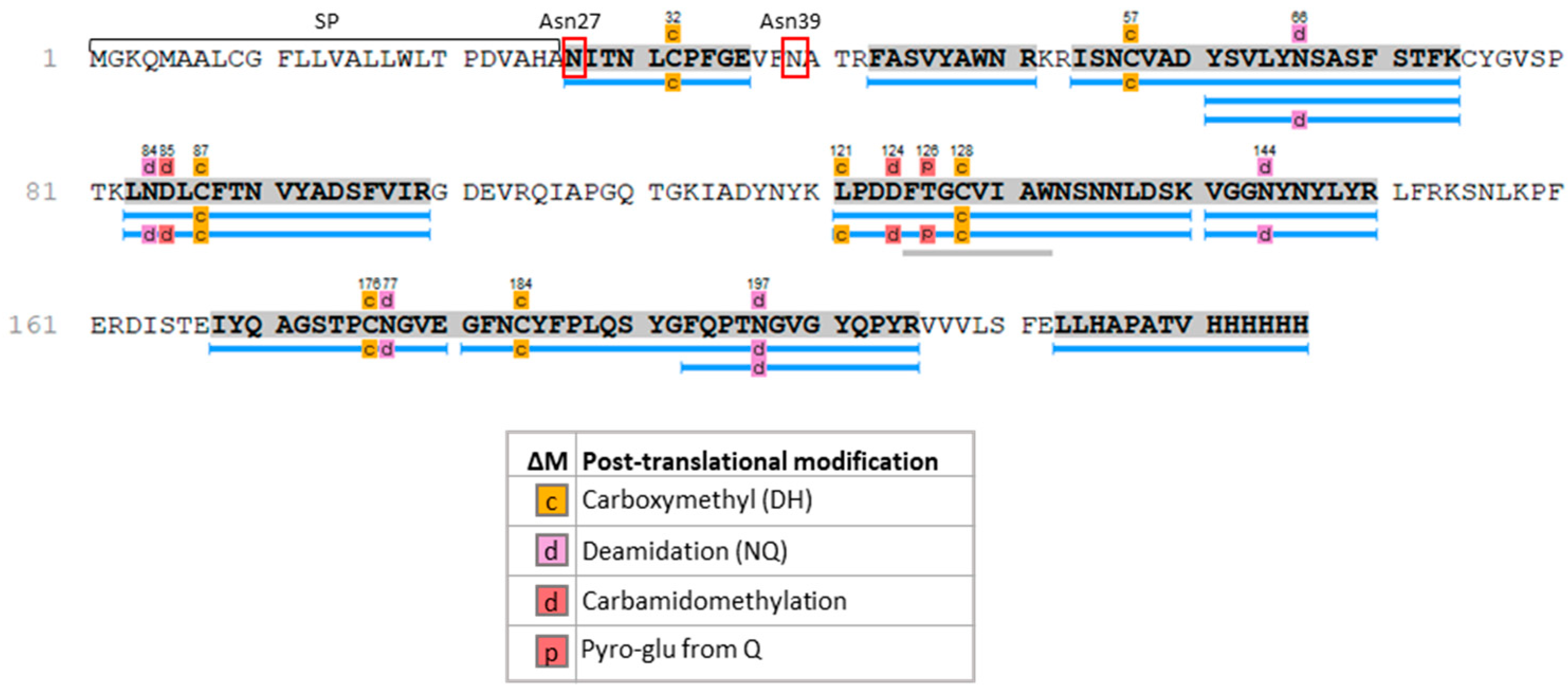
2. Results
3. Discussion
4. Materials and Methods
4.1. Genetic Constructs for Rice Transformation
4.2. Transformation and Regeneration of Transgenic Rice Callus
4.3. DNA Extraction and Analysis
4.4. Protein Extraction
4.5. ELISA
4.6. SDS-PAGE and Western Blot Analysis
4.7. Gel Staining
4.8. Recombinant Protein Purification
4.9. Ammonium Sulfate Precipitation
4.10. Glycosylation Analysis
Endoprotease Digestions
4.11. LC-ESI-MS/MS Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Basta, N.E.; Moodie, E.M.M., on behalf of the VIPER (Vaccines, Infectious disease Prevention, and Epidemiology Research) Group COVID-19 Vaccine Development and Approvals Tracker Team (2020). COVID-19 Vaccine Development and Approvals Tracker. Available online: https://covid19.trackvaccines.org/our-team/ (accessed on 30 August 2022).
- Lupala, C.S.; Kumar, V.; Su, X.D.; Wu, C.; Liu, H. Computational insights into differential interaction of mammalian angiotensin-converting enzyme 2 with the SARS-CoV-2 spike receptor binding domain. Comput. Biol. Med. 2022, 141, 105017. [Google Scholar] [CrossRef]
- Capell, T.; Twyman, R.M.; Armario-Najera, V.; Ma, J.K.-C.; Schillberg, S.; Christou, P. Potential applications of plant biotechnology against SARS-CoV-2. Trends Plant Sci. 2020, 25, 635–643. [Google Scholar] [CrossRef]
- Diego-Martin, B.; González, B.; Vazquez-Vilar, M.; Selma, S.; Mateos-Fernández, R.; Gianoglio, S.; Fernández-del-Carmen, A.; Orzáez, D. Pilot production of SARS-CoV-2 related proteins in plants: A proof of concept for rapid repurposing of indoor farms into biomanufacturing facilities. Front. Plant Sci. 2020, 11, 2101. [Google Scholar] [CrossRef]
- Santoni, M.; Gutierrez-Valdes, N.; Pivotto, D.; Zanichelli, E.; Rosa, A.; Sobrino-Mengual, G.; Balieu, J.; Lerouge, P.; Bardor, M.; Cecchetto, R.; et al. Performance of plant-produced RBDs as SARS-CoV-2 diagnostic reagents: A tale of two plant platforms. Front. Plant Sci. 2024, 14, 1325162. [Google Scholar] [CrossRef]
- Hemmati, F.; Hemmati-Dinarvand, M.; Karimzade, M.; Rutkowska, D.; Eskandari, M.H.; Khanizadeh, S.; Afsharifar, A. Plant-derived VLP: A worthy platform to produce vaccine against SARS-CoV-2. Biotechnol. Lett. 2022, 44, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Kumari, N.; Thakur, N.; Bhatia, S.K.; Saratale, G.D.; Ghodake, G.; Mistry, B.M.; Alavilli, H.; Kishor, D.S.; Du, X.; et al. A comprehensive overview on the production of vaccines in plant-based expression systems and the scope of plant biotechnology to combat against SARS-CoV-2 virus pandemics. Plants 2021, 10, 1213. [Google Scholar] [CrossRef]
- Siriwattananon, K.; Manopwisedjaroen, S.; Shanmugaraj, B.; Rattanapisit, K.; Phumiamorn, S.; Sapsutthipas, S.; Trisiriwanich, S.; Prompetchara, E.; Ketloy, C.; Buranapraditkun, S.; et al. Plant-produced receptor-binding domain of SARS-CoV-2 elicits potent neutralizing responses in mice and non-human primates. Front. Plant Sci. 2021, 12, 682953. [Google Scholar] [CrossRef] [PubMed]
- Hager, K.J.; Pérez-Marc, G.; Gobeil, P.; Diaz, R.S.; Heizer, G.; Llapur, C.; Makarkov, A.I.; Vasconcellos, E.; Pillet, S. Efficacy and safety of a recombinant plant-based adjuvanted COVID-19 vaccine. N. Engl. J. Med. 2022, 386, 2084–2096. [Google Scholar] [CrossRef] [PubMed]
- Ward, B.J.; Gobeil, P.; Séguin, A.; Atkins, J.; Boulay, I.; Charbonneau, P.Y.; Couture, M.; D’Aoust, M.A.; Dhaliwall, J.; Finkle, C.; et al. Phase 1 randomized trial of a plant-derived virus-like particle vaccine for COVID-19. Nat. Med. 2021, 27, 1071–1078. [Google Scholar] [CrossRef]
- Grant, O.C.; Montgomery, D.; Ito, K.; Woods, R.J. Analysis of the SARS-CoV-2 spike protein glycan shield revelas implications for immune recognition. Sci. Rep. 2020, 10, 14991. [Google Scholar] [CrossRef]
- Maharjan, P.M.; Cheon, J.; Jung, J.; Kim, H.; Lee, J.; Song, M.; Jeong, G.U.; Kwon, Y.; Shim, B.; Choe, S. Plant-Expressed Receptor Binding Domain of the SARS-CoV-2 Spike Protein Elicits Humoral Immunity in Mice. Vaccines 2021, 9, 978. [Google Scholar] [CrossRef]
- Butler, M.; Meneses-Acosta, A. Recent advances in technology supporting biopharmaceutical production from mammalian cells. Appl. Microbiol. Biotechnol. 2012, 96, 885–894. [Google Scholar] [CrossRef]
- Schillberg, S.; Raven, N.; Spiegel, H.; Rasche, S.; Buntru, M. Critical analysis of the commercial potential of plants for the production of recombinant proteins. Front. Plant Sci. 2019, 10, 720. [Google Scholar] [CrossRef] [PubMed]
- Donini, M.; Marusic, C. Current state-of-the-art in plant-based antibody production systems. Biotechnol. Lett. 2019, 41, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Saba-Mayoral, A.; Rosa, C.; Sobrino-Mengual, G.; Armario-Najera, V.; Christou, P.; Capell, T. Production of the SARS-CoV-2 receptor-binding domain in stably-transformed rice plants for developing country applications. Plant Biotechnol. J. 2023, 21, 1094–1096. [Google Scholar] [CrossRef] [PubMed]
- Vamvaka, E.; Twyman, R.M.; Murad, A.; Melnik, S.; Teh, A.; Arcalis, E.; Altmann, F.; Stoger, E.; Rech, E.; Ma, J.K.-C.; et al. Rice endosperm produces an underglycosylated and potent form of the HIV-neutralizing monoclonal antibody 2G12. Plant Biotechnol. J. 2015, 14, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.X.; Amin, M.N. Chemical and chemoenzymatic synthesis of glycoproteins for deciphering functions. Chem. Soc. Rev. 2014, 43, 6844–6858. [Google Scholar] [CrossRef] [PubMed]
- Balieu, J.; Jung, J.W.; Chan, P.; Lomonossoff, G.P.; Lerouge, P.; Bardor, M. Investigation of the N-Glycosylation of the SARS-CoV-2 S Protein Contained in VLPs Produced in Nicotiana benthamiana. Molecules 2022, 11, 5119. [Google Scholar] [CrossRef] [PubMed]
- Lerouge, P.; Cabanes-Macheteau, M.; Rayon, C.; Fitchette-Lainé, A.-C.; Gomord, V.; Faye, L. N-glycoprotein biosynthesis in plants: Recent developments and future trends. Plant Mol. Biol. 1998, 38, 31–48. [Google Scholar] [CrossRef]
- Paul, M.J.; Teh, A.Y.H.; Twyman, R.M.; Ma, J.K. Target product selection—Where can Molecular Pharming make the difference? Curr. Pharm. Des. 2013, 19, 5478–5485. [Google Scholar] [CrossRef]
- He, Y.; Ning, T.; Xie, T.; Qiu, Q.; Zhang, L.; Sun, Y.; Jiang, D.; Fu, K.; Yin, F.; Zhang, W.; et al. Large-scale production of functional human serum albumin from transgenic rice seeds. Proc. Natl. Acad. Sci. USA 2011, 108, 19078–19083. [Google Scholar] [CrossRef]
- Zhang, L.; Shi, J.; Jiang, D.; Stupak, J.; Ou, Q.; An, N.; Li, J.; Yang, D. Expression and characterization of recombinant human alpha-antitrypsin in transgenic rice seed. J. Biotechnol. 2012, 164, 300–308. [Google Scholar] [CrossRef]
- Nandi, S.; Yalda, D.; Lu, S.; Nikolov, Z.; Misaki, R.; Fujiyama, K.; Huang, N. Process development and economic evaluation of recombinant human lactoferrin expressed in rice grain. Transgenic Res. 2005, 14, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Fischer, R.; Holland, T.; Sack, M.; Schillberg, S.; Stoger, E.; Twyman, R.M.; Buyel, J.F. Glyco-engineering of plant-based expression systems. Adv. Biochem. Eng./Biotechnol. 2018, 175, 137–166. [Google Scholar] [CrossRef]
- Rademacher, T.; Sack, M.; Arcalis, E.; Stadlmann, J.; Balzer, S.; Altmann, F.; Quendler, H.; Stiegler, G.; Kunert, R.; Fischer, R.; et al. Recombinant antibody 2G12 produced in maize endosperm efficiently neutralizes HIV-1 and contains predominantly single-GlcNAc N-glycans. Plant Biotechnol. J. 2008, 6, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Loos, A.; Van Droogenbroeck, B.; Hillmer, S.; Grass, J.; Pabst, M.; Castilho, A.; Kunert, R.; Liang, M.; Arcalis, E.; Robinson, D.G.; et al. Expression of antibody fragments with a controlled N-glycosylation pattern and induction of endoplasmic reticulum-derived vesicles in seeds of Arabidopsis. Plant Physiol. 2011, 155, 2036–2048. [Google Scholar] [CrossRef]
- Floss, D.M.; Sack, M.; Arcalis, E.; Stadlmann, J.; Quendler, H.; Rademacher, T.; Stoger, E.; Scheller, J.; Fischer, R.; Conrad, U. Influence of elastin-like peptide fusions on the quantity and quality of a tobacco-derived human immunodeficiency virus-neutralizing antibody. Plant Biotechnol. J. 2009, 7, 899–913. [Google Scholar] [CrossRef]
- Rebelo, B.A.; Folgado, A.; Ferreira, A.C.; Abranches, R. Production of the SARS-CoV-2 spike protein and its receptor binding domain in plant cell suspension cultures. Front. Plant Sci. 2022, 13, 995429. [Google Scholar] [CrossRef] [PubMed]
- Castro, R.; Nobre, L.S.; Eleutério, R.P.; Thomaz, M.; Pires, A.; Monteiro, S.M.; Alves, P.M. Production of high-quality SARS-CoV-2 antigens: Impact of bioprocess and storage on glycosylation, biophysical attributes, and ELISA serologic tests performance. Biotechnol. Bioeng. 2021, 118, 2202–2219. [Google Scholar] [CrossRef]
- Shin, Y.J.; König-Beihammer, J.; Vavra, U.; Schwestka, J.; Kienzl, N.F.; Klausberger, M.; Laurent, E.; Grünwald-Gruber, C.; Vierlinger, K.; Hofner, M.; et al. N-Glycosylation of the SARS-CoV-2 Receptor Binding Domain Is Important for Functional Expression in Plants. Front. Plant Sci. 2021, 12, 689104. [Google Scholar] [CrossRef]
- Schwestka, J.; König-Beihammer, J.; Shin, Y.J.; Vavra, U.; Kienzl, N.F.; Grünwald-Gruber, C.; Maresch, D.; Klausberger, M.; Laurent, E.; Stadler, M.; et al. Impact of Specific N-Glycan Modifications on the Use of Plant-Produced SARS-CoV-2 Antigens in Serological Assays. Front. Plant Sci. 2021, 12, 747500. [Google Scholar] [CrossRef]
- Christensen, A.H.; Quail, P.H. Ubiquitin promoter-based vectors for high-level expression of selectable and/or screenable marker genes in monocotyledonous plants. Transgenic Res. 1996, 5, 213–218. [Google Scholar] [CrossRef]
- Van Den Elzen, P.J.; Townsend, J.; Lee, K.Y.; Bedbrook, J.R. A chimaeric hygromycin resistance gene as a selectable marker in plant cells. Plant Mol. Biol. 1985, 5, 299–302. [Google Scholar] [CrossRef]
- Yanisch-Perron, C.; Vieira, J.; Messing, J. Improved M13 phage cloning vectors and host strains: Nucleotide sequences of the M13mp18 and pUC19 vectors. Gene 1985, 33, 103–119. [Google Scholar] [CrossRef]
- Saba-Mayoral, A.; Bassie, L.; Christou, P.; Capell, T. Development of a facile genetic transformation system for the Spanish elite rice paella genotype Bomba. Transgenic Res. 2022, 31, 325–340. [Google Scholar] [CrossRef]
- Creissen, G.P.; Mullineaux, P.M. Cloning and characterisation of glutathione reductase cDNAs and identification of two genes encoding the tobacco enzyme. Planta 1995, 197, 422–425. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Observed Glycopeptide Ions ([M+2H/2]2+) | Observed Peptide Ions ([M+H]+) | Peptide Sequence | N-Glycan Mass | Glycan Structure | % |
|---|---|---|---|---|---|
| 1142.06 | 1274.75 | VFNATRFASVY | 1026.38 |  | 13 |
| 1223.09 | 1274.76 | VFNATRFASVY | 1188.42 |  | 54 |
| 1325.13 | 1274.75 | VFNATRFASVY | 1391.51 |  | 33 |
| Oligo Name | Sequence (5′ → 3′) |
|---|---|
| F-RBD-pAL-BamHI (FW 1) | AAGGGATCCATGGGCAAGCAGATG |
| R-RBD-pAL-HindIII (RV3) | GCGCAAGCTTTCAGTGATG |
| R-His01 (RV1) | ATGATGATGGCGGGCCCTGCGA |
| R-His02 (RV2) | TCAGTGATGATGATGATGATGGCGGGCCCT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sobrino-Mengual, G.; Armario-Nájera, V.; Balieu, J.; Walet-Balieu, M.-L.; Saba-Mayoral, A.; Pelacho, A.M.; Capell, T.; Christou, P.; Bardor, M.; Lerouge, P. The SARS-CoV-2 Spike Protein Receptor-Binding Domain Expressed in Rice Callus Features a Homogeneous Mix of Complex-Type Glycans. Int. J. Mol. Sci. 2024, 25, 4466. https://doi.org/10.3390/ijms25084466
Sobrino-Mengual G, Armario-Nájera V, Balieu J, Walet-Balieu M-L, Saba-Mayoral A, Pelacho AM, Capell T, Christou P, Bardor M, Lerouge P. The SARS-CoV-2 Spike Protein Receptor-Binding Domain Expressed in Rice Callus Features a Homogeneous Mix of Complex-Type Glycans. International Journal of Molecular Sciences. 2024; 25(8):4466. https://doi.org/10.3390/ijms25084466
Chicago/Turabian StyleSobrino-Mengual, Guillermo, Victoria Armario-Nájera, Juliette Balieu, Marie-Laure Walet-Balieu, Andrea Saba-Mayoral, Ana M. Pelacho, Teresa Capell, Paul Christou, Muriel Bardor, and Patrice Lerouge. 2024. "The SARS-CoV-2 Spike Protein Receptor-Binding Domain Expressed in Rice Callus Features a Homogeneous Mix of Complex-Type Glycans" International Journal of Molecular Sciences 25, no. 8: 4466. https://doi.org/10.3390/ijms25084466
APA StyleSobrino-Mengual, G., Armario-Nájera, V., Balieu, J., Walet-Balieu, M. -L., Saba-Mayoral, A., Pelacho, A. M., Capell, T., Christou, P., Bardor, M., & Lerouge, P. (2024). The SARS-CoV-2 Spike Protein Receptor-Binding Domain Expressed in Rice Callus Features a Homogeneous Mix of Complex-Type Glycans. International Journal of Molecular Sciences, 25(8), 4466. https://doi.org/10.3390/ijms25084466