
Revealing the Complete Bispecific Phosphatase Genes (DUSPs) across the Genome and Investigating the Expression Patterns of GH_A11G3500 Resistance against Verticillium wilt
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Identification and Basic Information of Cotton DUSP Phosphatase Family Members
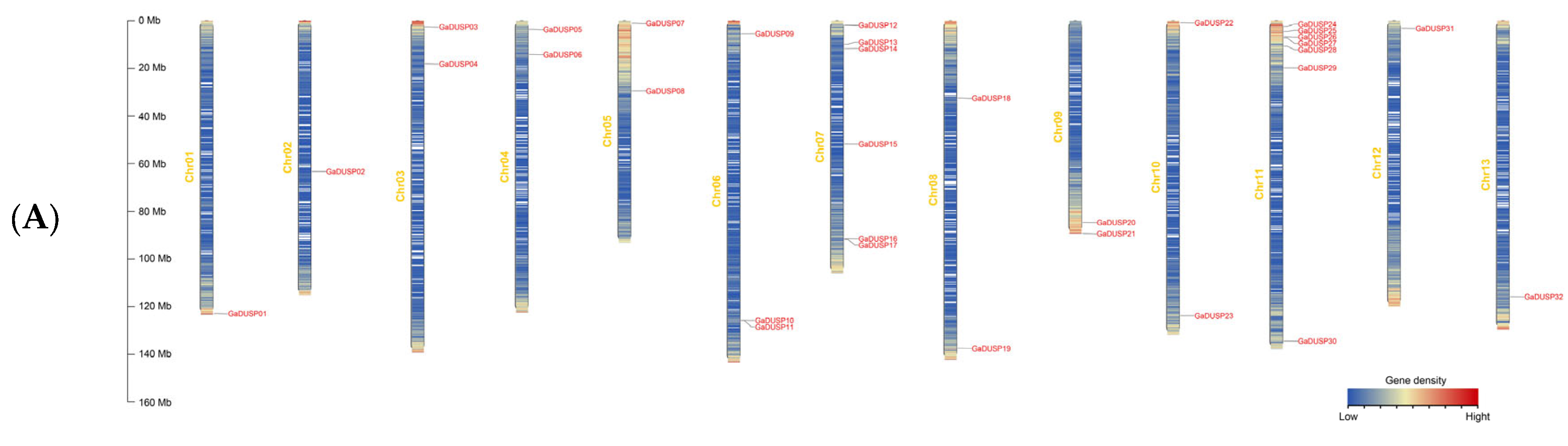
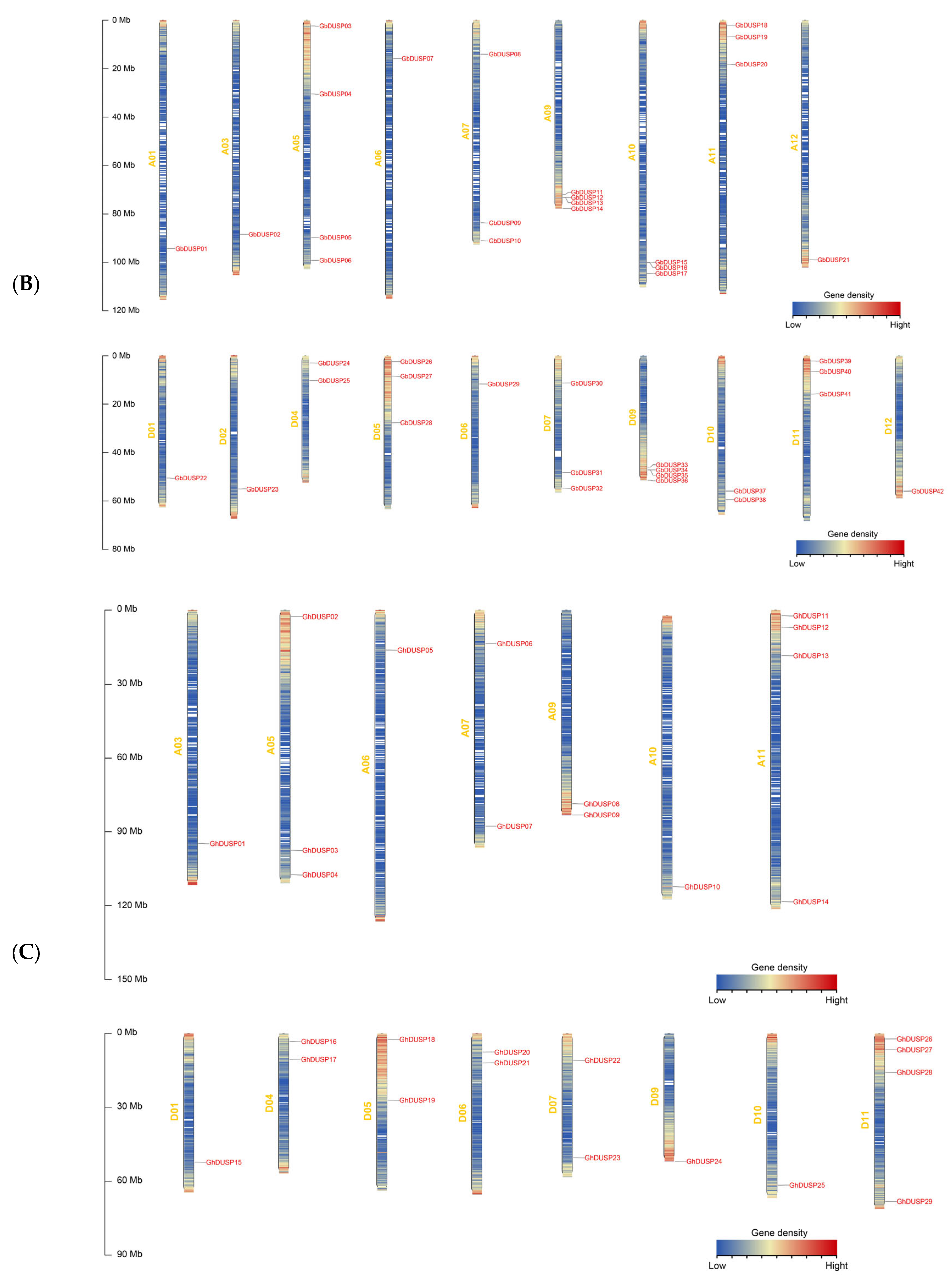
2.2. Distribution of DUSP Family Gene Chromosomes in Cotton
2.3. Phylogenetic Assessment of DUSP Family Genes in Cotton
2.4. Motifs and Gene Structure of DUSP Family Genes in Cotton
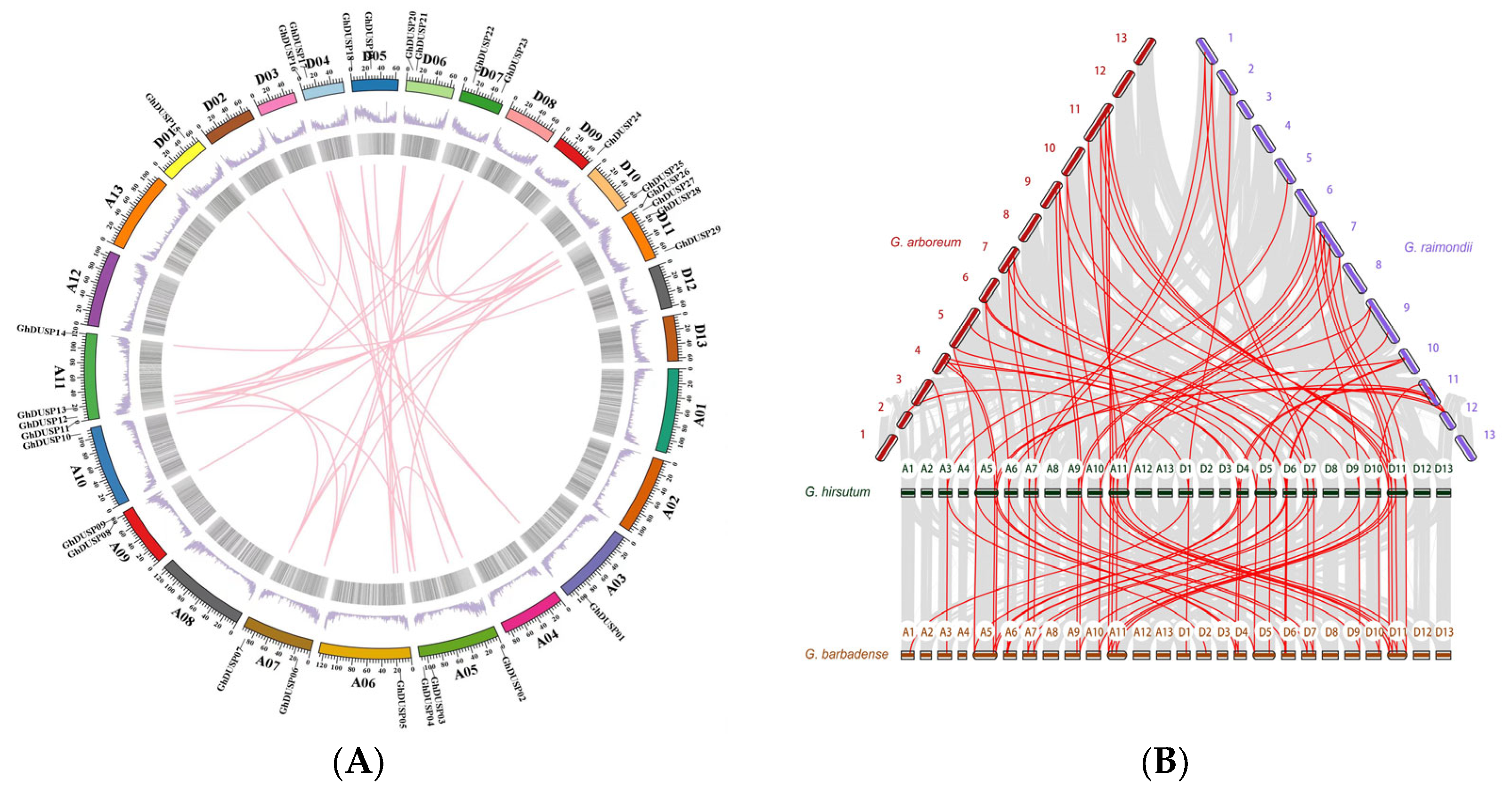
2.5. Exploring the Collinearity among Members of the DUSP Family in Cotton
2.6. Examination of Regulatory Elements in the GhDUSPs
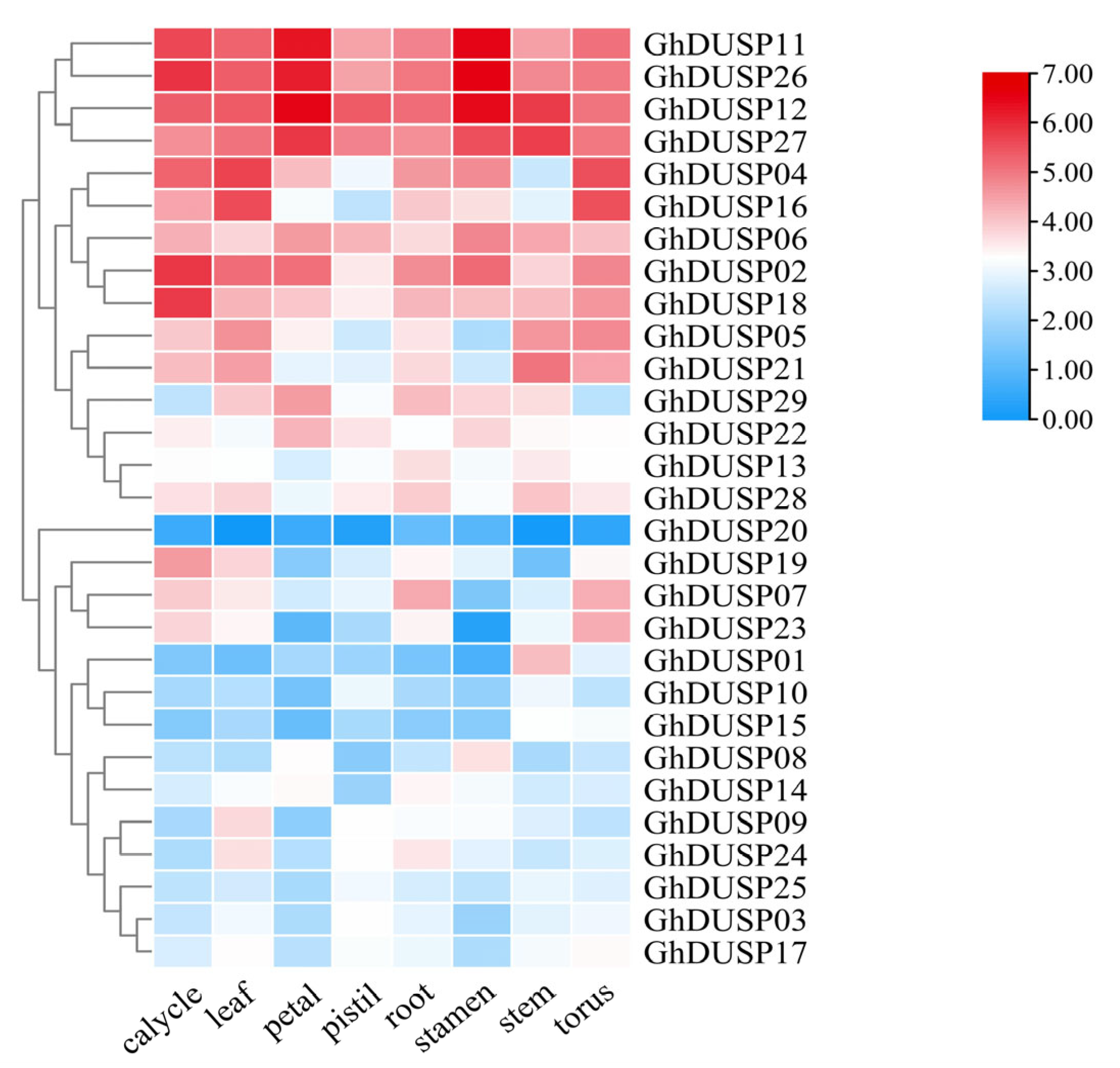
2.7. Tissue-Specific Examination of Gene Expression within the GhDUSPs
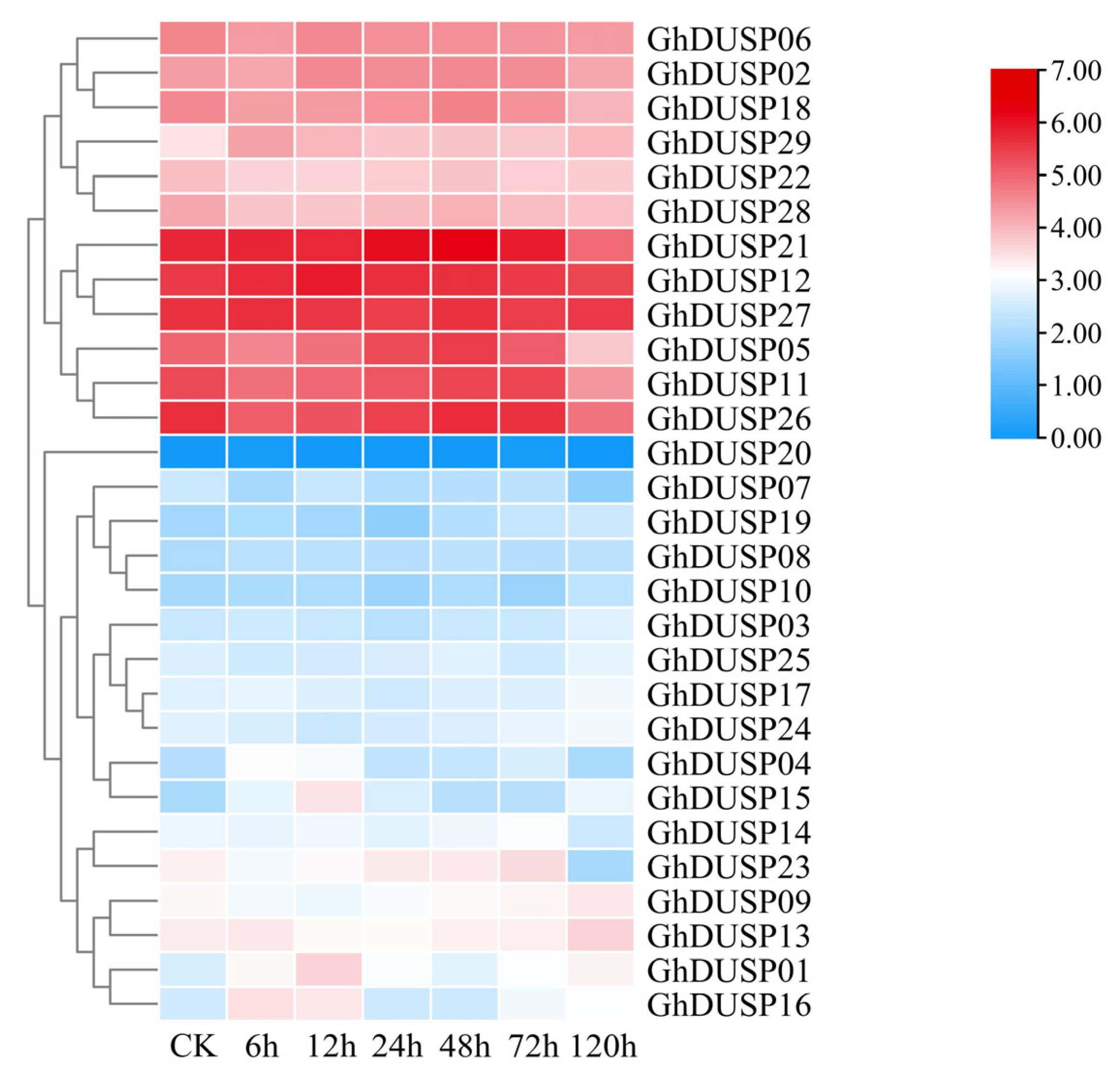
2.8. Exploring the Impact of Various Stressors on the Expression Patterns of GhDUSPs
2.9. Expression Analysis of GhDUSPs in Verticillium Dahliae
2.10. Detection of Verticillium wilt GhDUSPs Using qRT-PCR
2.11. Functional Verification of GH_A11G3500
3. Discussion
3.1. Phosphorylation of DUSP Family Genes
3.2. Basic Analysis of DUSP Families in Four Major Cotton Species
3.3. Potential Role of DUSPs Gene in Disease Resistance of Upland Cotton
4. Materials and Methods
4.1. Gene Members Belonging to DUSPs in Cotton were Identified
4.2. Localization of DUSP Family Genes on Chromosomes
4.3. Development and Evaluation of a Phylogenetic Tree for the DUSP Gene Family in Cotton
4.4. Analysis of Conserved Patterns and Genetic Organization in the DUSPs of Cotton
4.5. Exploring the Collinearity of DUSPs in Cotton
4.6. Prediction of GhDUSPs’ Cis-Acting Components
4.7. Expression Analysis of GhDUSPs in Cotton under Different Tissues and Stresses
4.8. Examination of the Disease Resistance Expression Patterns in GhDUSPs
4.9. RNA Extraction and qRT-PCR
4.10. Silencing the Target Gene GhDUSP14
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Poulaki, E.G.; Triviza, M.F.; Malai, M.; Tjamos, S.E. A Case of Plant Vaccination: Enhancement of Plant Immunity against Verticillium Dahliae by Necrotized Spores of the Pathogen. Plants 2022, 11, 1691. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Coaker, G.; Zhou, J.M.; Dong, X. Plant Immune Mechanisms: From Reductionistic to Holistic Points of View. Mol. Plant 2020, 13, 1358–1378. [Google Scholar] [CrossRef] [PubMed]
- Jeffrey, K.L.; Camps, M.; Rommel, C.; Mackay, C.R. Targeting Dual-Specificity Phosphatases: Manipulating MAP Kinase Signalling and Immune Responses. Nat. Rev. Drug Discov. 2007, 6, 391–403. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Leary, E.; Saffaf, O.; Frank Baker, R.; Zhang, S. Overlapping Functions of YDA and MAPKKK3/MAPKKK5 Upstream of MPK3/MPK6 in Plant Immunity and Growth/Development. J. Integr. Plant Biol. 2022, 64, 1531–1542. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Nitta, Y.; Zhang, Q.; Wu, D.; Tian, H.; Lee, J.S.; Zhang, Y. Antagonistic Interactions between Two MAP Kinase Cascades in Plant Development and Immune Signaling. EMBO Rep. 2018, 19, e45324. [Google Scholar] [CrossRef]
- Xie, C.; Yang, L.; Gai, Y. MAPKKKs in Plants: Multidimensional Regulators of Plant Growth and Stress Responses. Int. J. Mol. Sci. 2023, 24, 4117. [Google Scholar] [CrossRef] [PubMed]
- Viswanath, K.K.; Kuo, S.Y.; Tu, C.W.; Hsu, Y.H.; Huang, Y.W.; Hu, C.C. The Role of Plant Transcription Factors in the Fight against Plant Viruses. Int. J. Mol. Sci. 2023, 24, 8433. [Google Scholar] [CrossRef]
- Wang, Y.; Pruitt, R.N.; Nürnberger, T.; Wang, Y. Evasion of Plant Immunity by Microbial Pathogens. Nat. Rev. Microbiol. 2022, 20, 449–464. [Google Scholar] [CrossRef]
- Wang, D.; Wei, L.; Liu, T.; Ma, J.; Huang, K.; Guo, H.; Huang, Y.; Zhang, L.; Zhao, J.; Tsuda, K.; et al. Suppression of ETI by PTI Priming to Balance Plant Growth and Defense through an MPK3/MPK6-WRKYs-PP2Cs Module. Mol. Plant 2023, 16, 903–918. [Google Scholar] [CrossRef]
- Das, D.; Seemann, J.; Greenstein, D.; Schedl, T.; Arur, S. Reevaluation of the Role of LIP-1 as an ERK/MPK-1 Dual Specificity Phosphatase in the C. Elegans Germline. Proc. Natl. Acad. Sci. USA 2022, 119, e21136491192022. [Google Scholar] [CrossRef]
- Lang, R.; Raffi, F. Dual-Specificity Phosphatases in Immunity and Infection: An Update. Int. J. Mol. Sci. 2019, 20, 2710. [Google Scholar] [CrossRef] [PubMed]
- Kondoh, K.; Nishida, E. Regulation of MAP Kinases by MAP Kinase Phosphatases. Biochim. Biophys. Acta Mol. Cell Res. 2007, 1773, 1227–1237. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, S. Mitogen-activated Protein Kinase Cascades in Plant Signaling. J. Integr. Plant Biol. 2022, 64, 301–341. [Google Scholar] [CrossRef] [PubMed]
- Pulido, R.; Lang, R. Dual Specificity Phosphatases: From Molecular Mechanisms to Biological Function. Int. J. Mol. Sci. 2019, 20, 4372. [Google Scholar] [CrossRef] [PubMed]
- Berry, H.M.; Argueso, C.T. More than Growth: Phytohormone-Regulated Transcription Factors Controlling Plant Immunity, Plant Development and Plant Architecture. Curr. Opin. Plant Biol. 2022, 70, 102309. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tian, H.; Chen, D.; Zhang, H.; Sun, M.; Chen, S.; Qin, Z.; Ding, Z.; Dai, S. Cysteine-Rich Receptor-like Protein Kinases: Emerging Regulators of Plant Stress Responses. Trends Plant Sci. 2023, 28, 776–794. [Google Scholar] [CrossRef] [PubMed]
- Hahn, C.K.; West, R.J.; Macari, E.R.; Schaeffer, E.K.; Lowrey, C.H. Dual-Specificity Phosphatases (DUSPs) Are Potential Targets for Pharmacologic Induction of Fetal Hemoglobin. Blood 2010, 116, 2075. [Google Scholar] [CrossRef]
- Ramkissoon, A.; Chaney, K.E.; Milewski, D.; Williams, K.B.; Williams, R.L.; Choi, K.; Miller, A.; Kalin, T.V.; Pressey, J.G.; Szabo, S.; et al. Targeted Inhibition of the Dual Specificity Phosphatases DUSP1 and DUSP6 Suppress MPNST Growth via JNK. Clin. Cancer Res. 2019, 25, 4117–4127. [Google Scholar] [CrossRef] [PubMed]
- Tögel, L.; Nightingale, R.; Wu, R.; Chüeh, A.C.; Al-Obaidi, S.; Luk, I.; Dávalos-Salas, M.; Chionh, F.; Murone, C.; Buchanan, D.D. DUSP5 Is Methylated in CIMP-High Colorectal Cancer but Is Not a Major Regulator of Intestinal Cell Proliferation and Tumorigenesis. Sci. Rep. 2018, 8, 1767. [Google Scholar] [CrossRef]
- Gao, P.P.; Qi, X.W.; Sun, N.; Sun, Y.Y.; Zhang, Y.; Tan, X.N.; Ding, J.; Han, F.; Zhang, Y. The Emerging Roles of DualSpecificity Phosphatases and Their Specific Characteristics in Human Cancer. Biochim. Biophys. Acta Rev. Cancer 2021, 1876, 188562. [Google Scholar] [CrossRef]
- Chandrasekhar, A.; Komirishetty, P.; Areti, A.; Krishnan, A.; Zochodne, D.W. Dual Specificity Phosphatases Support Axon Plasticity and Viability. Mol. Neurobiol. 2021, 58, 391–407. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P. The Regulation of Protein Function by Multisite Phosphorylation—A 25 Year Update. Trends Biochem. Sci. 2000, 25, 596–601. [Google Scholar] [CrossRef] [PubMed]
- Subbannayya, Y.; Pinto, S.M.; Bösl, K.; Prasad, T.S.K.; Kandasamy, R.K. Dynamics of Dual Specificity Phosphatases and Their Interplay with Protein Kinases in Immune Signaling. Int. J. Mol. Sci. 2019, 20, 2086. [Google Scholar] [CrossRef] [PubMed]
- Lang, R.; Hammer, M.; Mages, J. DUSP Meet Immunology: Dual Specificity MAPK Phosphatases in Control of the Inflammatory Response. J. Immunol. 2006, 177, 7497–7504. [Google Scholar] [CrossRef] [PubMed]
- Patterson, K.I.; Brummer, T.; O’brien, P.M.; Daly, R.J. Dual-Specificity Phosphatases: Critical Regulators with Diverse Cellular Targets. Biochem. J. 2009, 418, 475–489. [Google Scholar] [CrossRef] [PubMed]
- Thompson, E.M.; Stoker, A.W. A Review of DUSP26: Structure, Regulation and Relevance in Human Disease. Int. J. Mol. Sci. 2021, 22, 776. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Xiong, J.; Du, Y.; Huang, Y.; Zhao, J. Dual-Specificity Phosphatases and Kidney Diseases. Kidney Dis. 2022, 8, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Relav, L.; Dos Santos, E.C.; Zamberlam, G.; Price, C.A. Abundance of Dual Specificity Phosphatase (DUSP) 1 and DUSP6 mRNA Is Regulated by Hippo Signaling in Bovine Pre-Ovulatory Granulosa Cells. Reprod. Sci. 2023, 30, 1782–1788. [Google Scholar] [CrossRef] [PubMed]
- De Jong, R.N.; Ab, E.; Diercks, T.; Truffault, V.; Daniëls, M.; Kaptein, R.; Folkers, G.E. Solution Structure of the Human Ubiquitin-Specific Protease 15 DUSP Domain. J. Biol. Chem. 2006, 281, 5026–5031. [Google Scholar] [CrossRef]
- Prabhakar, S.; Asuthkar, S.; Lee, W.; Chigurupati, S.; Zakharian, E.; Tsung, A.J.; Velpula, K.K. Targeting DUSPs in Glioblastomas—Wielding a Double-edged Sword? Cell Biol. Int. 2014, 38, 145–153. [Google Scholar] [CrossRef]
- Muda, M.; Theodosiou, A.; Gillieron, C.; Smith, A.; Chabert, C.; Camps, M.; Boschert, U.; Rodrigues, N.; Davies, K.; Ashworth, A.; et al. The Mitogen-Activated Protein Kinase Phosphatase-3 N-Terminal Noncatalytic Region is Responsible for Tight Substrate Binding and Enzymatic Specificity. J. Biol. Chem. 1998, 273, 9323–9329. [Google Scholar] [CrossRef]
- Wei, C.H.; Min, H.G.; Kim, M.; Kim, G.H.; Chun, H.J.; Ryu, S.E. Two Intermediate States of the Conformational Switch in Dual Specificity Phosphatase 13a. Pharmacol. Res. 2018, 128, 211–219. [Google Scholar] [CrossRef]
- Bernoux, M.; Zetzsche, H.; Stuttmann, J. Connecting the Dots between Cell Surface- and Intracellular-Triggered Immune Pathways in Plants. Curr. Opin. Plant Biol. 2022, 69, 102276. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, H.; Wang, P.; Zhong, H.; Liu, W.; Zhang, S.; Xiong, L.; Wu, Y.; Xia, Y. Arabidopsis Extra-Large G Protein 1 (XLG1) Functions Together with XLG2 and XLG3 in PAMP-triggered MAPK Activation and Immunity. J. Integr. Plant Biol. 2023, 65, 825–837. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.T.; Kim, S.H.; Kim, K.E.; Bahk, S.; Nguyen, X.C.; Kim, M.G.; Hong, J.C.; Chung, W.S. Ca2+/CaM Increases the Necrotrophic Pathogen Resistance through the Inhibition of a CaM-Regulated Dual-Specificity Protein Phosphatase 1 in Arabidopsis. Plant Biotechnol. Rep. 2022, 16, 71–78. [Google Scholar] [CrossRef]
- Liu, R.; Liu, Y.; Ye, N.; Zhu, G.; Chen, M.; Jia, L.; Xia, Y.; Shi, L.; Jia, W.; Zhang, J. AtDsPTP1 Acts as a Negative Regulator in Osmotic Stress Signalling during Arabidopsis Seed Germination and Seedling Establishment. J. Exp. Bot. 2015, 66, 1339–1353. [Google Scholar] [CrossRef]
- Ulm, R.; Revenkova, E.; di Sansebastiano, G.-P.; Bechtold, N.; Paszkowski, J. Mitogen-Activated Protein Kinase Phosphatase Is Required for Genotoxic Stress Relief in Arabidopsis. Genes Dev. 2001, 15, 699–709. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.T. Study on the Relationship between Protein Phosphatase TOPP4 and PP2A in Arabidopsis and the Participation of TOPP4 in Geophily. Ph.D. Thesis, Lanzhou University, Lanzhou, China, 2021. [Google Scholar] [CrossRef]
- Xu, R. Study on the Regulatory Mechanism of Bispecific Protein Phosphatase AtDsPTP1 in Arabidopsis. Ph.D. Thesis, Soochow University, Suzhou, China, 2020. [Google Scholar] [CrossRef]
- Yoo, J.H.; Cheong, M.S.; Park, C.Y.; Moon, B.C.; Kim, M.C.; Kang, Y.H.; Park, H.C.; Choi, M.S.; Lee, J.H.; Jung, W.Y.; et al. Regulation of the Dual Specificity Protein Phosphatase, DsPTP1, through Interactions with Calmodulin. J. Biol. Chem. 2004, 279, 848–858. [Google Scholar] [CrossRef]
- Mu, C.; Li, F.J.; Tian, X.L. Regulation of GhANN8 Dephosphorylation Mediated by Bispecific Tyrosine Protein Phosphatase GhDsPTP1 on Salt Tolerance of Cotton. In Proceedings of the 2017 Annual Conference and the 9th Member Congress of Cotton Branch of Chinese Agricultural Society, Zhengzhou, China, 7–9 August 2017. [Google Scholar]
- Yu, Q.; Zhang, L.; Sun, D.; Hu, Y.; Li, P.; Zhang, X.; Ding, L.; Zhou, L.; Guan, Z.; Fang, W.; et al. Interaction between MAPKs and MKPs in Hexaploid Chrysanthemum Illuminates Functional Paralogue Diversification in Polyploids. Hortic. Plant J. 2023, 9, 158–168. [Google Scholar] [CrossRef]
- Gao, M.; Schornack, S. Antibodies for a Bespoke Plant Immunity. Cell Host Microbe 2023, 31, 683–684. [Google Scholar] [CrossRef]
- Smailys, J.; Jiang, F.; Prioleau, T.; Kelley, K.; Mitchell, O.; Nour, S.; Ali, L.; Buchser, W.; Zavada, L.; Hinton, S.D. The DUSP Domain of Pseudophosphatase MK-STYX Interacts with G3BP1 to Decrease Stress Granules. Arch. Biochem. Biophys. 2023, 744, 109702. [Google Scholar] [CrossRef] [PubMed]
- Tzipilevich, E.; Russ, D.; Dangl, J.L.; Benfey, P.N. Plant Immune System Activation Is Necessary for Efficient Root Colonization by Auxin-Secreting Beneficial Bacteria. Cell Host Microbe 2021, 29, 1507–1520.e4. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Lu, X.; Gou, X. Receptor-like Protein Kinases in Plant Reproduction: Current Understanding and Future Perspectives. Plant Commun. 2022, 3, 100273. [Google Scholar] [CrossRef]
- Li, X.; Zhang, J.; Shi, H.; Li, B.; Li, J. Rapid Responses: Receptor-like Kinases Directly Regulate the Functions of Membrane Transport Proteins in Plants. J. Integr. Plant Biol. 2022, 64, 1303–1309. [Google Scholar] [CrossRef]
- Shu, L.-J.; Kahlon, P.S.; Ranf, S. The Power of Patterns: New Insights into Pattern-triggered Immunity. New Phytol. 2023, 240, 960–967. [Google Scholar] [CrossRef]
- Jiang, Y.; Ding, P. Calcium Signaling in Plant Immunity: A Spatiotemporally Controlled Symphony. Trends Plant Sci. 2023, 28, 74–89. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Morea, F.A.; Liu, J.; Shan, L.; He, P. Malectin-like Receptor Kinases as Protector Deities in Plant Immunity. Nat. Plants 2021, 8, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Outram, M.A.; Dodds, P.N. Wheeling in a New Era in Plant Immunity. Nat. Plants 2022, 8, 1142–1143. [Google Scholar] [CrossRef] [PubMed]
- Garner, C.M.; Kim, S.H.; Spears, B.J.; Gassmann, W. Express Yourself: Transcriptional Regulation of Plant Innate Immunity. Semin. Cell Dev. Biol. 2016, 56, 150–162. [Google Scholar] [CrossRef]
- Gibson, S.A.; Benveniste, E.N. Protein Kinase CK2: An Emerging Regulator of Immunity. Trends Immunol. 2018, 39, 82–85. [Google Scholar] [CrossRef]
- Förderer, A.; Yu, D.; Li, E.; Chai, J. Resistosomes at the Interface of Pathogens and Plants. Curr. Opin. Plant Biol. 2022, 67, 102212. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Qi, J.; Hu, M.; Bi, G.; Zhou, J.-M.; Han, G.-Z. The Origin and Evolution of a Plant Resistosome. Plant Cell. 2022, 34, 1600–1620. [Google Scholar] [CrossRef] [PubMed]
- Ngou, B.P.M.; Jones, J.D.G.; Ding, P. Plant Immune Networks. Trends Plant Sci. 2022, 27, 255–273. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.-W.; Niu, Y.; Jia, Y.; Ordon, J.; Copeland, C.; Emonet, A.; Geldner, N.; Guan, R.; Stolze, S.C.; Nakagami, H.; et al. Coordination of Microbe–Host Homeostasis by Crosstalk with Plant Innate Immunity. Nat. Plants 2021, 7, 814–825. [Google Scholar] [CrossRef] [PubMed]
- Grumont, R.J.; Rasko, J.E.J.; Strasser, A.; Gerondakis, S. Activation of the Mitogen-Activated Protein Kinase Pathway Induces Transcription of the PAC-1 Phosphatase Gene. Mol. Cell. Biol. 1996, 16, 2913–2921. [Google Scholar] [CrossRef] [PubMed]
- Enebe, M.C.; Erasmus, M. Susceptibility and Plant Immune Control—A Case of Mycorrhizal Strategy for Plant Colonization, Symbiosis, and Plant Immune Suppression. Front. Microbiol. 2023, 14, 1178258. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Wang, L.; Zhao, H.; Zhang, Y.; Chen, Z.; Xu, L.; Yi, K. The Origin and Evolution of Salicylic Acid Signaling and Biosynthesis in Plants. Mol. Plant 2023, 16, 245–259. [Google Scholar] [CrossRef]
- Hemenway, E.A.; Gehring, M. Epigenetic Regulation during Plant Development and the Capacity for Epigenetic Memory. Annu. Rev. Plant Biol. 2023, 74, 87–109. [Google Scholar] [CrossRef]
- Larson, E.R.; Armstrong, E.M.; Harper, H.; Knapp, S.; Edwards, K.J.; Grierson, D.; Poppy, G.; Chase, M.W.; Jones, J.D.G.; Bastow, R.; et al. One Hundred Important Questions for Plant Science—Reflecting on a Decade of Plant Research. New Phytol. 2023, 238, 464–469. [Google Scholar] [CrossRef]
- Liu, J.Y. Function Analysis of Bispecific Phosphatase IBR5 in Temperature-Dependent Defense Response of Arabidopsis. Ph.D. Thesis, China Agricultural University, Beijing, China, 2016. [Google Scholar]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, Y.; Deng, X.; Zhao, J.; Ning, S.; Gu, A.; Chen, Q.; Qu, Y. Revealing the Complete Bispecific Phosphatase Genes (DUSPs) across the Genome and Investigating the Expression Patterns of GH_A11G3500 Resistance against Verticillium wilt. Int. J. Mol. Sci. 2024, 25, 4500. https://doi.org/10.3390/ijms25084500
Deng Y, Deng X, Zhao J, Ning S, Gu A, Chen Q, Qu Y. Revealing the Complete Bispecific Phosphatase Genes (DUSPs) across the Genome and Investigating the Expression Patterns of GH_A11G3500 Resistance against Verticillium wilt. International Journal of Molecular Sciences. 2024; 25(8):4500. https://doi.org/10.3390/ijms25084500
Chicago/Turabian StyleDeng, Yahui, Xiaojuan Deng, Jieyin Zhao, Shuo Ning, Aixing Gu, Quanjia Chen, and Yanying Qu. 2024. "Revealing the Complete Bispecific Phosphatase Genes (DUSPs) across the Genome and Investigating the Expression Patterns of GH_A11G3500 Resistance against Verticillium wilt" International Journal of Molecular Sciences 25, no. 8: 4500. https://doi.org/10.3390/ijms25084500
APA StyleDeng, Y., Deng, X., Zhao, J., Ning, S., Gu, A., Chen, Q., & Qu, Y. (2024). Revealing the Complete Bispecific Phosphatase Genes (DUSPs) across the Genome and Investigating the Expression Patterns of GH_A11G3500 Resistance against Verticillium wilt. International Journal of Molecular Sciences, 25(8), 4500. https://doi.org/10.3390/ijms25084500