
The Deep Proteomics Approach Identified Extracellular Vesicular Proteins Correlated to Extracellular Matrix in Type One and Two Endometrial Cancer

 , , ,
, , ,  ,
,  ,
,  ,
,  ,
,  , , ,
, , ,  and
and
Abstract
:1. Introduction
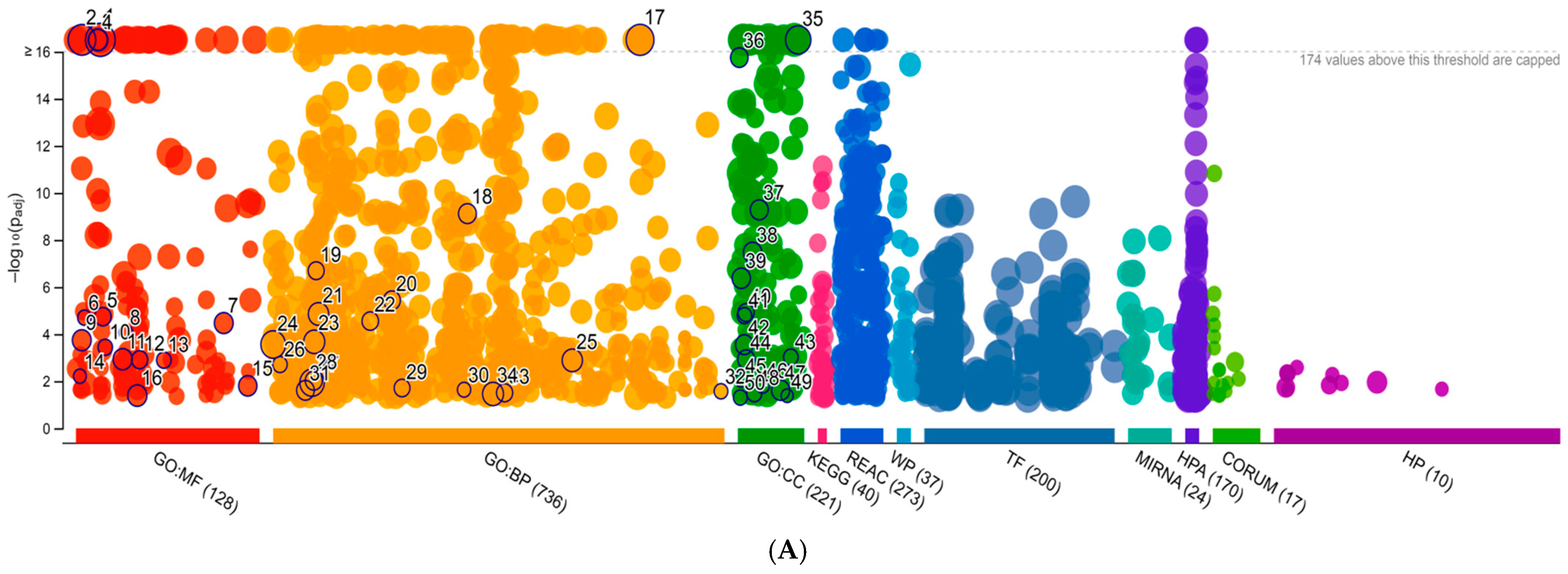
2. Results
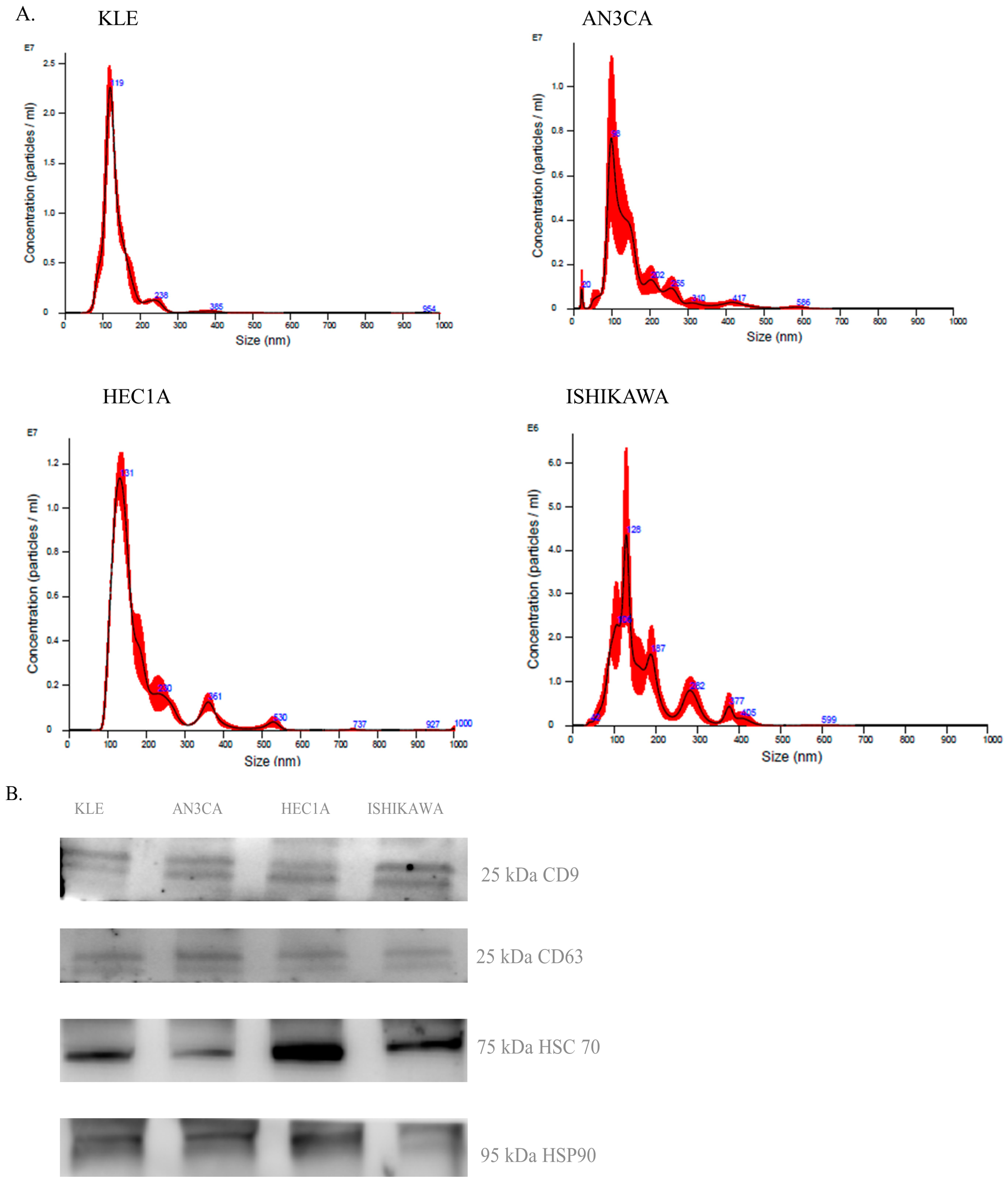
2.1. Extracellular Vesicle Characterization
Proteomics Study
2.2. Western Blotting
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. EV Isolation
4.3. Patients
4.4. Proteome Analysis
4.5. Western Blotting
Nanoparticle Tracking Analysis
4.6. Bioinformatic Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Chava, S.; Gupta, R. Identification of the Mutational Landscape of Gynecological Malignancies. J. Cancer 2020, 11, 4870–4883. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.T. Risk factors for endometrial cancer. Ceska Gynekol. 2013, 78, 448–459. [Google Scholar] [PubMed]
- Ebring, C.; Marlin, R.; Macni, J.; Vallard, A.; Bergerac, S.; Beaubrun-Renard, M.; Joachim, C.; JeanLaurent, M. Type II endometrial cancer: Incidence, overall and disease-free survival in Martinique. PLoS ONE 2023, 18, e0278757. [Google Scholar] [CrossRef] [PubMed]
- Vanderstraeten, A.; Tuyaerts, S.; Amant, F. The immune system in the normal endometrium and implications for endometrial cancer development. J. Reprod. Immunol. 2015, 109, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Yang, Y.; Zhang, Y.; Jiang, S.; Li, X.; Wan, J. Identification of prognostic immune-related genes in the tumor microenvironment of endometrial cancer. Aging 2020, 12, 3371–3387. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, M.; Wu, Y. Features of the immunosuppressive tumor microenvironment in endometrial cancer based on molecular subtype. Front. Oncol. 2023, 13, 1278863. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Weaver, V.M.; Werb, Z. The extracellular matrix: A dynamic niche in cancer progression. J. Cell Biol. 2012, 13, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Sheta, M.; Taha, E.A.; Lu, Y.; Eguchi, T. Extracellular Vesicles: New Classification and Tumor Immunosuppression. Biology 2023, 12, 110. [Google Scholar] [CrossRef]
- Yu, W.; Hurley, J.; Roberts, D.; Chakrabortty, S.K.; Enderle, D.; Noerholm, M.; Breakefield, X.O.; Skog, J.K. Exosome-based liquid biopsies in cancer: Opportunities and challenges. Ann. Oncol. 2021, 32, 466–477. [Google Scholar] [CrossRef]
- Mariscal, J.; Fernandez-Puente, P.; Calamia, V.; Abalo, A.; Santacana, M.; Matias-Guiu, X.; Lopez-Lopez, R.; Gil-Moreno, A.; Alonso-Alconada, L.; Abal, M. Proteomic Characterization of Epithelial-Like Extracellular Vesicles in Advanced Endometrial Cancer. J. Proteome Res. 2019, 18, 1043–1053. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tang, Y.; Chen, X.; Sun, X.; Zhao, M.; Chen, Q. Therapeutic potential of miRNAs in placental extracellular vesicles in ovarian and endometrial cancer. Hum. Cell 2024, 37, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Esfandyari, S.; Elkafas, H.; Chugh, R.M.; Park, H.S.; Navarro, A.; Al-Hendy, A. Exosomes as Biomarkers for Female Reproductive Diseases Diagnosis and Therapy. Int. J. Mol. Sci. 2021, 22, 2165. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.J.; Ashraf, A.; Chung, E.J. Extracellular Vesicles as Regulators of the Extracellular Matrix. Bioengineering 2023, 10, 136. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef]
- Chen, L.; Fang, B.; Giorgianni, F.; Gingrich, J.R.; Beranova-Giorgianni, S. Investigation of phosphoprotein signatures of archived prostate cancer tissue specimens via proteomic analysis. Electrophoresis 2011, 32, 1984–1991. [Google Scholar] [CrossRef] [PubMed]
- Sommella, E.; Capaci, V.; Aloisio, M.; Salviati, E.; Campiglia, P.; Molinario, G.; Licastro, D.; Di Lorenzo, G.; Romano, F.; Ricci, G.; et al. A Label-Free Proteomic Approach for the Identification of Biomarkers in the Exosome of Endometrial Cancer Serum. Cancers 2022, 14, 6262. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-Rodriguez, P.; Selheim, F.; Hernandez-Valladares, M. Mass Spectrometry-Based Proteomics Workflows in Cancer Research: The Relevance of Choosing the Right Steps. Cancers 2023, 15, 555. [Google Scholar] [CrossRef] [PubMed]
- Brennan, K.; Martin, K.; FitzGerald, S.P.; Sullivan, J.O.; Wu, Y.; Blanco, A.; CRichardson, C.; Mc Gee, M.M. A comparison of methods for the isolation and separation of extracellular vesicles from protein and lipid particles in human serum. Sci. Rep. 2020, 10, 1039. [Google Scholar] [CrossRef]
- Patel, G.K.; Khan, M.A.; Zubair, H.; Sanjeev Kumar Srivastava, S.K.; Khushman, M.; Singh, S.; Pratap Singh, A. Comparative analysis of exosome isolation methods using culture supernatant for optimum yield, purity and downstream applications. Sci. Rep. 2019, 9, 5335. [Google Scholar] [CrossRef]
- Kwon, Y.W.; Jo, H.S.; Bae, S.; Seo, Y.; Song, P.; Song, M.; Yoon, J.H. Application of Proteomics in Cancer: Recent Trends and Approaches for Biomarkers Discovery. Front. Med. 2021, 8, 747333. [Google Scholar] [CrossRef]
- Matzinger, M.; Schmücker, A.; Yelagandula, R.; Stejskal, K.; Krššáková, G.; Berger, F.; Mechtler, K.; Mayer, R.L. Micropillar arrays, wide window acquisition and AI-based data analysis improve comprehensiveness in multiple proteomic applications. Nat. Commun. 2024, 15, 1019. [Google Scholar] [CrossRef] [PubMed]
- Popova, N.V.; Jücker, M. The Functional Role of Extracellular Matrix Proteins in Cancer. Cancers 2022, 14, 238. [Google Scholar] [CrossRef] [PubMed]
- Winkler, J.; Abisoye-Ogunniyan, A.; Metcalf, K.J.; Werb, Z. Concepts of extracellular matrix remodelling in tumour progression and metastasis. Nat. Commun. 2020, 11, 5120. [Google Scholar] [CrossRef] [PubMed]
- Gusman, H.; Travis, J.; Helmerhorst, E.J.; Potempa, J.; Troxler, R.F.; Oppenheim, F.G. Salivary histatin 5 is an inhibitor of both host and bacterial enzymes implicated in periodontal disease. Infect. Immun. 2001, 69, 1402–1408. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Li, H. Prognostic values of tumoral MMP2 and MMP9 overexpression in breast cancer: A systematic review and meta-analysis. BMC Cancer 2021, 21, 149. [Google Scholar] [CrossRef] [PubMed]
- Maity, G.; Sen, T.; Chatterjee, A. Laminin Induces Matrix Metalloproteinase-9 Expression and Activation in Human Cervical Cancer Cell Lineproposal. J. Cancer Res. Clin. Oncol. 2011, 137, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Li, Y.; Hu, S.; Chen, Y.; Gao, L.; Liu, D.; Guo, H.; Yang, Y. Clinical significance of matrix metalloproteinase-2 in endometrial cancer: A systematic review and meta-analysis. Medicine 2018, 97, e10994. [Google Scholar] [CrossRef]
- Chattopadhyay, N.; Mitra, A.; Frei, E.; Chatterjee, A. Human cervical tumor cell (SiHa) surface alphavbeta3 integrin receptor has associated matrix metalloproteinase (MMP-2) activity. J. Cancer Res. Clin. Oncol. 2001, 127, 653–658. [Google Scholar] [CrossRef]
- Goldberg, G.I.; Marmer, B.L.; Grant, G.A.; Eisen, A.Z.; Wilhelm, S.; He, C.S. Human 72-kilodalton type IV collagenase forms a complex with a tissue inhibitor of metalloproteases designated TIMP-2. Proc. Natl. Acad. Sci. USA 1989, 86, 8207–8211. [Google Scholar] [CrossRef]
- Graesslin, O.; Cortez, A.; Uzan, C.; Birembaut, P.; Quereux, C.; Daraï, E. Endometrial tumor invasiveness is related to metalloproteinase 2 and tissue inhibitor of metalloproteinase 2 expressions. Int. J. Gynecol. Cancer 2006, 16, 1911–1917. [Google Scholar] [CrossRef]
- Schröpfer, A.; Kammerer, U.; Kapp, M.; Dietl, J.; Feix, S.; Anacker, J. Expression Pattern of Matrix Metalloproteinases in Human Gynecological Cancer Cell Lines. BMC Cancer 2010, 10, 553. [Google Scholar] [CrossRef]
- Rak, B.; Mehlich, D.; Garbicz, F.; Domosud, Z.; Paskal, W.; Marczewska, J.M.; Włodarski, P.K. Post-transcriptional Regulation of MMP16 and TIMP2 Expression via miR-382, miR-410 and miR-200b in Endometrial Cancer. Cancer Genom. Proteom. 2017, 14, 389–401. [Google Scholar]
- Dai, Y.; Xia, W.; Song, T.; Su, X.; Li, J.; Li, S.; Chen, Y.; Wang, W.; Ding, H.; Liu, X.; et al. MicroRNA-200b is overexpressed in endometrial adenocarcinomas and enhances MMP2 activity by downregulating TIMP2 in human endometrial cancer cell line HEC-1A cells. Nucleic Acid. Ther. 2013, 23, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Chen, Z.; Zhang, Y.; Xiao, S.; Zhang, W.; Hu, X.; Xiao, Q.; Liu, Q.; Wang, X. Flotillin-1 is a prognostic biomarker for glioblastoma and promotes cancer development through enhancing invasion and altering tumour microenvironment. J. Cell Mol. Med. 2023, 27, 392–402. [Google Scholar] [CrossRef]
- Zhan, Z.; Ye, M.; Jin, X. The roles of FLOT1 in human diseases (Review). Mol. Med. Rep. 2023, 28, 212. [Google Scholar] [CrossRef]
- Winship, A.L.; Rainczuk, K.; Dimitriadis, E. Flotillin-1 protein is upregulated in human endometrial cancer and localization shifts from epithelial to stromal with increasing tumor grade. Cancer Investig. 2016, 34, 26–31. [Google Scholar] [CrossRef]
- Song, T.; Hu z Chong Zeng, C.; Luo, H.; Liu, J. FLOT1, stabilized by WTAP/IGF2BP2 mediated N6-methyladenosine modification, predicts poor prognosis and promotes growth and invasion in gliomas. Heliyon 2023, 9, e16280. [Google Scholar] [CrossRef]
- Jagroop, R.; Martin, C.J.; Moorehead, R.A. Nidogen 1 regulates proliferation and migration/invasion in murine claudin-low mammary tumor cells. Oncol. Lett. 2021, 21, 52. [Google Scholar] [CrossRef]
- Pedrola, N.; Devis, L.; Llauradó, M.; Campoy, I.; Martinez-Garcia, E.; Garcia, M.; Muinelo-Romay, L.; Alonso-Alconada, L.; Abal, M.; Alameda, F.; et al. Nidogen 1 and Nuclear Protein 1: Novel targets of ETV5 transcription factor involved in endometrial cancer invasion. Clin. Exp. Metastasis 2015, 32, 467–478. [Google Scholar] [CrossRef]
- Ramón, L.A.; Braza-Boïls, A.; Gilabert, J.; Chirivella, M.; España, F.; Estellés, A.; Gilabert-Estellés, J. microRNAs related to angiogenesis are dysregulated in endometrioid endometrial cancer. Hum. Reprod. 2012, 27, 3036–3045. [Google Scholar] [CrossRef] [PubMed]
- ‘Cui, D.; Cui, X.; Xu, X.; Zhang, W.; Yu, Y.; Gao, Y.; Mei, C.; Zheng, W. Identification of TLN1 as a prognostic biomarker to effect cell proliferation and differentiation in acute myeloid leukemia. BMC Cancer 2022, 22, 1027. [Google Scholar]
- Wegener, K.L.; Partridge, A.W.; Han, J.; Pickford, A.R.; Liddington, R.C.; Ginsberg, M.H.; Campbell, I.D. Structural basis of integrin activation by talin. Cell 2007, 128, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Goult, B.T.; Yan, J.; Schwartz, M.A. Talin as a mechanosensitive signaling hub. J. Cell Biol. 2018, 217, 3776–3784. [Google Scholar] [CrossRef] [PubMed]
- Singel, S.M.; Cornelius, C.; Batten, K.; Fasciani, G.; Wright, W.E.; Lum, L.; Shay, J.W. A targeted RNAi screen of the breast cancer genome identifies KIF14 and TLN1 as genes that modulate docetaxel chemosensitivity in triple-negative breast cancer. Clin. Cancer Res. 2013, 19, 2061–2070. [Google Scholar] [CrossRef] [PubMed]
- Slater, M.; Cooper, M.; Murphy, C.R. The cytoskeletal proteins alpha-actinin, Ezrin, and talin are De-expressed in endometriosis and endometrioid carcinoma compared with normal uterine epithelium. Appl. Immunohistochem. Mol. Morphol. 2007, 15, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Sykaras, A.G.; Christofidis, K.; Politi, E.; Theocharis, S. Exosomes on Endometrial Cancer: A Biomarkers Treasure Trove? Cancers 2022, 14, 1733. [Google Scholar] [CrossRef]
- Fan, J.T.; Zhou, Z.Y.; Luo, Y.L.; Luo, Q.; Chen, S.B.; Zhao, J.C.; Chen, Q.R. Exosomal lncRNA NEAT1 from cancer-associated fibroblasts facilitates endometrial cancer progression via miR-26a/b-5p-mediated STAT3/YKL-40 signaling pathway. Neoplasia 2021, 23, 692–703. [Google Scholar] [CrossRef]
- Shi, S.; Tan, Q.; Feng, F.; Huang, H.; Liang, J.; Cao, D.; Wang, Z. Identification of core genes in the progression of endometrial cancer and cancer cell-derived exosomes by an integrative analysis. Sci. Rep. 2020, 10, 9862. [Google Scholar] [CrossRef]
- Gu, S.; Ni, T.; Wang, J.; Liu, Y.; Fan, Q.; Wang, Y.; Huang, T.; Chu, Y.; Sun, X.; Wang, Y. CD47 Blockade Inhibits Tumor Progression through Promoting Phagocytosis of Tumor Cells by M2 Polarized Macrophages in Endometrial Cancer. J. Immunol. Res. 2018, 2018, 6156757. [Google Scholar] [CrossRef]
- Zhao, W.; Geng, P.; Li, Y.; Wei, X.; Cheng, J. MicroRNA-21 promotes endometrial carcinoma proliferation and invasion by targeting PTEN. Int. J. Clin. Exp. Pathol. 2017, 10, 11489–11495. [Google Scholar]
- Ura, B.; Scrimin, F.; Arrigoni, G.; Athanasakis, E.; Aloisio, M.; Monasta, L.; Ricci, G. Abnormal expression of leiomyoma cytoskeletal proteins involved in cell migration. Oncol. Rep. 2016, 35, 3094–3100. [Google Scholar] [CrossRef]
- Celsi, F.; Monasta, L.; Arrigoni, G.; Battisti, I.; Licastro, D.; Aloisio, M.; Di Lorenzo, G.; Romano, F.; Ricci, G.; Ura, B. Gel-Based Proteomic Identification of Suprabasin as a Potential New Candidate Biomarker in Endometrial Cancer. Int. J. Mol. Sci. 2022, 23, 2076. [Google Scholar] [CrossRef] [PubMed]
- Ura, B.; Scrimin, F.; Franchin, C.; Arrigoni, G.; Licastro, D.; Monasta, L.; Ricci, G. Identification of proteins with different abundance associated with cell migration and proliferation in leiomyoma interstitial fluid by proteomics. Oncol. Lett. 2017, 13, 3912–3920. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biological Functions | Adjusted p-Value |
|---|---|
| Extracellular matrix structural constituent | 4.26 × 10−24 |
| Extracellular matrix binding | 3.53 × 10−6 |
| Extracellular matrix structural constituent conferring tensile strength | 4.94 × 10−5 |
| Extracellular matrix organization | 1.52 × 10−15 |
| Cell-matrix adhesion | 6.60 × 10−8 |
| Regulation of cell-matrix adhesion | 2.64 × 10−5 |
| Extracellular matrix assembly | 2.18 × 10−3 |
| Collagen-containing extracellular matrix | 1.52 × 10−43 |
| Extracellular matrix | 2.10 × 10−42 |
| Protein complex involved in cell-matrix adhesion | 9.51 × 10−4 |
| Extracellular matrix organization | 4.25 × 10−16 |
| Degradation of the extracellular matrix | 1.96 × 10−6 |
| Cell-extracellular matrix interactions | 1.75 × 10−4 |
| Protein Accessions | Genes | Protein Descriptions | Fold Change | p Value |
|---|---|---|---|---|
| P16035 | TIMP2 | Metalloproteinase inhibitor 2 | 5.45 | 0.02 |
| P08253 | MMP2 | 72 kDa type IV collagenase | 5.002 | 0.004 |
| Q16674 | MIA | Melanoma-derived growth regulatory protein | 4.26 | 0.01 |
| O75955 | FLOT1 | Flotillin-1 | 4.06 | 0.004 |
| P07858 | CTSB | Cathepsin B | 3.57 | 0.007 |
| Q06828 | FMOD | Fibromodulin | 3.52 | 0.02 |
| P10909-2 | CLU | Isoform 2 of Clusterin | 3.5 | 0.003 |
| P51654 | GPC3 | Glypican-3 | 3.18 | 0.012 |
| P04632 | CAPNS1 | Calpain small subunit 1 | 3.143 | 0.001 |
| P07602 | PSAP | Prosaposin | 2.76 | 0.001 |
| Q02809 | PLOD1 | Procollagen-lysine,2-oxoglutarate 5-dioxygenase 1 | 2.61 | 0.008 |
| P08123 | COL1A2 | Collagen alpha-2(I) chain | 2.61 | 0.002 |
| Q99467 | CD180 | CD180 antigen | 2.47 | 0.03 |
| O60568 | PLOD3 | Multifunctional procollagen lysine hydroxylase and glycosyltransferase LH3 | 2.37 | 0.003 |
| P61586 | RHOA | Transforming protein RhoA | 2.34 | 0.04 |
| P78539-2 | SRPX | Isoform 2 of Sushi repeat-containing protein SRPX | 2.33 | 0.01 |
| Q86UW8 | HAPLN4 | Hyaluronan and proteoglycan link protein 4 | 2.26 | 0.01 |
| P03950 | ANG | Angiogenin | 2.252 | 0.03 |
| Q15063 | POSTN | Periostin | 2.1 | 0.004 |
| Q9GZM7 | TINAGL1 | Tubulointerstitial nephritis antigen-like | 2.09 | 0.03 |
| Q9NVD7 | PARVA | Alpha-parvin | 2.01 | 0.04 |
| O75325 | LRRN2 | Leucine-rich repeat neuronal protein 2 | 1.97 | 0.004 |
| Q9NQX1 | PRDM5 | PR domain zinc finger protein 5 | 1.96 | 0.02 |
| P00747 | PLG | Plasminogen | 1.95 | 0.009 |
| Q9HBI1 | PARVB | Beta-parvin | 1.89 | 0.0002 |
| P14625 | HSP90B1 | Endoplasmin | 1.82 | 0.03 |
| P07384 | CAPN1 | Calpain-1 catalytic subunit | 1.79 | 0.002 |
| O95967 | EFEMP2 | EGF-containing fibulin-like extracellular matrix protein 2 | 1.78 | 0.0005 |
| P63000-2 | RAC1 | Isoform B of Ras-related C3 botulinum toxin substrate 1 | 1.78 | 0.03 |
| P27797 | CALR | Calreticulin | 1.74 | 0.04 |
| P35590 | TIE1 | Tyrosine-protein kinase receptor Tie-1 | 1.74 | 0.01 |
| Q6UVK1 | CSPG4 | Chondroitin sulfate proteoglycan 4 | 1.71 | 0.02 |
| Q13162 | PRDX4 | Peroxiredoxin-4 | 1.64 | 0.001 |
| P35858 | IGFALS | Insulin-like growth factor-binding protein complex acid labile subunit | 1.64 | 0.01 |
| Q8NI99 | ANGPTL6 | Angiopoietin-related protein 6 | 1.62 | 0.01 |
| P04004 | VTN | Vitronectin | 1.62 | 0.01 |
| Q13751 | LAMB3 | Laminin subunit beta-3 | 1.58 | 0.01 |
| Q92819 | HAS2 | Hyaluronan synthase 2 | 1.57 | 0.02 |
| P19823 | ITIH2 | Inter-alpha-trypsin inhibitor heavy chain H2 | 1.54 | 0.03 |
| P35555 | FBN1 | Fibrillin-1 | 1.5 | 0.01 |
| P07942 | LAMB1 | Laminin subunit beta-1 | 0.62 | 0.004 |
| Q92839 | HAS1 | Hyaluronan synthase 1 | 0.59 | 0.01 |
| P11047 | LAMC1 | Laminin subunit gamma-1 | 0.59 | 0.003 |
| P20936 | RASA1 | Ras GTPase-activating protein 1 | 0.58 | 0.01 |
| Q96A84-2 | EMID1 | Isoform 2 of EMI domain-containing protein 1 | 0.55 | 0.006 |
| O15230 | LAMA5 | Laminin subunit alpha-5 | 0.36 | 0.006 |
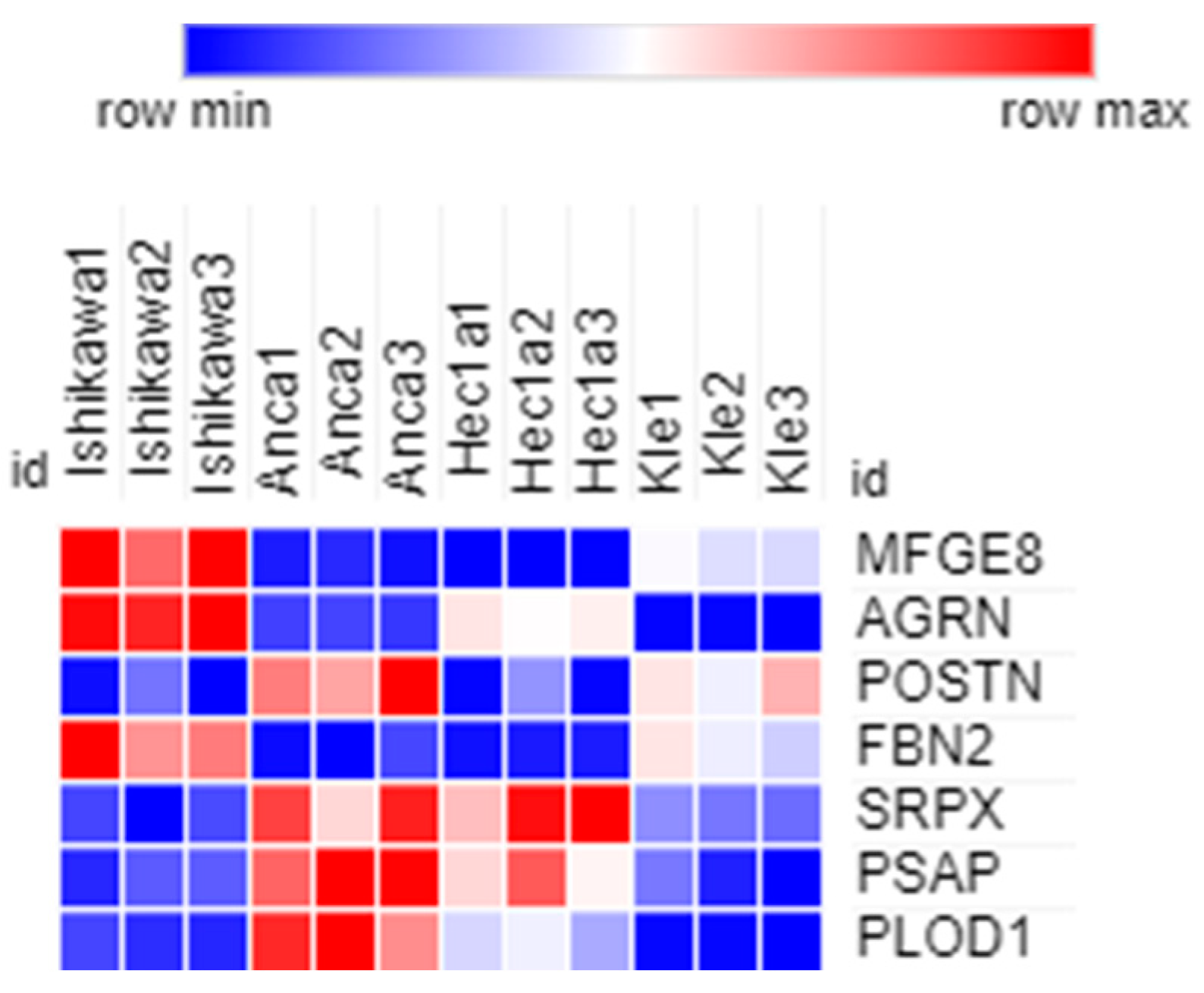
| P35556 | FBN2 | Fibrillin-2 | 0.32 | 0.005 |
| O00468-6 | AGRN | Isoform 6 of Agrin | 0.20 | 0.0002 |
| Q08431 | MFGE8 | Lactadherin | 0.12 | 0.005 |
| Protein Accessions | Genes | Protein Descriptions | Fold Change | p Value |
|---|---|---|---|---|
| P14543 | NID1 | Nidogen-1 | 14.53 | 1.48 × 10−6 |
| Q08431 | MFGE8 | Lactadherin | 8.3 | 0.001 |
| Q9Y490 | TLN1 | Talin-1 | 2.17 | 0.003 |
| P35556 | FBN2 | Fibrillin-2 | 2.13 | 0.009 |
| Q9ULV4-3 | CORO1C | Isoform 3 of coronin-1C | 1.92 | 0.04 |
| O60911 | CTSV | Cathepsin L2 | 1.73 | 0.03 |
| Q15063 | POSTN | Periostin | 1.66 | 0.01 |
| Q92743 | HTRA1 | Serine protease HTRA1 | 1.56 | 0.003 |
| Q6NUI6 | CHADL | Chondroadherin-like protein | 0.64 | 0.01 |
| P26022 | PTX3 | Pentraxin-related protein PTX3 | 0.61 | 0.01 |
| P80108 | GPLD1 | Phosphatidylinositol-glycan-specific phospholipase D | 0.6 | 0.008 |
| P07355-2 | ANXA2 | Isoform 2 of Annexin A2 | 0.59 | 0.009 |
| P26447 | S100A4 | Protein S100-A4 | 0.57 | 0.0005 |
| Q7RTU9 | STRC | Stereocilin | 0.54 | 0.04 |
| P78539-2 | SRPX | Isoform 2 of sushi repeat-containing protein SRPX | 0.51 | 0.03 |
| Q02809 | PLOD1 | Procollagen-lysine,2-oxoglutarate 5-dioxygenase 1 | 0.5 | 0.008 |
| P07339 | CTSD | Cathepsin D | 0.5 | 0.005 |
| P05121 | SERPINE1 | Plasminogen activator inhibitor 1 | 0.48 | 0.02 |
| P07602 | PSAP | Prosaposin | 0.42 | 0.01 |
| Q9UPN3 | MACF1 | Microtubule-actin cross-linking factor 1, isoforms 1/2/3/4/5 | 0.41 | 0.01 |
| P53634 | CTSC | Dipeptidyl peptidase 1 | 0.31 | 0.01 |
| Q08380 | LGALS3BP | Galectin-3-binding protein | 0.3 | 0.003 |
| O00300 | TNFRSF11B | Tumor necrosis factor receptor superfamily member 11B | 0.18 | 0.001 |
| O00468-6 | AGRN | Isoform 6 of agrin | 0.16 | 9.41 × 10−5 |
| Q16270 | IGFBP7 | Insulin-like growth factor-binding protein 7 | 0.12 | 0.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Capaci, V.; Kharrat, F.; Conti, A.; Salviati, E.; Basilicata, M.G.; Campiglia, P.; Balasan, N.; Licastro, D.; Caponnetto, F.; Beltrami, A.P.; et al. The Deep Proteomics Approach Identified Extracellular Vesicular Proteins Correlated to Extracellular Matrix in Type One and Two Endometrial Cancer. Int. J. Mol. Sci. 2024, 25, 4650. https://doi.org/10.3390/ijms25094650
Capaci V, Kharrat F, Conti A, Salviati E, Basilicata MG, Campiglia P, Balasan N, Licastro D, Caponnetto F, Beltrami AP, et al. The Deep Proteomics Approach Identified Extracellular Vesicular Proteins Correlated to Extracellular Matrix in Type One and Two Endometrial Cancer. International Journal of Molecular Sciences. 2024; 25(9):4650. https://doi.org/10.3390/ijms25094650
Chicago/Turabian StyleCapaci, Valeria, Feras Kharrat, Andrea Conti, Emanuela Salviati, Manuela Giovanna Basilicata, Pietro Campiglia, Nour Balasan, Danilo Licastro, Federica Caponnetto, Antonio Paolo Beltrami, and et al. 2024. "The Deep Proteomics Approach Identified Extracellular Vesicular Proteins Correlated to Extracellular Matrix in Type One and Two Endometrial Cancer" International Journal of Molecular Sciences 25, no. 9: 4650. https://doi.org/10.3390/ijms25094650
APA StyleCapaci, V., Kharrat, F., Conti, A., Salviati, E., Basilicata, M. G., Campiglia, P., Balasan, N., Licastro, D., Caponnetto, F., Beltrami, A. P., Monasta, L., Romano, F., Di Lorenzo, G., Ricci, G., & Ura, B. (2024). The Deep Proteomics Approach Identified Extracellular Vesicular Proteins Correlated to Extracellular Matrix in Type One and Two Endometrial Cancer. International Journal of Molecular Sciences, 25(9), 4650. https://doi.org/10.3390/ijms25094650