Muscle Contraction and Force: the Importance of an Ancillary Network, Nutrient Supply and Waste Removal

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Historical
1.2. Contemporaneous
2. Results and Discussion
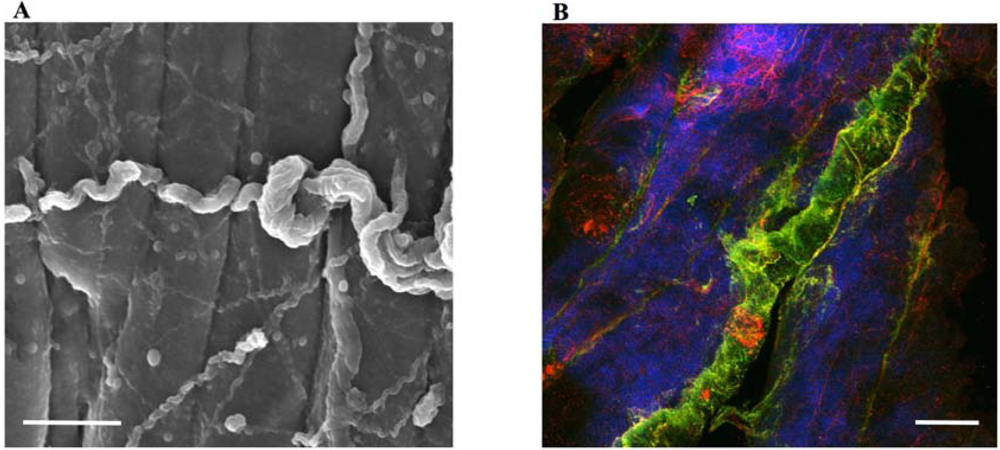
2.1. Collagen (wires)
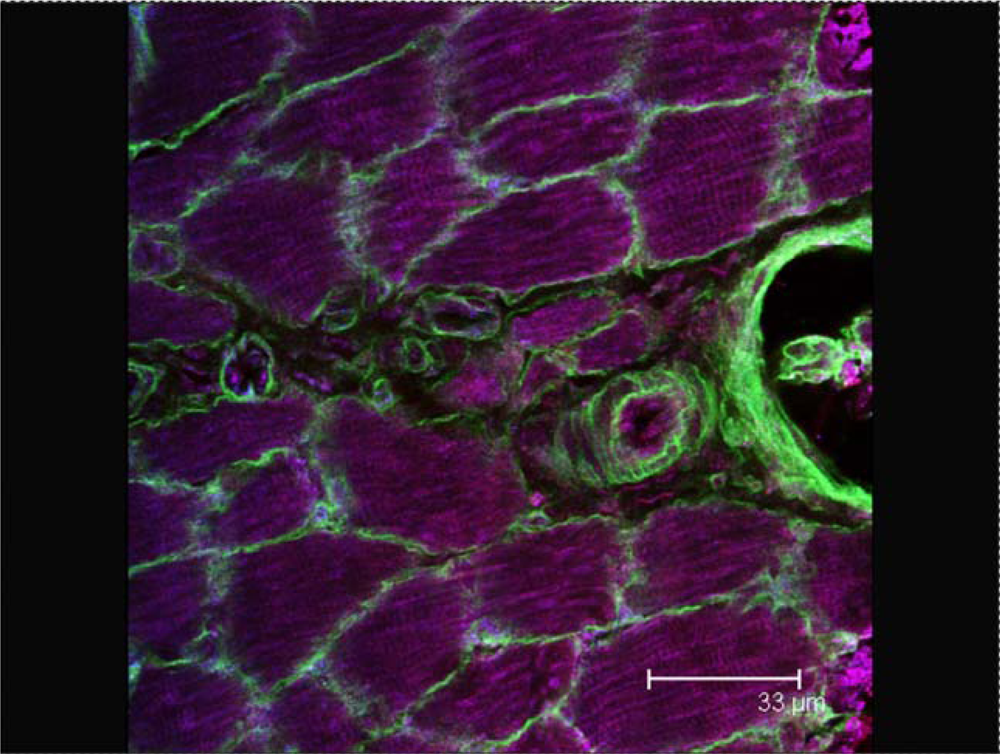
2.2. The vascular network as an optimization problem (tubes)
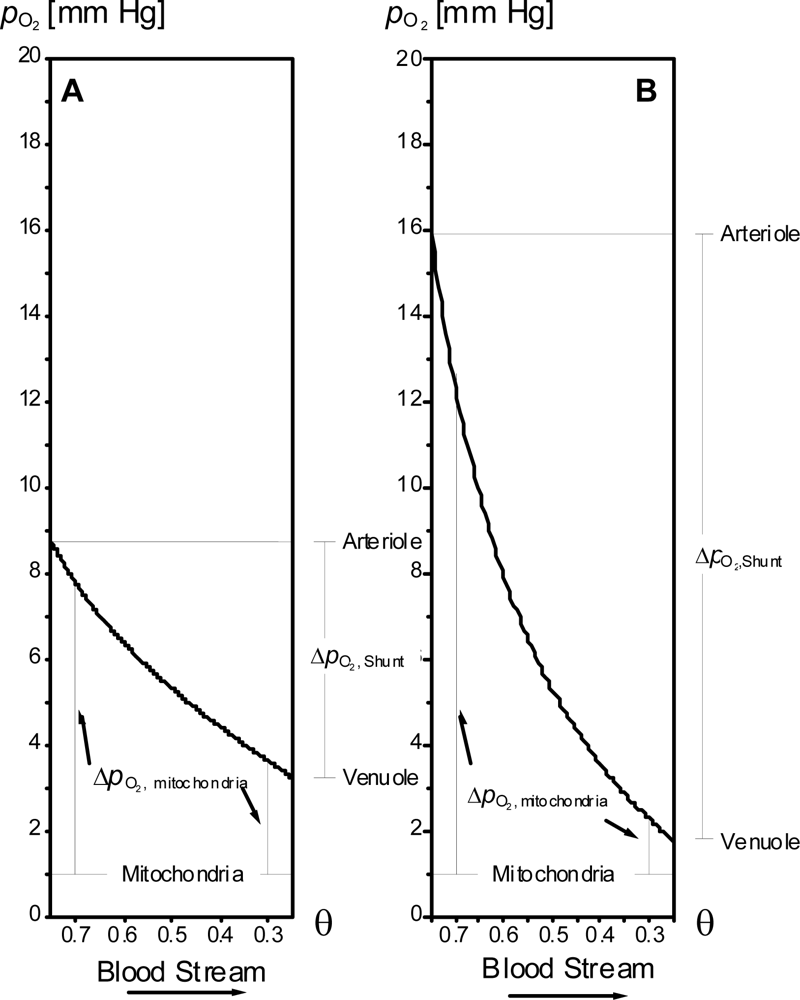
2.3. Cooperative binding of oxygen to haemoglobin – implications for optimal architecture
2.4. Waste removal in muscles (cables)
2.5. Beneficial nutrients and muscles
3. Experimental Section
3.1. Ethics
3.2. Animals
3.3. Immuno-histochemistry
3.4. Immunofluorescence
3.5. Confocal microscopy
3.6. Electron microscopy
3.7. Muscle preparation
3.8. Incubation solutions
3.9. Force measurements
3.10. Statistics
4. Conclusions
Acknowledgments
References and Notes
- Abovsky, M; Lanir, Y; Nevo, E. Tethering affects the mechanics of coronary capillaries. J. Biomech 1996, 29, 597–607. [Google Scholar]
- Bailey, AJ; Light, ND. Structure and localisation of connective tissue components in muscle. In Connective Tissue in Meat and Meat Products; Elsevier Applied Science: London, 1989; pp. 120–140. [Google Scholar]
- Barlow, TE; Haigh, AL; Walder, DN. Evidence for two vascular pathways in skeletal muscle. Clin. Sci 1961, 20, 367–385. [Google Scholar]
- Bartels, EM; Pierzynowski, SG; Harrison, AP. Benefits of alpha-ketoglutarate versus succinate on rat muscle dysfunction as a result of exposure to a uremic environment. J. Pre-Clin. Clin. Res 2007, 1, 51–54. [Google Scholar]
- Bogusch, G. Development of the vascular supply in rat skeletal muscle. Acta Anat 1984, 120, 228–233. [Google Scholar]
- Boot-Handford, RP; Tuckwell, DS; Plumb, DA; Rock, CF; Poulsom, R. A novel and highly conserved (pro (alpha) 18XXVII) with a unique expression pattern and unusual molecular characteristics establishes a new clade within the vertebrate fibrillar collagen family. J. Biol. Chem 2003, 278, 31067–31077. [Google Scholar]
- Borg, TK; Caulfield, JB. The collagen matrix of the heart. Fed. Proc 1981, 40, 2037–2041. [Google Scholar]
- Borgström, P; Lindbom, L; Arfos, KE; Intaglietta, M. Beta-adrenergic control of resistance in individual vessels in rabbit tenuissimus muscle. Am. J. Physiol 1988, 254, H631–H635. [Google Scholar]
- Bro, S; Rasmussen, RA; Handberg, J; Olgaard, K; Feldt-Rasmussen, B. Randomized crossover study comparing the phosphate-binding efficacy of calcium ketoglutarate versus calcium carbonate in patients on chronic hemodialysis. Am. J. Kidney Dis 1998, 31, 257–262. [Google Scholar]
- Brüggemann, DA; Lawson, MA. The extracellular matrix of Gadus morhua muscle contains types III, V, VI and IV collagens in addition to type I. J. Fish Biol 2005, 66, 810–821. [Google Scholar]
- Castagnola, P; Tavella, S; Gerecke, DR; Dublet, B; Gordon, MK; Seyer, J; Cancedda, R; van der Rest, M; Olson, BR. Tissue-specific expression of type XIV collagen-a member of the FACIT class of collagens. EJCB 1992, 59, 340–347. [Google Scholar]
- Clark, MG; Rattigan, S; Clerk, LH; Vincent, MA; Clark, AD; Youd, JM; Newman, JMB. Nutritive and non-nutritive blood flow: Rest and exercise. Acta Physiol. Scand 2000, 168, 519–530. [Google Scholar]
- Delp, MD; Duan, C. Composition and size of type I, IIA, IID/X, and IIB fibers and citrate synthase activity of rat muscle. J. Appl. Physiol 1996, 80, 261–270. [Google Scholar]
- Diegelmann, RF. Analysis of collagen synthesis. Methods Mol. Med 2003, 78, 349–358. [Google Scholar]
- Dodge, FA, Jr; Rahamimoff, R. Co-operative action of calcium ions in transmitter release at the neuromuscular junction. J. Physiol 1967, 193, 419–432. [Google Scholar]
- Fitts, RH. Cellular mechanisms of muscle fatigue. Physiol. Rev 1994, 74, 49–94. [Google Scholar]
- The Medical Revolution of the Seventeenth-Century; French, R; Wear, A (Eds.) Cambridge University Press: Cambridge, U.K., 1989.
- Galen on Anatomical Procedures (De Anatomicis Administrationibus); , Book VIII; Kuhn. Translated with an introduction and notes by Singer, C, Translator; Oxford University Press: London, 1956; Chapter 4pp. 667–675.
- Gelse, K; Pöschl, E; Aigner, T. Collagens-structure, function and biosynthesis. Adv. Drug Deliv. Rev 2003, 55, 1531–1546. [Google Scholar]
- Hammersen, F. The terminal vascular bed in skeletal muscle with special regard to the problem of shunts. In Capillary Permeability: The Transfer of Molecules and Ions Between Capillary Blood and Tissue; Crone, C, Lassen, NA, Eds.; Munksgaard; Copenhagen, 1970; pp. 351–365. [Google Scholar]
- Hanson, J; Huxley, HE. Structural basis of the cross-striations in muscle. Nature 1953, 172, 530–532. [Google Scholar]
- Harrison, AP; Flatman, JA. Measurement of force and both surface and deep M wave properties in isolated rat soleus muscles. Am. J. Physiol 1999, 277, R1646–R1653. [Google Scholar]
- Harrison, AP; Nielsen, AH; Eidemak, I; Molsted, S; Bartels, EM. The uremic environment and muscle dysfunction in man and rat. Nephron Physiol 2006, 103, 33–42. [Google Scholar]
- He, W; Miao, FJ; Lin, DC; Schwandner, RT; Wang, Z; Gao, J; Chen, JL; Tian, H; Ling, L. Citric acid cycle intermediates as ligands for orphan G-protein-coupled receptors. Nature 2004, 429, 188–193. [Google Scholar]
- Hess, OM; Schneider, J; Koch, R; Bamert, C; Krayenbuehl, HP. Diastolic function and myocardial structure in patients with myocardial hypertrophy. Special reference to normalized viscoelastic properties. Circulation 1983, 63, 360–371. [Google Scholar]
- Huxley, AF; Niedergerke, R. Structural changes in muscle during contraction; interference microscopy of living muscle fibres. Nature 1954, 173, 971–973. [Google Scholar]
- Icardo, JM; Colvee, E. Collagenous skeleton of the human mitral papillary muscle. Anat. Rec 1998, 252, 509–518. [Google Scholar]
- Jeevanandam, M; Petersen, SR. Substrate fuel kinetics in enterally fed trauma patients supplemented with ornithine alpha ketoglutarate. Clin. Nutr 1999, 18, 209–217. [Google Scholar]
- Ker, RF. The design of soft collagens load bearing tissues. J. Exp. Biol 1999, 202, 3315–3324. [Google Scholar]
- Koch, M; Bohrmann, M; Matthison, M; Hagios, C; Chiquet, M. Large and Small Splice Variants of Collagen XII: Differential Expression and Ligand Binding. J. Cell Biol 1995, 130, 1005–1014. [Google Scholar]
- Kubinova, L; Janacek, J; Ribaric, S; Cebasek, V; Erzen, I. Three-dimensional study of the capillary supply of skeletal muscle fibres using confocal microscopy. J. Musc. Res. Cell Motil 2001, 22, 217–227. [Google Scholar]
- Light, N; Champion, AE. Characterisation of muscle epimysium, perimysium and endomysium collagens. Biochem. J 1984, 219, 1017–1026. [Google Scholar]
- Lindbom, L; Arfos, KE. Non-homogeneous blood flow distribution in the rabbit tenuissimus muscle. Differential control of total blood flow and capillary perfusion. Acta Physiol. Scand 1984, 122, 225–233. [Google Scholar]
- Listrat, A; Lethias, C; Hocquette, JF; Renand, G; Menissier, F; Geay, Y; Picard, B. Age-related changes and location of types I, III, XII and XIV collagen during development of skeletal muscles from genetically different animals. Histochem. J 2000, 32, 349–356. [Google Scholar]
- Metzger, JM; Moss, RL. pH modulation of the kinetics of a Ca2+-sensitive cross-bridge state transition in mammalian single skeletal muscle fibres. J. Physiol 1990, 428, 751–764. [Google Scholar]
- Momose, A; Takeda, T; Itai, Y. Blood vessels: Depiction at phase-contrast X-ray imaging without contrast agents in the mouse and rat-feasibility study. Radiology 2000, 217, 593–596. [Google Scholar]
- Moukarzel, AA; Goulet, O; Salas, JS; Marti-Henneberg, C; Buchman, AL; Cynober, L; Rappaport, R; Ricour, C. Growth retardation in children receiving long-term total parenteral nutrition: Effects of ornithine alpha-ketoglutarate. Am. J. Clin. Nutr 1994, 60, 408–413. [Google Scholar]
- Mudge, GH. Studies on potassium accumulation by rabbit kidney slices: Effect of metabolic activity. Am. J. Physiol 1951, 165, 113–127. [Google Scholar]
- Myrhage, R; Eriksson, E. Vascular arrangements in hindlimb muscles of the cat. J. Anat 1980, 131, 1–17. [Google Scholar]
- Nielsen, OB; Overgaard, K. Ion gradients and contractility in skeletal muscle: The role of active Na+, K+ transport. Acta Physiol. Scand 1996, 156, 247–256. [Google Scholar]
- Purslow, PP. The structure and functional significance of variations in the connective tissue within muscle. Comp. Biochem. Physiol. A. Mol. Integr. Physiol 2002, 133, 947–966. [Google Scholar]
- Rathazer, ES; Cline, HT. Multiphotom imaging of neurons in living tissue: Acquisition and analysis of time-lapse morphological data. Real-Time Imaging 2002, 8, 175–188. [Google Scholar]
- Rowe, RWD. Morphology of perimysial and endomysial connective tissue in skeletal muscle. Tissue Cell 1981, 13, 681–690. [Google Scholar]
- Rowell, RB. Ideas about control of skeletal and cardiac muscle blood flow (1876–2003): Cycles of revision and new vision. J. Appl. Physiol 2004, 97, 384–392. [Google Scholar]
- Rutten, EP; Engelen, MP; Schols, AM; Deutz, NE. Skeletal muscle glutamate metabolism in health and disease: state of the art. Curr. Opin. Clin. Nutr. Metab. Care 2005, 8, 41–51. [Google Scholar]
- Sato, K; Ohashi, C; Muraki, M; Itsuda, H; Yokoyama, Y; Kanamori, M; Ohtsuki, K; Kawabata, M. Isolation of intact type V collagen from fish intramuscular connective tissue. J. Food Biochem 1998, 22, 213–215. [Google Scholar]
- Takeda, T; Momose, A; Wu, J; Yu, Q; Zeniya, T; Thet-Thet-Lwin; Yoneyama, AMS; Itai, Y. Vessel imaging by interferometric phase-contrast X-ray technique. Circulation 2002, 105, 1708–1712. [Google Scholar]
- Thompson, LV; Fitts, RH. Muscle fatigue in the frog semitendinous: role of the high-energy phosphate and Pi. Am. J. Physiol 1992, 263, C803–C809. [Google Scholar]
- Weon, BM; Je, J; Hwu, Y; Margaritondo, G. Phase contrast X-ray imaging. Int. J. Nanotechnology 2006, 3, 280–297. [Google Scholar]
- Winegrad, BA; Robinson, TF. Force generation among cells in the relaxing heart. Eur. J. Cardiol 1978, 7, 63–70. [Google Scholar]
- Yu-Yahiro, JA. Electrolytes and their relationship to normal and abnormal muscle function. Orthop. Nurs 1994, 13, 38–40. [Google Scholar]



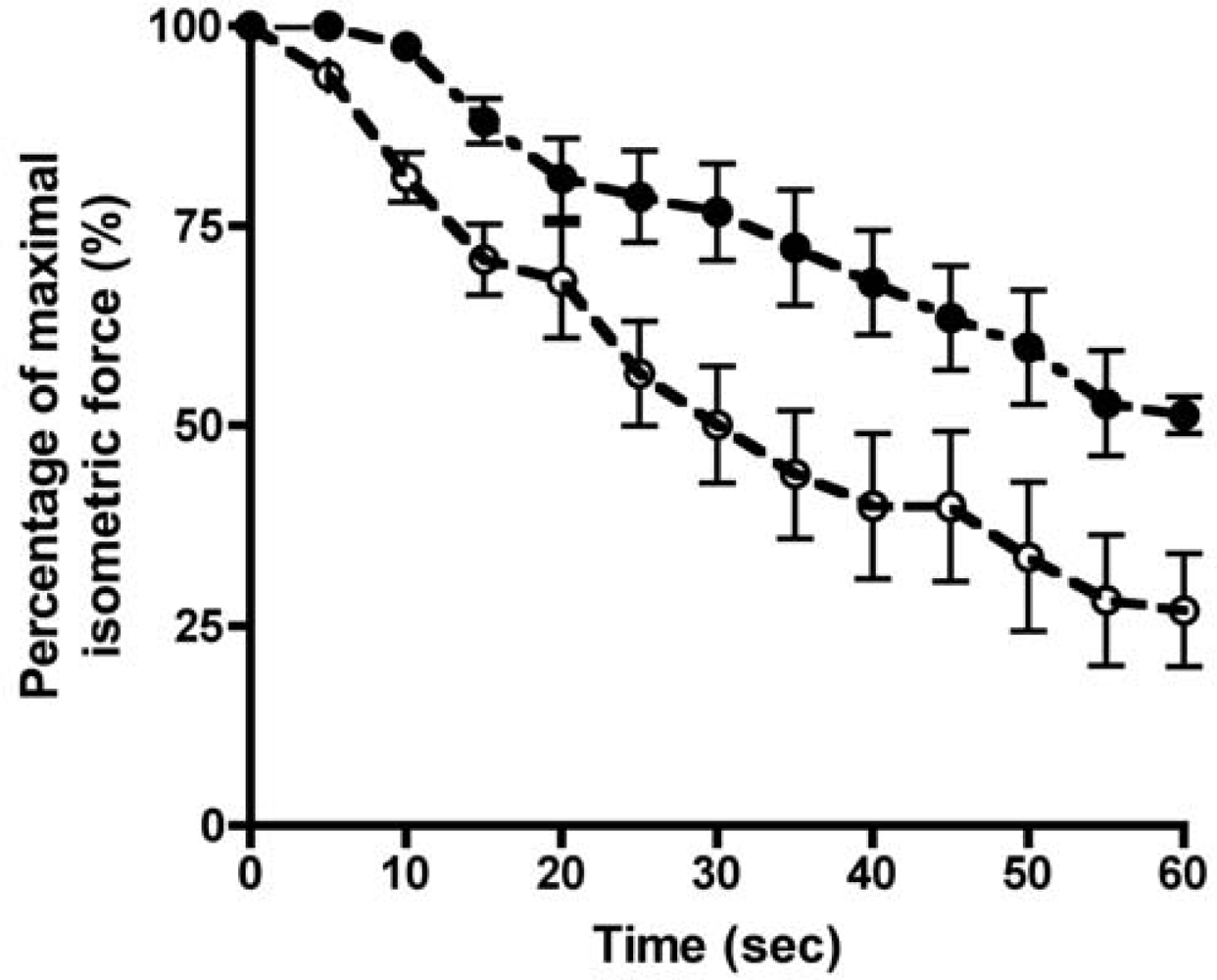
 ; n=8) or AKG treated rats (•; n=4), respectively (see materials & methods for details of the AKG treatment). Isolated muscles were continuously stimulated at 40 Hz for a period of 60 seconds with 32 mA pulses of 1 ms duration, which represents supramaximal, constant-current field stimulation. Force recordings were made via a force transducer attached to an A/D converter at a sampling rate of 1000 samples per second. Each point represents mean ± SE. Significant differences between the muscles were found using a Mann-Whitney unpaired two-tailed test (P<0.05).
; n=8) or AKG treated rats (•; n=4), respectively (see materials & methods for details of the AKG treatment). Isolated muscles were continuously stimulated at 40 Hz for a period of 60 seconds with 32 mA pulses of 1 ms duration, which represents supramaximal, constant-current field stimulation. Force recordings were made via a force transducer attached to an A/D converter at a sampling rate of 1000 samples per second. Each point represents mean ± SE. Significant differences between the muscles were found using a Mann-Whitney unpaired two-tailed test (P<0.05).
; n=8) or AKG treated rats (•; n=4), respectively (see materials & methods for details of the AKG treatment). Isolated muscles were continuously stimulated at 40 Hz for a period of 60 seconds with 32 mA pulses of 1 ms duration, which represents supramaximal, constant-current field stimulation. Force recordings were made via a force transducer attached to an A/D converter at a sampling rate of 1000 samples per second. Each point represents mean ± SE. Significant differences between the muscles were found using a Mann-Whitney unpaired two-tailed test (P<0.05).
; n=8) or AKG treated rats (•; n=4), respectively (see materials & methods for details of the AKG treatment). Isolated muscles were continuously stimulated at 40 Hz for a period of 60 seconds with 32 mA pulses of 1 ms duration, which represents supramaximal, constant-current field stimulation. Force recordings were made via a force transducer attached to an A/D converter at a sampling rate of 1000 samples per second. Each point represents mean ± SE. Significant differences between the muscles were found using a Mann-Whitney unpaired two-tailed test (P<0.05).
© 2008 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/). This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Brüggemann, D.A.; Risbo, J.; Pierzynowski, S.G.; Harrison, A.P. Muscle Contraction and Force: the Importance of an Ancillary Network, Nutrient Supply and Waste Removal. Int. J. Mol. Sci. 2008, 9, 1472-1488. https://doi.org/10.3390/ijms9081472
Brüggemann DA, Risbo J, Pierzynowski SG, Harrison AP. Muscle Contraction and Force: the Importance of an Ancillary Network, Nutrient Supply and Waste Removal. International Journal of Molecular Sciences. 2008; 9(8):1472-1488. https://doi.org/10.3390/ijms9081472
Chicago/Turabian StyleBrüggemann, Dagmar A., Jens Risbo, Stefan G. Pierzynowski, and Adrian P. Harrison. 2008. "Muscle Contraction and Force: the Importance of an Ancillary Network, Nutrient Supply and Waste Removal" International Journal of Molecular Sciences 9, no. 8: 1472-1488. https://doi.org/10.3390/ijms9081472
APA StyleBrüggemann, D. A., Risbo, J., Pierzynowski, S. G., & Harrison, A. P. (2008). Muscle Contraction and Force: the Importance of an Ancillary Network, Nutrient Supply and Waste Removal. International Journal of Molecular Sciences, 9(8), 1472-1488. https://doi.org/10.3390/ijms9081472