Aflatoxin B1 Degradation by Stenotrophomonas Maltophilia and Other Microbes Selected Using Coumarin Medium


Abstract
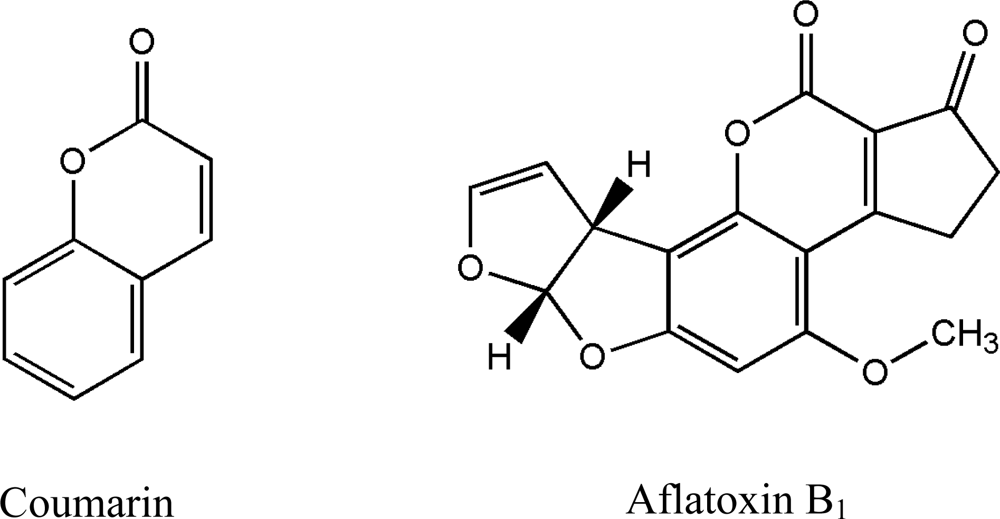
:1. Introduction
2. Results and Discussion
2.1. Screening for AFB1 degradation microbes
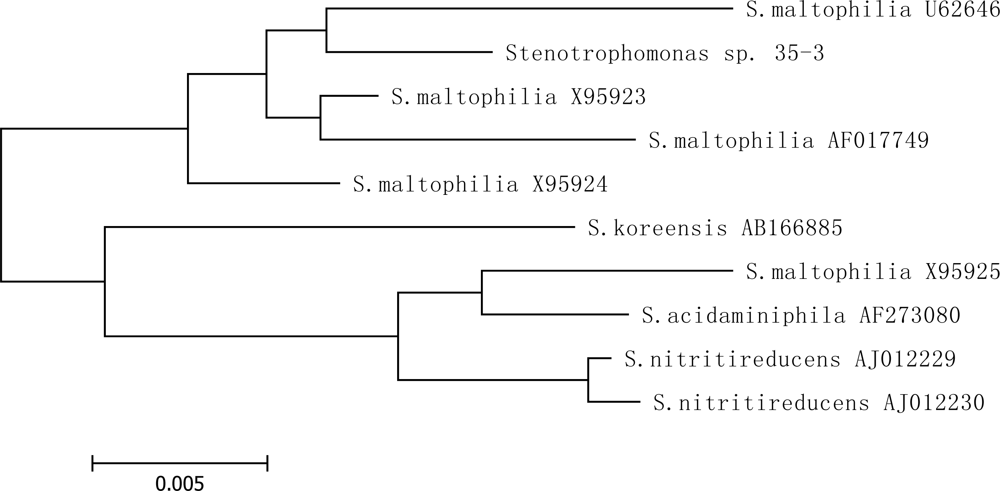
2.2. Identification of isolate 35-3
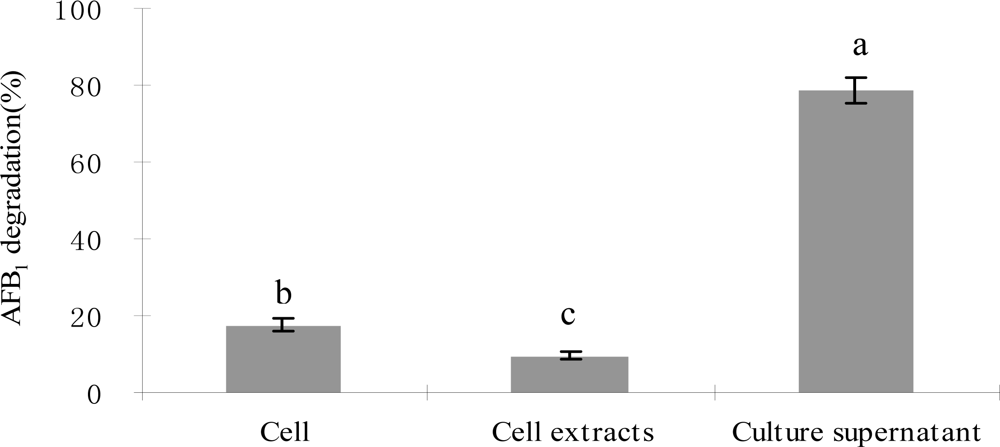
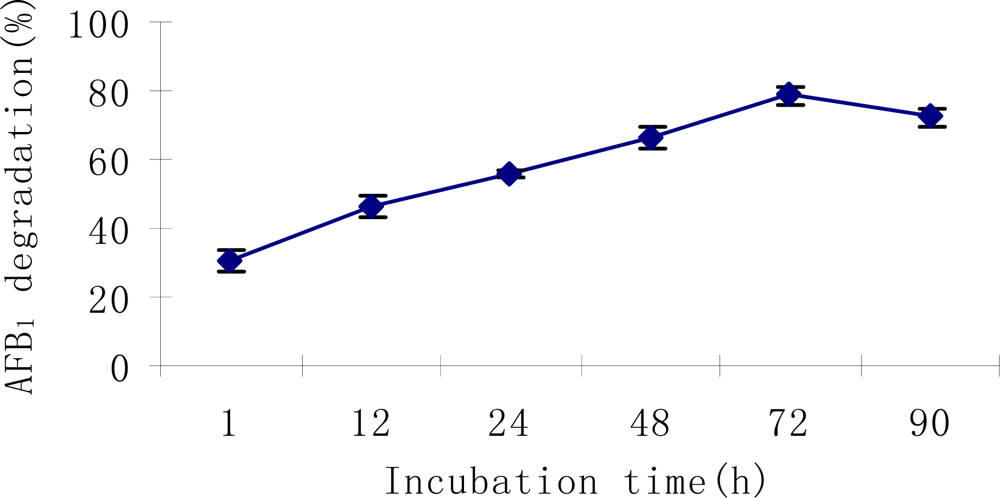
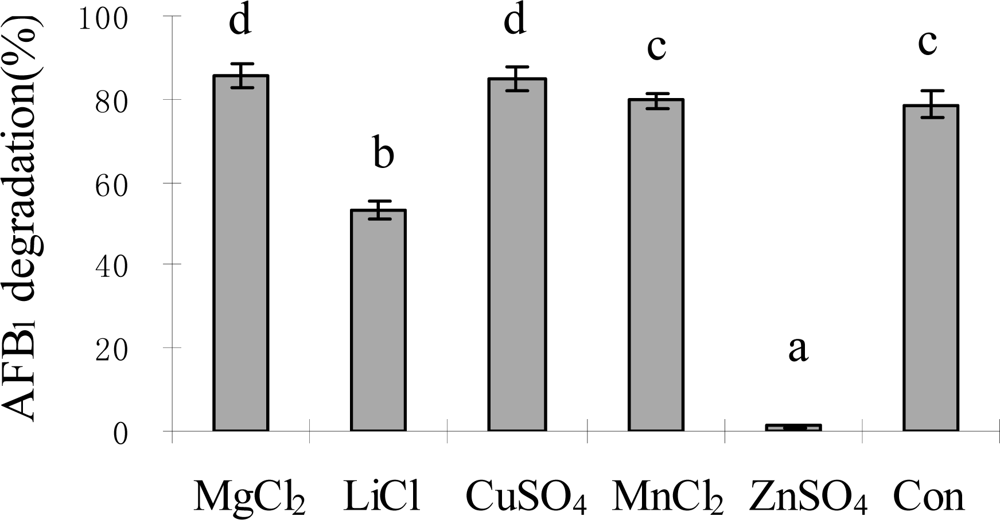
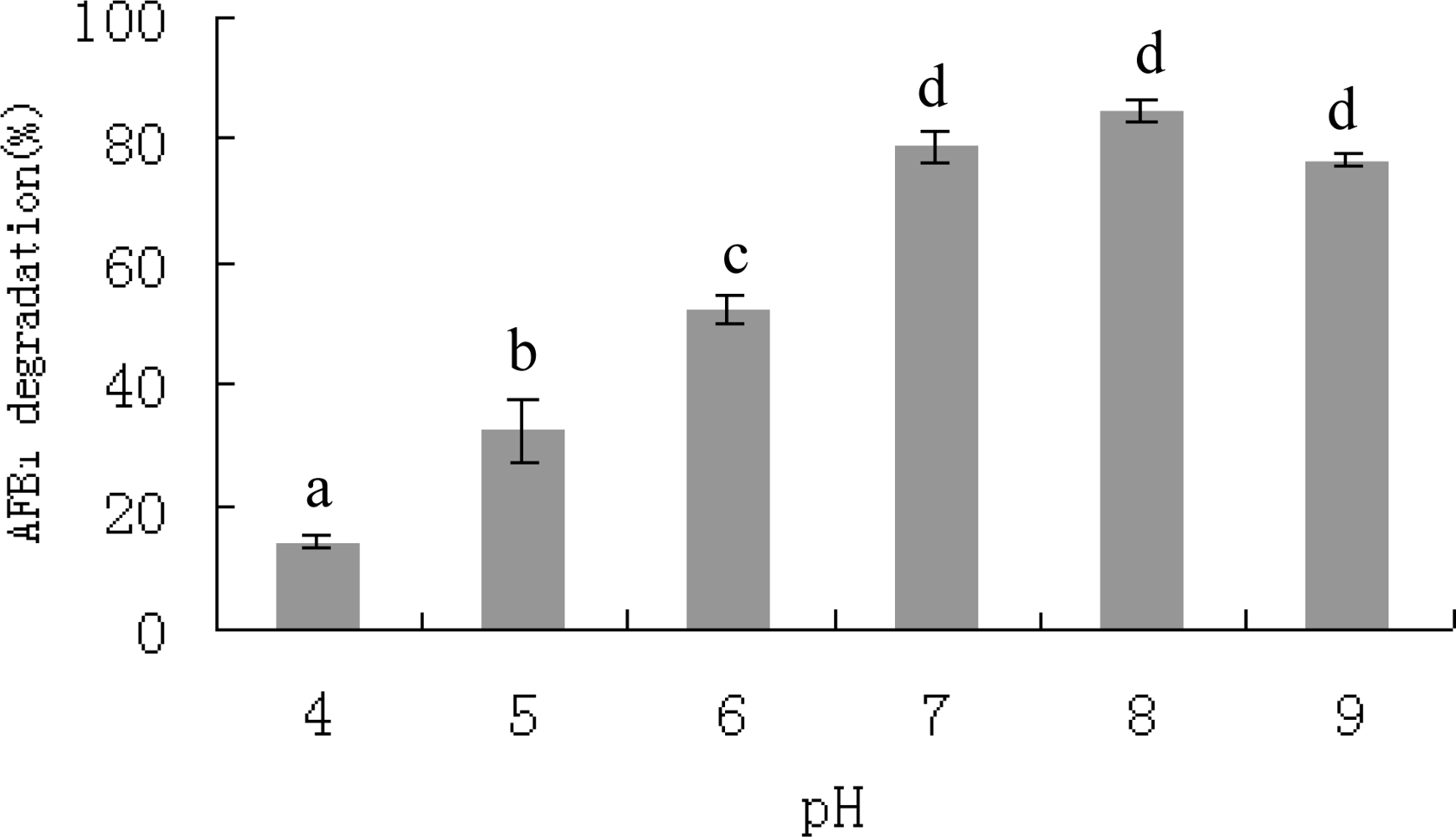
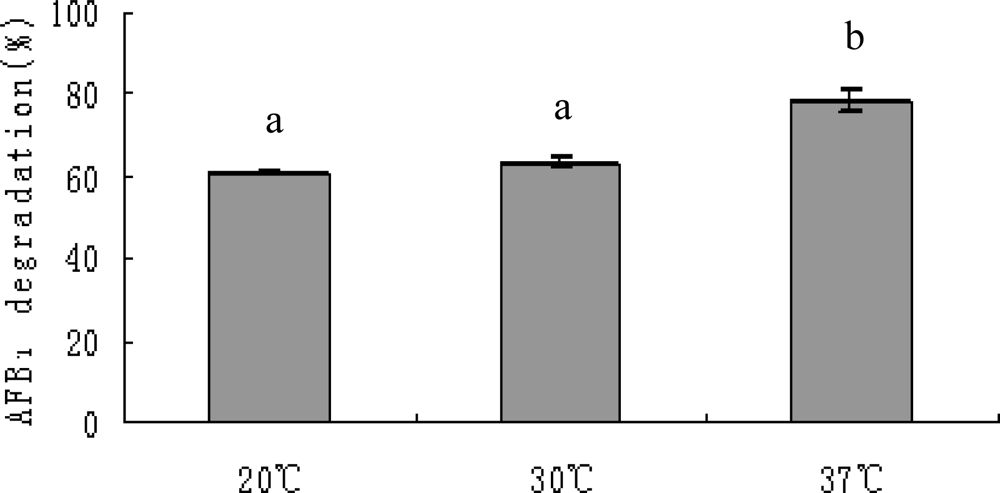
2.3. AFB1 degradation by S. maltophilia 35-3
3. Experimental Section
3.1. Culture media
3.2. Isolation of microorganisms
3.2.1. Samples
3.2.2. Isolation
3.3. Tests of AFB1 degradation
3.4. Characterization of S.maltophilia 35-3
3.4.1. Physiological and biochemical tests
3.4.2. Determination of 16S rRNA gene sequence
3.4.3. Phylogenetic analyses
3.5. Degradation of AFB1 by S. maltophilia 35-3
3.5.1. Degradation of AFB1 by S. maltophilia 35-3 cells
3.5.2. Degradation of AFB1 by S. maltophilia 35-3 intracellular cell extracts
3.5.3. Effects of incubation period, temperature, pH, metal ions and proteinase K treatment on AFB1 degradation by S. maltophilia 35-3 supernatant
3.6. Statistical analyses
4. Conclusions
Acknowledgments
- #Supported by the National Natural Science Foundation of China (Contract number: 30571353)
References
- Bhatnagar, D; Ehrlich, KC; Cleveland, TE. Molecular genetic analysis and regulation of aflatoxin biosynthesis. Appl. Microbiol. Biotechnol 2003, 61, 83–93. [Google Scholar]
- Eaton, DL; Groopman, JD. The toxicology of aflatoxins: human health, veterinary and agricultural significance; Academic Press: San Diego, CA, 1994. [Google Scholar]
- Richard, JL; Payne, GA. Mycotoxins: risks in plant, animal, and human systems. Task Force Report No. 139; Council for Agricultural Science and Technology: Ames, Iowa, 2003. [Google Scholar]
- Diaz, DE. The mycotoxin blue book; Nottingham University Press: Nottingham, England, 2005. [Google Scholar]
- Fanelli, C; Taddei, F; Nisini, PT; Jestoi, M; Ricelli, A; Visconti, A; Fabbri, AA. Use of resveratrol and BHA to control fungal growth and mycotoxin production in wheat and maize seeds. Aspect Appl. Biol 2003, 68, 63–71. [Google Scholar]
- Piva, FP; Galvano, RD; Pietri, AP; Piva, RD. Detoxification methods of aflatoxins. Nutr. Res 1995, 15, 767–776. [Google Scholar]
- Yazdanpanah, H; Mohammadi, T; Abouhossain, G; Cheraghali, AM. Effect of roasting on degradation of aflatoxins in contaminated pistachio nuts. Food Chem. Toxicol 2005, 43, 1135–1139. [Google Scholar]
- Albores, AM; Villa, GA; Pina, MGFL; Tostado, EC; Martínez, EM. Safety and efficacy evaluation of aqueous citric acid to degrade B-aflatoxins in maize. Food Chem. Toxicol 2005, 43, 233–238. [Google Scholar]
- Gowda, NKS; Suganthi, RU; Malathi, V; Raghavendra, A. Efficacy of heat treatment and sun drying of aflatoxin-contaminated feed for reducing the harmful biological effects in sheep. Animal Feed Sci. Tech 2007, 133, 167–175. [Google Scholar]
- Mishra, HN; Das, C. A review on biological control and metabolism of aflatoxin. Crit. Rev. Food Sci 2003, 43, 245–264. [Google Scholar]
- Zhou, T; He, J; Gong, J. Microbial transformation of trichothecene mycotoxins. World Mycotoxin Journal 2008, 1, 23–30. [Google Scholar]
- Motomura, M; Toyomasu, T; Mizuno, K; Shinozawa, T. Purification and characterization of an aflatoxin degradation enzyme from Pleurotus ostreatus. Microbiol. Res 2003, 158, 237–242. [Google Scholar]
- Zjalic, S; Reverberi, M; Ricelli, A; Granito, VM; Fanelli, C; Fabbri, AA. Trametes versicolor: A possible tool for aflatoxin control. Int. J. Food Microbiol 2006, 107, 243–249. [Google Scholar]
- Varga, J; Peteri, Z; Tabori, K; Teren, J; Vagvolgyi, C. Degradation of ochratoxin A and other mycotoxins by Rhizopus isolates. Int. J. Food Microbiol 2005, 99, 321–328. [Google Scholar]
- Molnar, O; Schatzmayr, G; Elisabeth, F; Prillinger, H. Trichosporon mycotoxinivorans sp. nov., A new yeast species useful in biological detoxification of various mycotoxins. System. Appl. Microbiol 2004, 27, 661–671. [Google Scholar]
- Shetty, PH; Jespersen, L. Saccharomyces cerevisiae and lactic acid bacteria as potential mycotoxin decontaminating agents. Trends Food Sci. Tech 2006, 17, 48–55. [Google Scholar]
- Shantha, T. Fungal degradation of aflatoxin B1. Nat. Toxins 1999, 7, 175–178. [Google Scholar]
- Liu, DL; Yao, DS; Liang, R; Ma, L; Cheng, WQ; Gu, LQ. Detoxification of aflatoxin B1 by enzymes isolated from Armillariella tabescens. Food Chem. Toxicol 1998, 36, 563–574. [Google Scholar]
- El-Nezami, H; Kankaanpaa, P; Salminen, S; Ahokas, J. Ability of dairy strains of lactic acid bacteria to bind a common food carcinogen, aflatoxin B1. Food Chem. Toxicol 1998, 36, 321–326. [Google Scholar]
- Gratz, S; Mykkanen, H; El-Nezami, H. Aflatoxin B1 binding by a mixture of Lactobacillus and Propionibacterium: in vitro versus ex vivo. J Food Prot 2005, 68, 2470–2474. [Google Scholar]
- Peltonen, K; El-Nezami, H; Haskard, C; Ahokas, J; Salminen, S. Aflatoxin B1 binding by dairy strains of lactic acid bacteria and bifidobacteria. J. Dairy Sci 2001, 84, 2152–2156. [Google Scholar]
- Peltonen, K; El-Nezami, H; Salminen, S; Ahokas, J. Binding of aflatoxin B1 by probiotic bacteria. J. Sci. Food Agric 2000, 80, 1942–1945. [Google Scholar]
- El-Nezami, H; Mykkanen, H; Kankaanpaa, P; Salminen, S; Ahokas, J. Ability of Lactobacillus and Propionibacterium strains to remove aflatoxin B1 from the chicken duodenum. J Food Prot 2000, 63, 549–552. [Google Scholar]
- Pierides, M; El-nezami, H; Peltonen, K; Salminen, S; Ahokas, J. Ability of dairy strains of lactic acid bacteria to bind aflatoxin M1 in a food model. J Food Prot 2000, 63, 645–650. [Google Scholar]
- Alberts, JF; Engelbrecht, Y; Steyn, PS; Holzapfel, WH; Vanzyl, WH. Biological degradation of aflatoxin B1 by Rhodococcus erythropolis cultures. Int. J. Food Microbiol 2006, 109, 121–126. [Google Scholar]
- Hormisch, D; Brost, I; Kohring, GW; Giffhorn, F; Kroppensted, RM; Stackebrandt, E; Färber, P; Holzapfel, WH. Mycobacterium fluoranthenivorans sp. nov., a fluoranthene and aflatoxin B1 degrading bacterium from contaminated soil of a former coal gas plant. System. Appl. Microbiol 2004, 27, 653–660. [Google Scholar]
- Teniola, OD; Addo, PA; Brost, IM; Farber, P; Jany, KD; Alberts, JF; Vanzyl, WH; Steyn, PS; Holzapfel, WH. Degradation of aflatoxin B1 by cell-free extracts of Rhodococcus erythropolis and Mycobacterium fluoranthenivorans sp. nov. DSM 44556T. Int. J. Food Microbiol 2005, 105, 111–117. [Google Scholar]
- D'Souza, DH; Brackett, RE. The role of trace metal ions in aflatoxin B1 degradation by Flavobacterium aurantiacum. J Food Prot 1998, 61, 1666–1669. [Google Scholar]
- D'Souza, DH; Brackett, RE. The influence of divalent cations and chelators on aflatoxin B1 degradation by Flavobacterium aurantiacum. J Food Prot 2000, 63, 102–105. [Google Scholar]
- D'Souza, DH; Brackett, RE. Aflatoxin B1 degradation by Flavobacterium aurantiacum in the presence of reducing conditions and seryl and sulfhydryl group inhibitors. J Food Prot 2001, 64, 268–271. [Google Scholar]
- Smiley, RD; Draughon, FA. Preliminary evidence that degradation of aflatoxin B1 by Flavobacterium aurantiacum is enzymatic. J. Food Prot 2000, 63, 415–418. [Google Scholar]
- Volkl, A; Vogler, B; Schollenberger, M; Karlovsky, P. Microbial detoxification of mycotoxin deoxynivalenol. J. Basic Microbiol 2004, 44, 147–156. [Google Scholar]
- Lee, LS; Dunn, JJ; De Lucca, AJ; Ciegler, A. Role of lactone ring of aflatoxin B1 in toxicity and mutagenicity. Experientia 1981, 37, 16–17. [Google Scholar]
- Bergot, BJ; Stanley, WL; Masri, MS. Reaction of coumarin with aqua ammonia. Implications in detoxification of aflatoxin. J. Agr. Food Chem 1977, 25, 965–966. [Google Scholar]
- Grove, MD; Plattner, RD; Weisleder, D. Ammoniation products of an aflatoxin model coumarin. J. Agr. Food Chem 1981, 29, 1161–1164. [Google Scholar]
- Juhasz, AL; Stanley, GA; Britz, ML. Microbial degradation and detoxification of high molecular weight polycyclic aromatic hydrocarbons by Stenotrophomonas maltophilia strain VUN 10,003. Lett. Appl. Microbiol 2000, 30, 396–401. [Google Scholar]
- Binks, PR; Nicklin, S; Bruce, NC. Degradation of hexahydo-1, 3, 5-trinitor-1, 3, 5-triazine (RDX) by Stenotrophomonas maltophilia PB1. Appl. Environ. Microbiol 1995, 61, 1318–1322. [Google Scholar]
- Kim, JD; Kang, KH. Analysis of enzymes of Stenotrophomonas maltophilia LK-24 associated with phenol degradation. Kor. J. Microbiol. Biotechnol 2004, 32, 37–46. [Google Scholar]
- Lehninger, AL; Nelson, DL; Cox, MM. Principles of biochemistry; Worth Publishers: New York, 1993. [Google Scholar]
- AOAC. Method 2003.02. Official methods of AFB1 analysis in Cattle Feed; Association of Official Analytical Chemists: Washington, DC, 2005. [Google Scholar]
- Holt, JG; Krieg, NR; Sneath, PHA. Bergey’s manual of determinative bacteriology, 9th edition; Williams & Witkins Baltimore: Maryland, USA, 1994. [Google Scholar]
- Thompson, JD; Gibson, TJ; Plewniak, F; Jeanmougin, F; Higgins, DG. The Clustal X windows interface: flexible strategies for multilple sequence alignments aided by quality analysis tools. Nucl. Acids Res 1997, 24, 4874–4882. [Google Scholar]
- Kumar, S; Tamura, K; Nei, M. Moleculor evolutionary genetic analysis version 3.1 1993.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate1 | Source | Degradation (%) ± SE2 |
|---|---|---|
| Stenotrophomonas maltophilia (35-3) | South American tapir feces | 82.50 ± 3.20a |
| Bacillus sp. | Hog deer feces | 80.93 ± 2.65ab |
| Brevundimonas sp. | Yellow cheek feces | 78.10 ± 4.48bc |
| Bacillus sp. | Farm soil | 77.80 ± 1.63bcd |
| Klebsiella sp. | Rabbit feces | 77.57 ± 4.36cd |
| Brevundimonas sp. | Goral feces | 76.83 ± 0.72cd |
| Enterobacter sp. | Hog deer feces | 75.92 ± 3.44cd |
| Brachybacterium sp. | Rabbit feces | 74.83 ± 2.47cd |
| Rhodococcus sp. | Ostrich feces | 73.92 ± 5.48cd |
| Cellulosimicrobium sp. | Farm soil | 73.75 ± 3.60d |
| 32-2 | Goral feces | 67.64 ± 1.72e |
| K2 | Deer feces | 67.64 ± 0.75e |
| 41-4 | Zebra feces | 64.81 ± 4.84e |
| K3 | Deer feces | 64.23 ± 1.44e |
| I1 | Francois monkey feces | 58.76 ± 2.48f |
| N1 | Farm soil | 51.50 ± 0.57g |
| 23-5 | Goral feces | 48.69 ± 3.18gh |
| G3 | Zebra feces | 46.39 ± 1.25h |
| 42-1 | Compound feed | 45.18 ± 1.30h |
| J1 | Red goral feces | 30.88 ± 2.82i |
| 39-3 | White cheek feces | 28.08 ± 1.25i |
| 37-1 | Leopard feces | 18.71 ± 0.87j |
| H1 | Farm soil | 13.94 ± 1.01k |
| 31-3 | Compound feed | 11.91 ± 2.01k |
| C1 | Grey leaf monkey feces | 9.18 ± 1.54k |
| Item | Result1 | Item | Result1 | Item | Result1 |
|---|---|---|---|---|---|
| Carbon utilization: | L-Glutamic acid | + | Casein | + | |
| Glucose | + | Nitrogen utilization: | Oxidase | − | |
| D(+)-Cellobiose | + | Ammonium oxalate | − | Degradation of: | |
| Sorbitol | w | (NH4)2SO4 | − | Sodium alga acid | − |
| L- Arginine | − | NH4H2PO4 | − | Cellulose | − |
| L-Phenylalanine | − | Glutamic acid | − | lignan xylan | − |
| Maltose | + | Proline | − | Lecithin | − |
| Mannitol | + | NaNO2 | − | Yeast cell | + |
| D- Fructose | + | NH4NO4 | + | Utilisation of acid: | |
| Galactose | + | Ammonium citrate | − | Citric acid | + |
| Amylum | + | Growth at: 10 °C/ 55 °C | − | Benzoic acid | + |
| D-Raffinose | + | Growth on: | Tartaric acid | + | |
| Mannose | w | 0% / 2% NaCl | + | Succinic acid | + |
| Glycine | + | 5% / 7% / 10% NaCl | − | Acetic acid | − |
| L- Cysteine | − | Hydrolysis of: | Other tests: | ||
| L-Tyrosine | − | Gelatin | + | Congo red tolerance | + |
| D- Xylose | + | Olein | − | V-P test | − |
| Sucrose | + | Tween 80 | + | Methyl red test | − |
| A-Lactose | + | Amylum | − | Methylene blue trihydrate reduction | + |
© 2008 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/). This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Guan, S.; Ji, C.; Zhou, T.; Li, J.; Ma, Q.; Niu, T. Aflatoxin B1 Degradation by Stenotrophomonas Maltophilia and Other Microbes Selected Using Coumarin Medium. Int. J. Mol. Sci. 2008, 9, 1489-1503. https://doi.org/10.3390/ijms9081489
Guan S, Ji C, Zhou T, Li J, Ma Q, Niu T. Aflatoxin B1 Degradation by Stenotrophomonas Maltophilia and Other Microbes Selected Using Coumarin Medium. International Journal of Molecular Sciences. 2008; 9(8):1489-1503. https://doi.org/10.3390/ijms9081489
Chicago/Turabian StyleGuan, Shu, Cheng Ji, Ting Zhou, Junxia Li, Qiugang Ma, and Tiangui Niu. 2008. "Aflatoxin B1 Degradation by Stenotrophomonas Maltophilia and Other Microbes Selected Using Coumarin Medium" International Journal of Molecular Sciences 9, no. 8: 1489-1503. https://doi.org/10.3390/ijms9081489
APA StyleGuan, S., Ji, C., Zhou, T., Li, J., Ma, Q., & Niu, T. (2008). Aflatoxin B1 Degradation by Stenotrophomonas Maltophilia and Other Microbes Selected Using Coumarin Medium. International Journal of Molecular Sciences, 9(8), 1489-1503. https://doi.org/10.3390/ijms9081489