Restoration of Legacy Trees as Roosting Habitat for Myotis Bats in Eastern North American Forests

Abstract
:1. Introduction
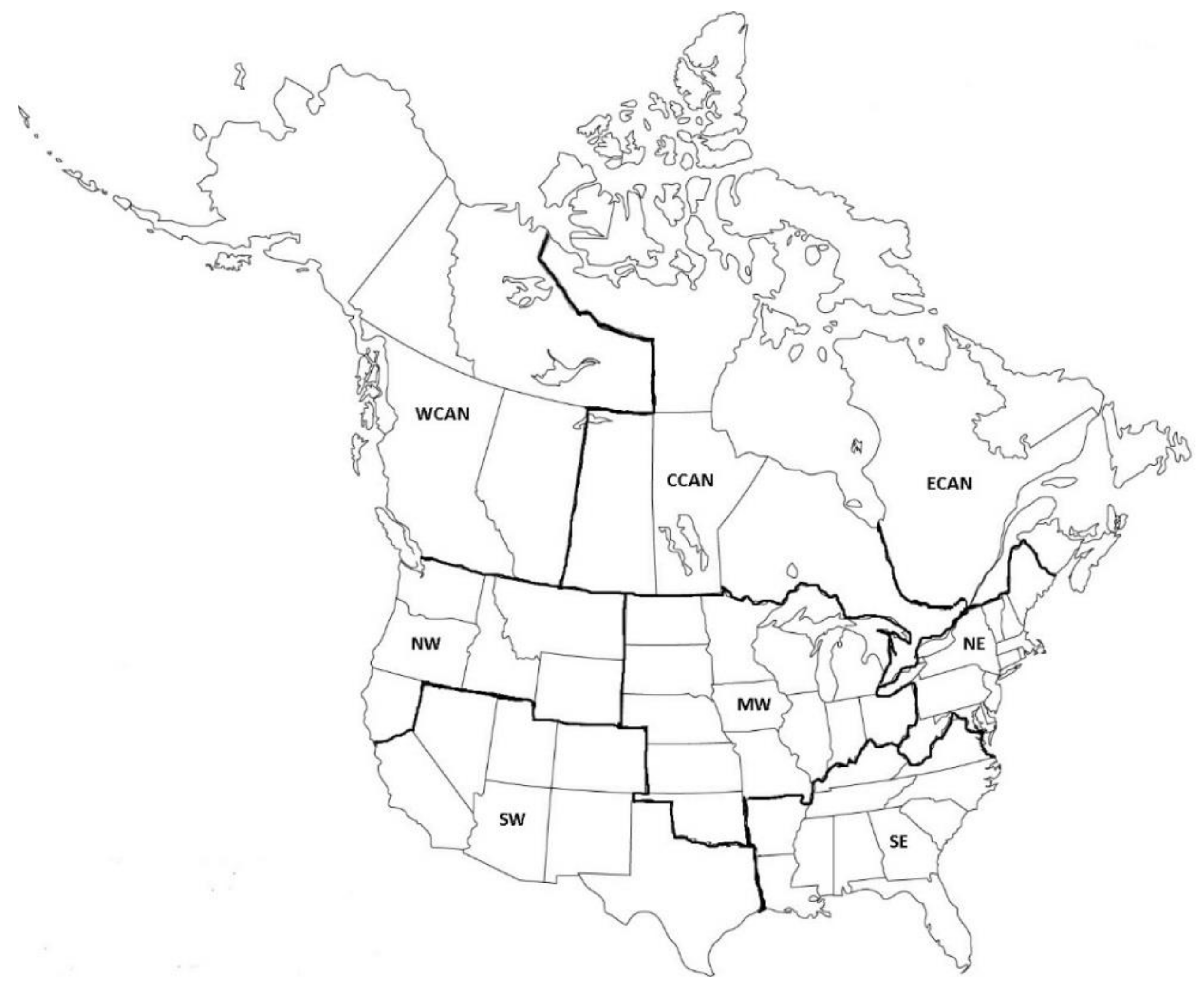
2. Materials and Methods
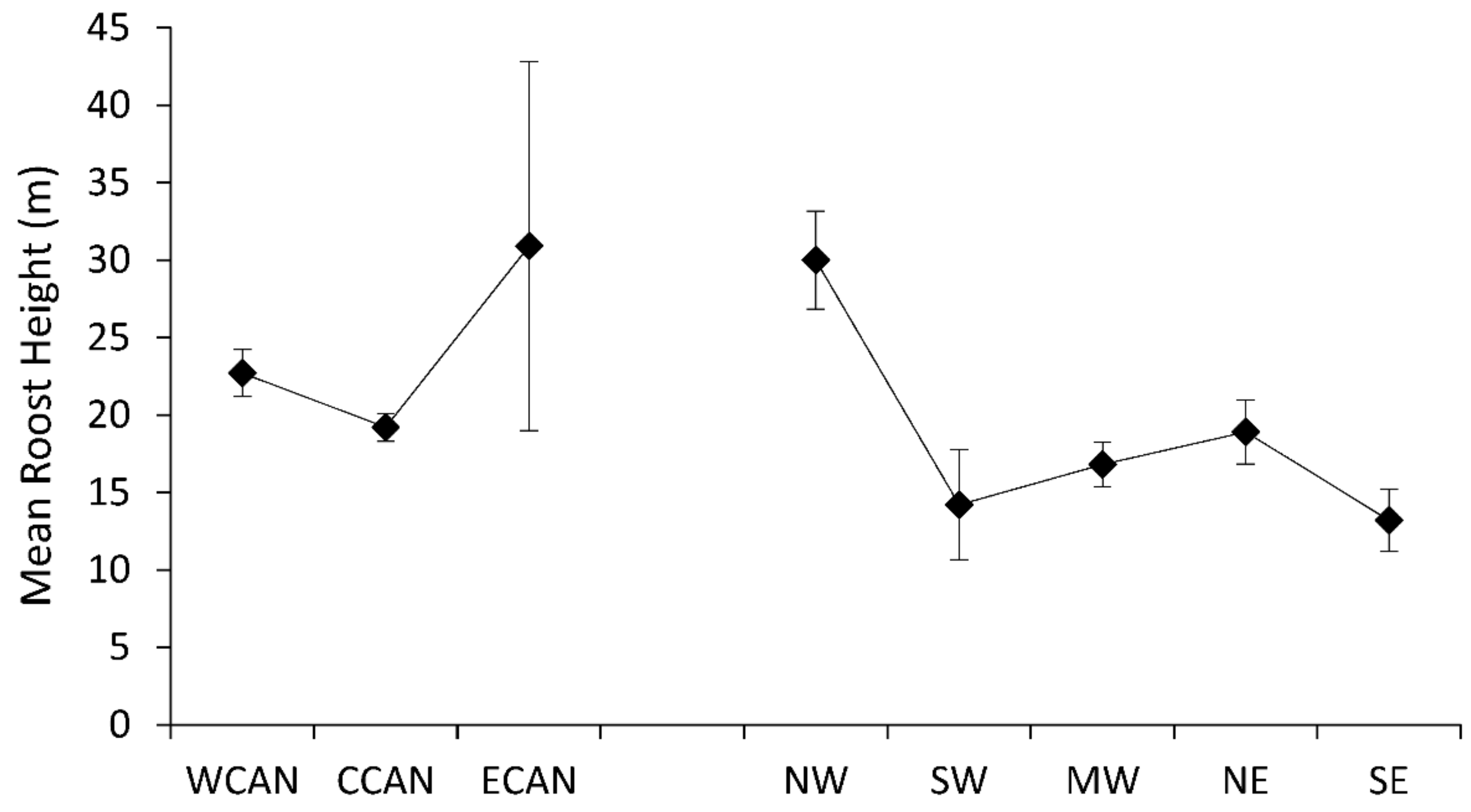
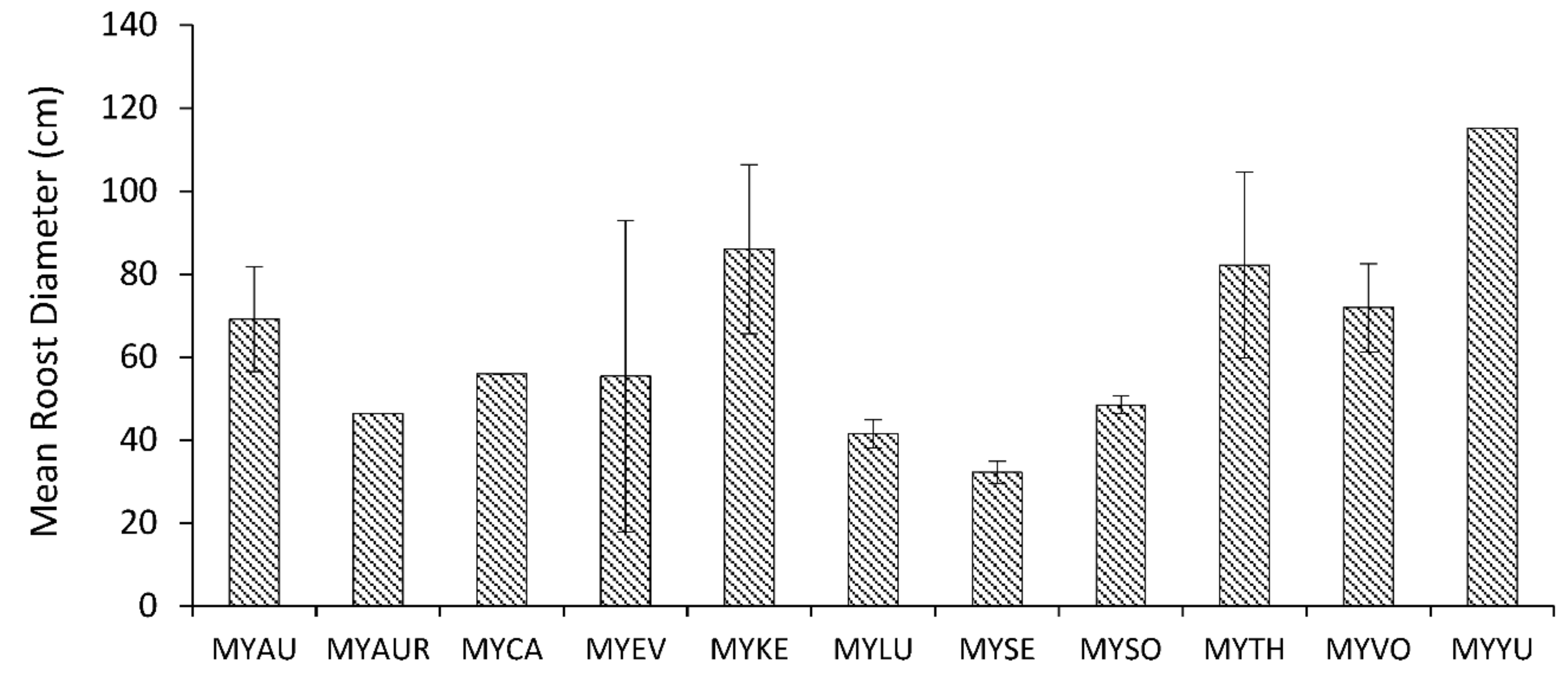
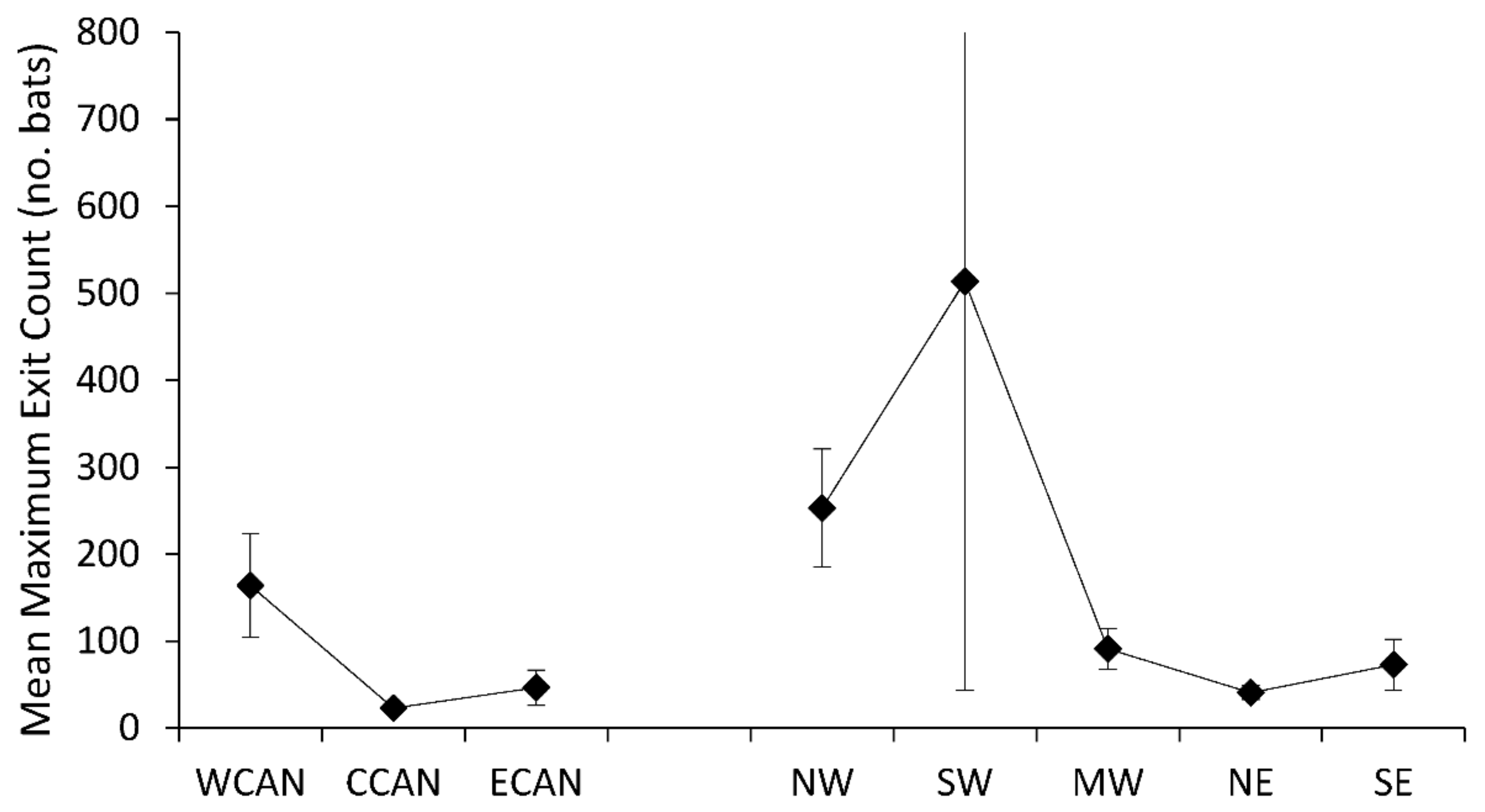
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
Appendix A. Literature Sources Used in Analyses of Roosting Preferences of Myotis in North America
| Baker, M.D.; Lacki, M.J. Day-roosting habitat of female long-legged Myotis in ponderosa pine forests. J. Wildl. Manag. 2006, 70, 207–215. |
| Bergeson, S.M.; Carter, T.C.; Whitby, M.D. Adaptive roosting gives little brown bats an advantage over endangered Indiana bats. Am. Midl. Nat. 2015, 174, 321–330. |
| Bernardos, D.A.; Chambers, C.L.; Rabe, M.J. Selection of gambel oak roosts by southwestern Myotis in ponderosa pine-dominated forests, northern Arizona. J. Wildl. Manag. 2004, 68, 595–601. |
| Boland, J.L.; Hayes, J.P.; Smith, W.P.; Huso, M.H. Selection of day-roosts by Keen’s Myotis (Myotis keenii) at multiple spatial scales. J. Mammal. 2009, 90, 222–234. |
| Brandebura, S.C.; Pannkuk, E.L.; Risch, T.S. Indiana bat (Myotis sodalis) maternity colonies in Arkansas. Southeast. Nat. 2011, 10, 529–532. |
| Brigham, R.M.; Vonhof, M.J.; Barclay, R.M.R.; Gwilliam, J.C. Roosting behavior and roost-site preferences of forest-dwelling California bats (Myotis californicus). J. Mammal. 1997, 78, 1231–1239. |
| Britzke, E.R.; Harvey, M.J.; Loeb, S.C. Indiana bat, Myotis sodalis, maternity roosts in the southern United States. Southeast. Nat. 2003, 2, 235–242. |
| Britzke, E.R.; Hicks, A.C.; von Oettingen, S.L.; Darling, S.R. Description of spring roost trees used by female Indiana bats (Myotis sodalis) in the Lake Champlain Valley of Vermont and New York. Am. Midl. Nat. 2006, 155, 181–187. |
| Broders, H.G.; Forbes, G.J. Interspecific and intersexual variation in roost-site selection of northern long-eared and little brown bats in the Greater Fundy National Park ecosystem. J. Wildl. Manag. 2004, 68, 602–610. |
| Callahan, E.V.; Drobney, R.D.; Clawson, R.L. Selection of summer roosting sites by Indiana bats (Myotis sodalis) in Missouri. J. Mammal. 1997, 78, 818–825. |
| Carter, T.C.; Feldhamer, G.A. Roost tree use by maternity colonies of Indiana bats and northern long-eared bats in southern Illinois. For. Ecol. Manag. 2005, 219, 259–268. |
| Carver, B.D.; Ashley, N. Roost tree use by sympatric Rafinesque’s big-eared bats (Corynorhinus rafinesquii) and southeastern Myotis (Myotis austroriparius). Am. Midl. Nat. 2008, 160, 364–373. |
| Clement, M.J.; Castleberry, S.B. Divergent roosting habits of Rafinesque’s big-eared bat and southeastern Myotis during winter floods. Am. Midl. Nat. 2013, 170, 158–170. |
| Cope, J.B.; Richter, A.R.; Mills, R.S. A summer concentration of the Indiana bat, Myotis sodalis, in Wayne County, Indiana. Proc. Ind. Acad. Sci. 1974, 83, 482–484. |
| Crampton, L.H.; Barclay, R.M.R. 1996. Habitat selection by bats in fragmented and unfragmented aspen mixedwood stands of different ages. In Bats and Forests Symposium; Barclay, R.M.R., Brigham, R.M., Eds.; Working Paper 23; Research Branch, British Columbia Ministry of Forests: Victoria, BC, Canada, 1996; pp. 238−259. |
| Crampton, L.H.; Barclay, R.M.R. Selection of roosting and foraging habitat by bats in different-aged aspen mixedwood stands. Conserv. Biol. 1998, 12, 1347–1358. |
| Cryan, P.M.; Bogan, M.A.; Yanega, G.M. Roosting habits of four bat species in the Black Hills of South Dakota. Acta Chiropterol. 2001, 3, 43–52. |
| Evelyn, M.J.; Stiles, D.A.; Young, R.A. Conservation of bats in suburban landscapes: Roost selection by Myotis yumanensis in a residential area in California. Biol. Conserv. 2004, 115, 463–473. |
| Fleming, H.L.; Jones, J.C.; Belant, J.L.; Richardson, D.M. Multi-scale roost site selection by Rafinesque’s big-eared bat (Corynorhinus rafinesquii) and southeastern Myotis (Myotis austroriparius) in Mississippi. Am. Midl. Nat. 2013, 169, 43–55. |
| Ford, W.M.; Owen, S.F.; Edwards, J.W.; Rodrigue, J.L. Robinia pseudoacacia (black locust) as day-roosts of male Myotis septentrionalis (northern bats) on the Fernow Experimental Forest, West Virginia. Northeast. Nat. 2006, 13, 15–24. |
| Foster, R.W.; Kurta, A. Roosting ecology of the northern bat (Myotis septentrionalis) and comparisons with the endangered Indiana bat (Myotis sodalis). J. Mammal. 1999, 80, 659–672. |
| Garroway, C.J.; Broders, H.G. Nonrandom association patterns at northern long-eared bat maternity roosts. Can. J. Zool. 2007, 85, 956–964. |
| Garroway, C.J.; Broders, H.G. Day roost characteristics of northern long-eared bats (Myotis septentrionalis) in relation to female reproductive status. Ecoscience 2008, 15, 89–93. |
| Gooding, G.; Langford, J.R. Characteristics of tree roosts of Rafinesque’s big-eared bat and southeastern bat in northeastern Louisiana. Southwest. Nat. 2004, 49, 61–67. |
| Grindal, S.D. Habitat use by bats, Myotis spp., in western Newfoundland. Can. Field-Nat. 1998, 113, 258–263. |
| Hofmann, J.E.; Gardner, J.E.; Krejca, J.K.; Garner, J.D. Summer records and a maternity roost of the southeastern Myotis (Myotis austroriparius) in Illinois. Trans. Il. State Acad. Sci. 1999, 92, 95–107. |
| Humphrey, S.R.; Richter, A.R.; Cope, J.B. Summer habitat and ecology of the endangered Indiana bat, Myotis sodalis. J. Mammal. 1977, 58, 334–346. |
| Johnson, J.B.; Gates, J.E. Observations of Indiana Myotis roosting and foraging behavior in Carroll County, Maryland. Md. Nat. 2009, 50, 11–30. |
| Johnson, J.B.; Ford, W.M.; Rodrigue, J.L.; Edwards, J.W.; Johnson, C.M. Roost selection by male Indiana Myotis following forest fires in central Appalachian hardwood forests. J. Fish Wildl. Manag. 2010, 1, 111–121. |
| Johnson, J.B.; Ford, W.M.; Edwards, J.W. Roost networks of northern Myotis (Myotis septentrionalis) in a managed landscape. For. Ecol. Manag. 2012, 266, 223–231. |
| Jung, T.S., Thompson, I.D.; Titman, R.D. Roost site selection by forest-dwelling male Myotis in central Ontario, Canada. For. Ecol. Manag. 2004, 202, 325–335. |
| Kalcounis, M.C.; Hecker, K.R. Intraspecific variation in roost-site selection by little brown bats (Myotis lucifugus). In Bats and Forests Symposium; Barclay, R.M.R., Brigham, R.M., Eds.; Working Paper 23; Research Branch, British Columbia Ministry of Forests: Victoria, BC, Canada, 1996; pp. 81–90. |
| Kiser, J.D.; MacGregor, J.R.; Bryan, H.D.; Howard, A. Use of concrete bridges as nightroosts. In The Indiana Bat: Biology and Management of An Endangered Species; Kurta, A., Kennedy, J., Eds.; Bat Conservation International, Inc.: Austin, TX, USA, 2002; pp. 208–215. |
| Kurta, A.; King, D.; Teramino, J.A.; Stribley, J.M.; Williams, K.J. Summer roosts of the endangered Indiana bat (Myotis sodalis) on the northern edge of its range. Am. Midl. Nat. 1993, 129, 132–138. |
| Kurta, A.; Kath, J.; Smith, E.L.; Foster, R.; Orick, M.W.; Ross, R. A maternity roost of the endangered Indiana bat (Myotis sodalis) in an unshaded, hollow, sycamore tree (Platanus occidentalis). Am. Midl. Nat. 1993b, 130, 405–407. |
| Kurta, A.; Williams, K.J.; Mies, R. Ecological, behavioural, and thermal observations of a peripheral population of Indiana bats (Myotis sodalis). In Bats and Forests Symposium; Barclay, R.M.R., Brigham, R.M., Eds.; Working Paper 23; Research Branch, British Columbia Ministry of Forests: Victoria, BC, Canada, 1996; pp. 102–117. |
| Kurta, A.; Murray, S.W.; Miller, D.H. Roost selection and movements across the summer landscape. In The Indiana Bat: Biology and Management of an Endangered Species; Kurta, A., Kennedy, J., Eds.; Bat Conservation International, Inc.: Austin, TX, USA, 2002; pp. 118–129. |
| Lacki, M.J.; Baker, M.D. Day roosts of female fringed Myotis (Myotis thysanodes) in xeric forests of the Pacific Northwest. J. Mammal. 2007, 88, 967–973. |
| Lacki, M.J.; Schwierjohann, J.H. Day-roost characteristics of northern bats in mixed mesophytic forest. J. Wildl. Manag. 2001, 65, 482–488. |
| Lacki, M.J.; Cox, D.R.; Dodd, L.E.; Dickinson, M.B. Response of northern bats (Myotis septentrionalis) to prescribed fires in eastern Kentucky forests. J. Mammal. 2009, 90, 1165–1175. |
| Lacki, M.J.; Baker, M.D.; Johnson, J.S. Geographic variation in roost-site selection of long-legged Myotis in the Pacific Northwest. J. Wildl. Manag. 2010, 74, 1218–1228. |
| Menzel, M.A.; Owen, S.F.; Ford, W.M.; Edwards, J.W.; Wood, P.B.; Chapman, B.R.; Miller, K.V. Roost tree selection by northern long-eared bat (Myotis septentrionalis) maternity colonies in an industrial forest of the central Appalachian mountains. For. Ecol. Manag. 2002, 155, 107–114. |
| Olson, C.R.; Barclay, R.M.R. Concurrent changes in group size and roost use by reproductive female little brown bats (Myotis lucifugus). Can. J. Zool. 2013, 91, 149–155. |
| Ormsbee, P.C. Characteristics, use, and distribution of day roosts selected by female Myotis volans (long-legged Myotis) in forested habitat of the Central Oregon Cascades. In Bats and Forests Symposium, Barclay, R.M.R., Brigham, R.M., Eds.; Working Paper 23; Research Branch, British Columbia Ministry of Forests: Victoria, BC, Canada, 1996; pp. 124–131. |
| Ormsbee, P.C.; McComb, W.C. Selection of day roosts by female long-legged Myotis in the central Oregon Cascade Range. J. Wildl. Manag. 1998, 62, 596–603. |
| Parsons, S.; Lewis, K.J.; Psyllakis, J.M. Relationships between roosting habitat of bats and decay of aspen in the sub-boreal forests of British Columbia. For. Ecol. Manag. 2003, 177, 559–570. |
| Perry, R.W.; Thill, R.E. Roost selection by male and female northern long-eared bats in a pine-dominated landscape. For. Ecol. Manag. 2007, 247, 220–226. |
| Psyllakis, J.M.; Brigham, R.M. Characteristics of diurnal roosts used by female Myotis bats in sub-boreal forests. For. Ecol. Manag. 2006, 223, 93–102. |
| Rabe, M.J.; Morrell, T.E.; Green, H.; deVos, J.C., Jr.; Miller, C.R. Characteristics of ponderosa pine snag roosts used by reproductive bats in northern Arizona. J. Wildl. Manag. 1998, 62, 612–621. |
| Rancourt, S.J.; Rule, M.I.; O’Connell, M.A. Maternity roost site selection of long-eared Myotis, Myotis evotis. J. Mammal. 2005, 86, 77–84. |
| Sasse, D.B.; Pekins, P.J. Summer roosting ecology of northern long-eared bats (Myotis septentrionalis) in the White Mountain National Forest. In Bats and Forests Symposium; Barclay, R.M.R., Brigham, R.M., Eds.; Working Paper 23; Research Branch, British Columbia Ministry of Forests, Victoria, BC, Canada, 1996; pp. 91–101. |
| Silvis, A.; Ford, W.M.; Britzke, E.R.; Beane, N.R.; Johnson, J.B. Forest succession and maternity day roost selection by Myotis septentrionalis in a mesophytic hardwood forest. Inter. J. For. Res. 2012, doi:10.1155/2012/148106. |
| Silvis, A.; Kniowski, A.B.; Gehrt, S.D.; Ford, W.M. Roosting and foraging social structure of the endangered Indiana bat (Myotis sodalis). PLoS ONE 2014, 9, e96937, doi:10.1371/journal.pone.0096937. |
| Silvis, A.; Ford, W.M.; Britzke, E.R.; Johnson, J.B. Association, roost use and simulated disruption of Myotis septentrionalis maternity colonies. Behav. Processes 2014, 103, 283–290. |
| Silvis, A.; Ford, W.M.; Britzke, E.R. Effects of hierarchical roost removal on northern long-eared bat (Myotis septentrionalis) maternity colonies. PLoS ONE 2015, 10, e0116356, doi:10.1371/journal.pone.0116356. |
| Timpone, J.C.; Boyles, J.G.; Murray, K.L.; Aubrey, D.P.; Robbins, L.W. Overlap in roosting habits of Indiana bats (Myotis sodalis) and northern bats (Myotis septentrionalis). Am. Midl. Nat. 2010, 163, 115–123. |
| Veilleux, J.P.; Whitaker, J.O., Jr.; Vincent, E.A. Mammals of the Newport Chemical Depot, Vermillion County, Indiana. Proc. Ind. Acad. Sci. 1998, 107, 91–104. |
| Vonhof, M.J.; Gwilliam, J.C. Intra- and interspecific patterns of day roost selection by three species of forest-dwelling bats in southern British Columbia. For. Ecol. Manag. 2007, 252, 165–175. |
| Waldien, D.L.; Hayes, J.P.; Arnett, E.B. Day-roosts of female long-eared Myotis in western Oregon. J. Wildl. Manag. 2000, 64, 785–796. |
| Watrous, K.S.; Donovan, T.M.; Mickey, R.M.; Darling, S.R.; Hicks, A.C.; von Oettingen, S.L. Predicting minimum habitat characteristics for the Indiana bat in the Champlain Valley. J. Wildl. Manag. 2006, 70, 1228–1237. |
| Weller, T.J.; Zabel, C.J. Characteristics of fringed Myotis day roosts in northern California. J. Wildl. Manag. 2001, 65, 489–497. |
| Whitaker, J.O., Jr.; Brack, V., Jr. Distribution and summer ecology in Indiana. In The Indiana Bat: Biology and Management of an Endangered Species; Kurta, A., Kennedy, J., Eds.; Bat Conservation International, Inc.: Austin, TX, USA, 2002; pp. 48–54. |
| Whitaker, J.O., Jr.; Sparks, D.W. Roosts of Indiana bats (Myotis sodalis) near the Indianapolis International Airport (1997–2001). Proc. Ind. Acad. Sci. 2008, 117, 193–202. |
References
- Holmes, M.A.; Matlack, G.R. Agricultural history drives structure and tree species composition of second growth forest over 100 years in southeastern Ohio, USA. J. Veg. Sci. 2017, 28, 736–746. [Google Scholar] [CrossRef]
- Thompson, J.R.; Carpenter, D.N.; Cogbill, C.V.; Foster, D.R. Four centuries of change in northeastern United States forests. PLoS ONE 2013, 8, e72540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varner, J.M.; Arthur, M.A.; Clark, S.L.; Dey, D.C.; Hart, J.L.; Schweitzer, C.J. Fire in eastern North American oak ecosystems: Filling the gaps. Fire Ecol. 2016, 12, 1–6. [Google Scholar] [CrossRef]
- Nowacki, G.J.; Abrams, M.D. The demise of fire and “mesophication” of forests in the eastern United States. BioScience 2008, 58, 123–138. [Google Scholar] [CrossRef]
- Morin, R.S.; Liebhold, A.M. Invasive forest defoliator contributed to the impending downward trend of oak dominance in eastern North America. Forestry 2016, 89, 284–289. [Google Scholar] [CrossRef]
- Pautasso, M.; Schlegel, M.; Holdenrieder, O. Forest health in a changing world. Microb. Ecol. 2015, 69, 826–842. [Google Scholar] [CrossRef] [PubMed]
- Betts, B. Roosts used by maternity colonies of silver-haired bats in northeastern Oregon. J. Mammal. 1998, 79, 643–650. [Google Scholar] [CrossRef]
- Mazurek, M.J.; Zielinski, W.J. Individual legacy trees influence vertebrate wildlife diversity in commercial forests. For. Ecol. Manag. 2004, 193, 321–334. [Google Scholar] [CrossRef]
- Baker, M.D.; Lacki, M.J. Day-roosting habitat of female long-legged myotis in ponderosa pine forests. J. Wildl. Manag. 2006, 70, 207–215. [Google Scholar] [CrossRef]
- Jung, T.S.; Thompson, I.D.; Titman, R.D.; Applejohn, A.P. Habitat selection by forest bats in relation to mixed-wood stand types and structure in central Ontario. J. Wildl. Manag. 1999, 63, 1306–1319. [Google Scholar] [CrossRef]
- Campbell, L.A.; Hallett, J.G.; O’Connell, M.A. Conservation of bats in managed forests: Use of roosts by Lasionycteris noctivagans. J. Mammal. 1996, 77, 976–984. [Google Scholar] [CrossRef]
- Lacki, M.J.; Baker, M.D.; Johnson, J.S. Temporal dynamics of roost snags of long-legged myotis in the Pacific Northwest, USA. J. Wildl. Manag. 2012, 76, 1310–1316. [Google Scholar] [CrossRef]
- Barclay, R.M.R.; Kurta, A. Ecology and behavior of bats roosting in tree cavities and under bark. In Bats in Forests: Conservation and Management; Lacki, M.J., Hayes, J.P., Kurta, A., Eds.; Johns Hopkins University Press: Baltimore, MD, USA, 2007; pp. 17–59. [Google Scholar]
- Kerth, G.; König, B. Fission, fusion and nonrandom associations in female Bechstein’s bats (Myotis bechsteinii). Behaviour 1999, 136, 1187–1202. [Google Scholar] [CrossRef]
- Willis, C.K.R.; Brigham, R.M. Roost switching, roost sharing and social cohesion: Forest-dwelling big brown bats, Eptesicus fuscus, conform to the fission-fusion model. Anim. Behav. 2004, 68, 495–505. [Google Scholar] [CrossRef]
- Garroway, C.J.; Broders, H.G. Nonrandom association patterns at northern long-eared bat maternity roosts. Can. J. Zool. 2007, 85, 956–964. [Google Scholar] [CrossRef]
- Patriquin, K.J.; Leonard, M.L.; Broders, H.G.; Garroway, C.J. Do social networks of female northern long-eared bats vary with reproductive period and age? Behav. Ecol. Sociobiol. 2010, 64, 899–913. [Google Scholar] [CrossRef]
- Silvis, A.; Kniowski, A.B.; Gehrt, S.D.; Ford, W.M. Roosting and foraging social structure of the endangered Indiana bat (Myotis sodalis). PLoS ONE 2014, 9, e96937. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.B.; Ford, W.M.; Edwards, J.W. Roost networks of northern myotis (Myotis septentrionalis) in a managed landscape. For. Ecol. Manag. 2012, 266, 223–231. [Google Scholar] [CrossRef]
- Johnson, J.S.; Kropczynski, J.N.; Lacki, M.J.; Langlois, G.D. Social networks of Rafinesque’s big-eared bats (Corynorhinus rafinesquii) in bottomland hardwood forests. J. Mammal. 2012, 93, 1545–1558. [Google Scholar] [CrossRef]
- Blehert, D.S.; Hicks, A.C.; Behr, M.; Meteyer, C.U.; Berlowski-Zier, B.M.; Buckles, E.L.; Coleman, J.T.; Darling, S.R.; Gargas, A.; Niver, R.; et al. Bat white-nose syndrome: An emerging fungal pathogen? Science 2009, 323, 227. [Google Scholar] [CrossRef] [PubMed]
- Gargas, A.; Trest, M.T.; Christensen, M.; Volk, T.J.; Blehert, D.S. Geomyces destructans sp. nov associated with bat white-nose syndrome. Mycotaxon 2009, 108, 147–154. [Google Scholar] [CrossRef]
- Frick, W.F.; Puechmaille, S.J.; Hoyt, J.R.; Nickel, B.A.; Langwig, K.E.; Foster, J.T.; Barlow, K.E.; Bartonička, T.; Feller, D.; Haarsma, A.J. Disease alters macroecological patterns of North American bats. Glob. Ecol. Biogeogr. 2015, 24, 741–749. [Google Scholar] [CrossRef]
- Turner, G.G.; Reeder, D.M.; Coleman, J.T.H. A five-year assessment of mortality and geographic spread of white-nose syndrome in North American bats and a look to the future. Bat Res. News 2011, 52, 13–27. [Google Scholar]
- Frick, W.F.; Pollock, J.F.; Hicks, A.C.; Langwig, K.E.; Reynolds, D.S.; Turner, G.G.; Butchkoski, C.M.; Kunz, T.H. An emerging disease causes regional population collapse of a common North American bat species. Science 2010, 329, 679–682. [Google Scholar] [CrossRef] [PubMed]
- Thogmartin, W.E.; King, R.A.; McKann, P.C.; Szymanski, J.A.; Pruitt, L. Population-level impact of white-nose syndrome on the endangered Indiana bat. J. Mammal. 2012, 93, 1086–1098. [Google Scholar] [CrossRef]
- Thogmartin, W.E.; Sanders-Reed, C.A.; Szymanski, J.A.; McKann, P.C.; Pruitt, L.; King, R.A.; Runge, M.C.; Russell, R.E. White-nose syndrome is likely to extirpate the endangered Indiana bat over large parts of its range. Biol. Conserv. 2013, 160, 162–172. [Google Scholar] [CrossRef]
- Moore, M.S.; Field, K.A.; Behr, M.J.; Turner, G.G.; Furze, M.E.; Stern, D.W.F.; Allegra, P.R.; Bouboulis, S.A.; Musante, C.D.; Vodzak, M.E.; et al. Energy conserving thermoregulatory patterns and lower disease severity in a bat resistant to the impacts of white-nose syndrome. J. Comp. Physiol. B 2017. [Google Scholar] [CrossRef] [PubMed]
- Verant, M.L.; Meteyer, C.U.; Speakman, J.R.; Cryan, P.M.; Lorch, J.M.; Blehert, D.S. White-nose syndrome initiates a cascade of physiologic disturbances in the hibernating bat host. BMC Physiol. 2014, 14, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Meteyer, C.U.; Barber, D.; Mandl, J.N. Pathology in euthermic bats with white nose syndrome suggests a natural manifestation of immune reconstitution inflammatory syndrome. Virulence 2012, 3, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Micalizzi, E.W.; Mack, J.N.; White, G.P.; Avis, T.J.; Smith, M.L. Microbial inhibitors of the fungus Pseudogymnoascus destructans, the causal agent of white-nose syndrome in bats. PLoS ONE 2017, 12, e0179770. [Google Scholar] [CrossRef] [PubMed]
- Lacki, M.J.; Dodd, L.E.; Toomey, R.S.; Thomas, S.C.; Couch, Z.L.; Nichols, B.S. Temporal changes in body mass and body condition of cave-hibernating bats during staging and swarming. J. Fish Wildl. Manag. 2015, 6, 360–370. [Google Scholar] [CrossRef]
- Barclay, R.M.R.; Brigham, R.M. (Eds.) Bats and Forests Symposium; Research Branch, British Columbia Ministry of Forests: Victoria, BC, Canada, 1996; p. 292.
- Kurta, A.; Kennedy, J. The Indiana Bat: Biology and Management of an Endangered Species; Bat Conservation International: Austin, TX, USA, 2002; p. 253. [Google Scholar]
- U.S. Department of the Interior. Final register for endangered and threatened wildlife and plants: Threatened species status for the northern long-eared bat with 4(d) rule. Fed. Regist. 2015, 80, 17973–18033. [Google Scholar]
- Kiser, J.D.; MacGregor, J.R.; Bryan, H.D.; Howard, A. Use of concrete bridges as night roosts. In The Indiana Bat: Biology and Management of an Endangered Species; Kurta, A., Kennedy, J., Eds.; Bat Conservation International, Inc.: Austin, TX, USA, 2002; pp. 208–215. [Google Scholar]
- Fleming, H.L.; Jones, J.C.; Belant, J.L.; Richardson, D.M. Multi-scale roost site selection by Rafinesque’s big-eared bat (Corynorhinus rafinesquii) and southeastern myotis (Myotis austroriparius) in Mississippi. Am. Midl. Nat. 2013, 169, 43–55. [Google Scholar] [CrossRef]
- Ormsbee, P.C. Characteristics, use, and distribution of day roosts selected by female Myotis volans (long-legged myotis) in forested habitat of the Central Oregon Cascades. In Bats and Forests Symposium; Barclay, R.M.R., Brigham, R.M., Eds.; Working Paper 23; Research Branch, British Columbia Ministry of Forests: Victoria, BC, Canada, 1996; pp. 124–131. [Google Scholar]
- Ormsbee, P.C.; McComb, W.C. Selection of day roosts by female long-legged Myotis in the central Oregon Cascade Range. J. Wildl. Manag. 1998, 62, 596–603. [Google Scholar] [CrossRef]
- Rabe, M.J.; Morrell, T.E.; Green, H.; deVos, J.C., Jr.; Miller, C.R. Characteristics of ponderosa pine snag roosts used by reproductive bats in northern Arizona. J. Wildl. Manag. 1998, 62, 612–621. [Google Scholar] [CrossRef]
- Psyllakis, J.M.; Brigham, R.M. Characteristics of diurnal roosts used by female Myotis bats in sub-boreal forests. For. Ecol. Manag. 2006, 223, 93–102. [Google Scholar] [CrossRef]
- Olson, C.R.; Barclay, R.M.R. Concurrent changes in group size and roost use by reproductive female little brown bats (Myotis lucifugus). Can. J. Zool. 2013, 91, 149–155. [Google Scholar] [CrossRef]
- Foster, R.W.; Kurta, A. Roosting ecology of the northern bat (Myotis septentrionalis) and comparisons with the endangered Indiana bat (Myotis sodalis). J. Mammal. 1999, 80, 659–672. [Google Scholar] [CrossRef]
- Barclay, R.M.R.; Brigham, R.M. Year-to-year reuse of tree-roosts by California bats (Myotis californicus) in southern British Columbia. Am. Midl. Nat. 2001, 146, 80–85. [Google Scholar] [CrossRef]
- Lučan, R.K.; Hanák, V.; Horáček, I. Long-term re-use of tree roosts by European forest bats. For. Ecol. Manag. 2009, 258, 1301–1306. [Google Scholar] [CrossRef]
- Angers, V.A.; Drapeau, P.; Bergeron, Y. Snag degradation pathways of four North American boreal tree species. For. Ecol. Manag. 2010, 259, 246–256. [Google Scholar] [CrossRef]
- Everett, R.; Lehmkuhl, J.; Schellhaas, R.; Ohlson, P.; Keenum, D.; Riesterer, H.; Spurbeck, D. Snag dynamics in a chronosequence of 26 wildfires on the east slope of the cascade range in Washington state, USA. Int. J. Wildland Fire 1999, 9, 223–234. [Google Scholar] [CrossRef]
- DeLong, S.C.; Sutherland, G.D.; Daniels, L.D.; Heemskerk, B.H.; Storaunet, K.O. Temporal dynamics of snags and development of snag habitats in wet spruce−fir stands in east-central British Columbia. For. Ecol. Manag. 2008, 255, 3613–3620. [Google Scholar] [CrossRef]
- Klingsporn, S.; Webster, C.R.; Bump, J.K. Influence of legacy-tree retention on group-selection opening persistence. For. Ecol. Manag. 2012, 286, 121–128. [Google Scholar] [CrossRef]
- Borkin, K.M.; O’Donnell, C.; Parsons, S. Bat colony size reduction coincides with clear-fell harvest operations and high rates of roost loss in plantation forest. Biodivers. Conserv. 2011, 20, 3537–3548. [Google Scholar] [CrossRef]
- Chaverri, G.; Kunz, T.H. Response of a specialist bat to the loss of a critical resource. PLoS ONE 2011, 6, e28821. [Google Scholar] [CrossRef] [PubMed]
- Neilson, A.L.; Fenton, M.B. Responses of little brown myotis to exclusion and to bat houses. Wildl. Soc. Bull. 1994, 22, 8–14. [Google Scholar]
- Kerth, G. Causes and consequences of sociality in bats. BioScience 2008, 58, 737–746. [Google Scholar] [CrossRef]
- Johnson, J.S.; Kropczynski, J.N.; Lacki, M.J. Social network analysis and the study of sociality in bats. Acta Chiropterol. 2013, 15, 1–17. [Google Scholar] [CrossRef]
- Silvis, A.; Ford, W.M.; Britzke, E.R. Effects of hierarchical roost removal on northern long-eared bat (Myotis septentrionalis) maternity colonies. PLoS ONE 2015, 10, e0116356. [Google Scholar] [CrossRef] [PubMed]
- Dalgleish, H.J.; Nelson, C.D.; Scrivani, J.A.; Jacobs, D.F. Consequences of shifts in abundance and distribution of American chestnut for restoration of a foundation forest tree. Forests 2016, 7, 4. [Google Scholar] [CrossRef]
- Paillet, F.L. Chestnut: History and ecology of a transformed species. J. Biogeogr. 2002, 29, 1517–1530. [Google Scholar] [CrossRef]
- Wang, G.G.; Hu, H. The replacements of American chestnut: A range-wide assessment based on data from forest inventory and published studies. In Proceedings of the 17th Biennial Southern Silvicultural Research Conference; Gordon, H.A., Conner, K.F., Haywood, J.D., Eds.; General Technical Report SRS-203; U.S. Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2015; pp. 513–515. [Google Scholar]
- De Bruijn, A.; Gustafson, E.J.; Kashian, D.M.; Dalgleish, H.J.; Sturtevant, B.R.; Jacobs, D.F. Decomposition rates of American chestnut (Castanea dentata) wood and implications for coarse woody debris pools. Can. J. For. Res. 2014, 44, 1575–1585. [Google Scholar] [CrossRef]
- Faison, E.K.; Foster, D.R. Did American chestnut really dominate the eastern forest? Arnoldia 2014, 72, 18–32. [Google Scholar]
- Seiler, J.; Jensen, E.; Niemiera, A.; Peterson, J. American chestnut Fagaceae Castanea dentata (Marsh.) Borkh., Ag Handbook 654, Virginia Tech Department of Forest Resources and Environmental Conservation. 2018. Available online: http://dendro.cnre.vt.edu/dendrology/syllabus/factsheet.cfm?ID=21 (accessed on 23 February 2018).
- Dalgleish, H.J.; Swihart, R.K. American chestnut past and future: Implications of restoration for resource pulses and consumer populations of eastern U.S. forests. Restor. Ecol. 2012, 20, 490–497. [Google Scholar] [CrossRef]
- Davis, D.E. Historical significance of American chestnut to Appalachian culture and ecology. In Restoration of American Chestnut to Forest Lands-Proceedings of a Conference and Workshop; Steiner, K.C., Carlson, J.E., Eds.; Natural Resources Report NPS/NCR/CUE/NRR—2006/001; National Park Service: Washington, DC, USA, 2006; pp. 53–60. [Google Scholar]
- Gravatt, F. Chestnut blight in Asia and North America. Unasylva 1949, 3, 2–7. [Google Scholar]
- O’Shea, T.J.; Ellison, L.E.; Stanley, T.R. Survival estimation in bats: Historical overview, critical appraisal, and suggestions for new approaches. In Sampling Rare or Elusive Species: Concepts, Designs, and Techniques foe Estimating Population Parameters; Thompson, W.L., Ed.; Island Press: Washington, DC, USA, 2004; pp. 297–336. [Google Scholar]
- O’Donnell, C.F.J. Population dynamics and survivorship in bats. In Ecological and Behavioral Methods for the Study of Bats, 2nd ed.; Kunz, T.H., Parsons, S., Eds.; Johns Hopkins University Press: Baltimore, MD, USA, 2009; pp. 158–176. [Google Scholar]
- Foster, G.; Humphrey, S.R.; Humphrey, P.P. Survival rate of young southeastern brown bats, Myotis austroriparius, in Florida. J. Mammal. 1978, 59, 299–304. [Google Scholar] [CrossRef]
- Stevenson, D.E.; Tuttle, M.D. Survivorship in the endangered gray bat (Myotis grisescens). J. Mammal. 1981, 62, 244–257. [Google Scholar] [CrossRef]
- Sedgeley, J.A.; O’Donnell, C.F.J. Roost use by long-tailed bats in South Canterbury: Examining predictions of roost-site selection in a highly fragmented landscape. N. Z. J. Ecol. 2004, 28, 1–18. [Google Scholar]
- O’Donnell, C.F.J.; Sedgeley, J.A. Causes and consequences of tree-cavity roosting in a temperate bat, Chalinolobus tuberculatus, from New Zealand. In Functional and Evolutionary Ecology of Bats; Zubaid, A., McCracken, G.F., Kunz, T.H., Eds.; Oxford University Press, Inc.: New York, NY, USA, 2006; pp. 308–328. [Google Scholar]
- O’Donnell, C.F.J. Distribution, Status and Conservation of Long-Tailed Bat (Chalinolobus tuberculatus) Communities in Canterbury, New Zealand; Environment Canterbury Report U00/38; Environment Canterbury: Christchurch, New Zealand, 2000. [Google Scholar]
- Brigham, R.M. Bats in forests: What we know and what we need to learn. In Bats in Forests: Conservation and Management; Lacki, M.J., Hayes, J.P., Kurta, A., Eds.; Johns Hopkins University Press: Baltimore, MD, USA, 2007; pp. 1–15. [Google Scholar]
- Jachowski, D.S.; Rota, C.T.; Dobony, C.A.; Ford, W.M.; Edwards, J.W. Seeing the forest through the trees: Considering roost-site selection at multiple spatial scales. PLoS ONE 2015, 11, e0150011. [Google Scholar]
- Mori, A.S.; Kitagawa, R. retention forestry as a major paradigm for safeguarding forest biodiversity in productive landscapes: A global meta-analysis. Biol. Conserv. 2014, 175, 65–73. [Google Scholar] [CrossRef]
- Cowden, M.M.; Hart, J.L.; Schweitzer, C.J.; Dey, D.C. Effects of intermediate-scale wind disturbance on composition, structure, and succession in Quercus stands: Implications for natural disturbance-based silviculture. For. Ecol. Manag. 2014, 330, 240–251. [Google Scholar] [CrossRef]
- Puettmann, K. Silvicultural challenges and options in the context of global change: ‘simple’ fixes and opportunities for new management approaches. J. For. 2011, 109, 321–331. [Google Scholar]
- Kern, C.C.; Burton, J.I.; Raymond, P.; D’Amato, A.W.; Keeton, W.S.; Royo, A.A.; Walters, M.B.; Webster, C.R.; Willis, J.L. Challenges facing gap-based silviculture and possible solutions for mesic northern forests in North America. Forestry 2017, 90, 4–17. [Google Scholar] [CrossRef]
- Lacki, M.J.; Johnson, J.S.; Baker, M.D. Temperatures beneath bark of dead trees used as roosts by Myotis volans in forests of the Pacific Northwest, USA. Acta Chiropter. 2013, 15, 143–151. [Google Scholar] [CrossRef]
- Willis, C.K.R.; Brigham, R.M. Social thermoregulation exerts more influence than microclimate on forest roost preferences by a cavity-dwelling bat. Behav. Ecol. Sociobiol. 2007, 62, 97–108. [Google Scholar] [CrossRef]
- Jacobs, D.F.; Dalgleish, H.J.; Nelson, C.D. A conceptual framework for restoration of threatened plants: The effective model of American chestnut (Castanea dentata) restoration. New Phytol. 2013, 197, 378–393. [Google Scholar] [CrossRef] [PubMed]
- Palmer, J.M.; Drees, K.P.; Foster, J.T.; Lindner, D.L. Extreme sensitivity to ultraviolet light in the fungal pathogen causing white-nose syndrome of bats. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.L.; Schlarbaum, S.E.; Pinchot, C.C.; Anagnostakis, S.L.; Saunders, M.R.; Thomas-Van Gundy, M.; Schaberg, P.; McKenna, J.; Bard, J.F.; Berrang, P.C.; et al. Reintroduction of American chestnut in the National Forest system. J. For. 2014, 112, 502–512. [Google Scholar] [CrossRef]
- Hartley, M.J. Rationale and methods for conserving biodiversity in plantation forests. For. Ecol. Manag. 2002, 155, 81–95. [Google Scholar] [CrossRef]
- Mori, A.S.; Tatsumi, S.; Gustafsson, L. Landscape properties affect biodiversity response to retention approaches in forestry. J. Appl. Ecol. 2017, 54, 1627–1637. [Google Scholar] [CrossRef]
- Manning, A.D.; Fischer, J.; Lindenmayer, D.B. Scattered trees are keystone structures—Implications for conservation. Biol. Conserv. 2006, 132, 311–321. [Google Scholar] [CrossRef]
- Hallinger, M.; Johansson, V.; Schmalholz, M.; Sjöberg, S.; Ranius, T. Factors driving tree mortality in retained forest fragments. For. Ecol. Manag. 2016, 368, 163–172. [Google Scholar] [CrossRef]
- Périé, C.; de Blois, S. Dominant forest tree species are potentially vulnerable to climate change over large portions of their range even at high latitudes. PeerJ 2016, 4, e2218. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| % Difference | % Difference | |||
|---|---|---|---|---|
| Bat Species | na | in Diameter | n | in Height |
| M. austroriparius | 4 | 0.9 (0.25) | 2 | 0.85 (0.6) |
| M. auriculus | 1 | 1.06 (–) | 1 | 1.3 (–) |
| M. californicus | 2 | 1.77 (0.23) | 2 | 1.58 (0.005) |
| M. evotis | 1 | 0.83 (–) | 1 | 0.75 (–) |
| M. keenii | 2 | 1.7 (0.4) | ||
| M. lucifugus | 4 | 1.12 (0.06) | 3 | 1.1 (0.06) |
| M. septentrionalis | 14 | 1.33 (0.12) | 11 | 2.02 (0.84) |
| M. sodalis | 8 | 1.72 (0.23) | 3 | 1.73 (0.38) |
| M. thysanodes | 3 | 1.41 (0.08) | 3 | 1.62 (0.51) |
| M. volans | 4 | 1.29 (0.09) | 4 | 1.69 (0.26) |
| M. yumanensis | 1 | 3.74 (–) | 1 | 2.02 (–) |
| Adj-R2 | Parameter | β | SE | F | p | |
|---|---|---|---|---|---|---|
| Maximum | 0.0162 | Roost diameter | −1.347 | 1.23 | 1.2 | 0.297 |
| 0.0771 | Roost height | 8.748 | 7.35 | 1.42 | 0.299 | |
| 0.1221 | Roost diameter | −1.825 | 1.66 | 1.1 | 0.352 | |
| Roost height | 12.061 | 7.78 | 1.55 | 0.218 | ||
| Mean | −0.0021 | Mean diameter | 0.223 | 0.23 | 0.94 | 0.341 |
| −0.0155 | Mean height | 0.451 | 0.55 | 0.66 | 0.424 | |
| −0.0662 | Mean diameter | 0.016 | 0.38 | 0.04 | 0.967 | |
| Mean height | 0.421 | 0.92 | 0.46 | 0.653 |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lacki, M.J. Restoration of Legacy Trees as Roosting Habitat for Myotis Bats in Eastern North American Forests. Diversity 2018, 10, 29. https://doi.org/10.3390/d10020029
Lacki MJ. Restoration of Legacy Trees as Roosting Habitat for Myotis Bats in Eastern North American Forests. Diversity. 2018; 10(2):29. https://doi.org/10.3390/d10020029
Chicago/Turabian StyleLacki, Michael J. 2018. "Restoration of Legacy Trees as Roosting Habitat for Myotis Bats in Eastern North American Forests" Diversity 10, no. 2: 29. https://doi.org/10.3390/d10020029
APA StyleLacki, M. J. (2018). Restoration of Legacy Trees as Roosting Habitat for Myotis Bats in Eastern North American Forests. Diversity, 10(2), 29. https://doi.org/10.3390/d10020029