On the Phylogenetic Position of the Weevil Tribe Acentrusini Alonso-Zarazaga, 2005 (Coleoptera: Curculionidae: Curculioninae)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Taxonomic and Morphological Methods
2.2. Characters Used for Phylogenetic and Multi-Dimensional Phenetic Analyses
2.2.1. Morphological Characters
2.2.2. Host Plant Characters
2.3. Phylogenetic and Multi-Dimensional Phenetic Methods
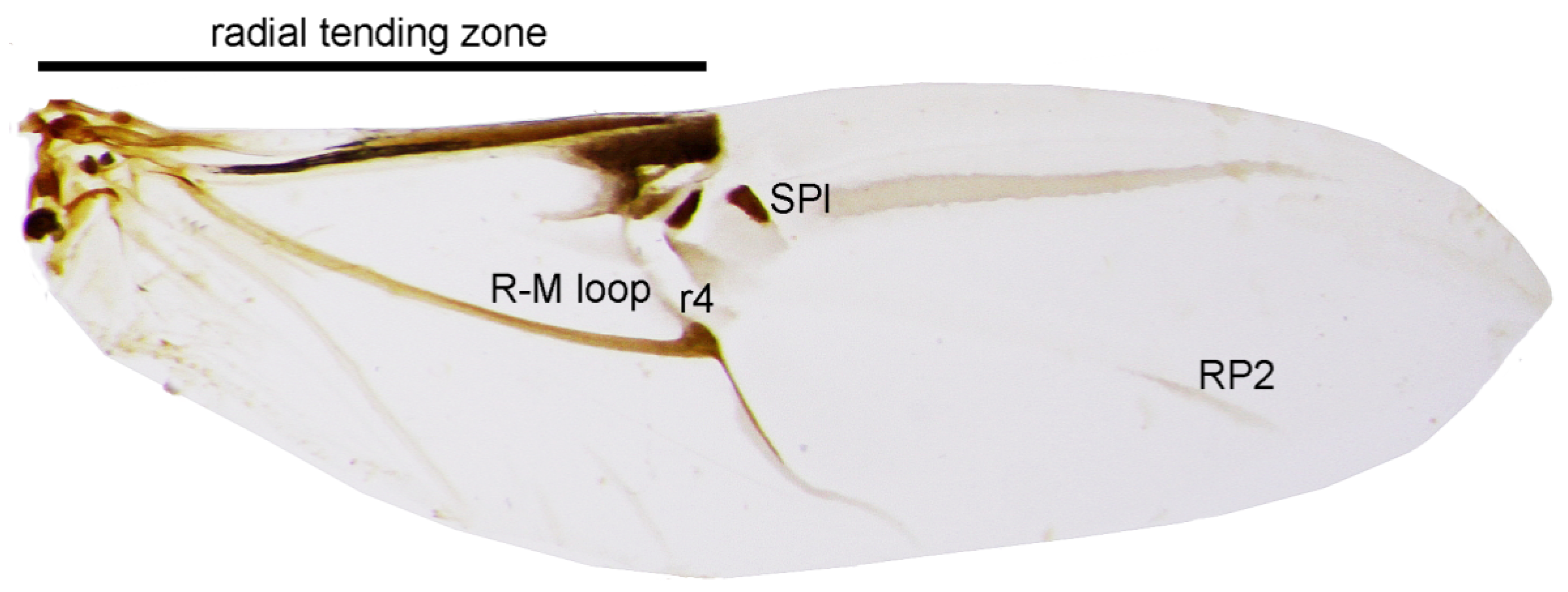
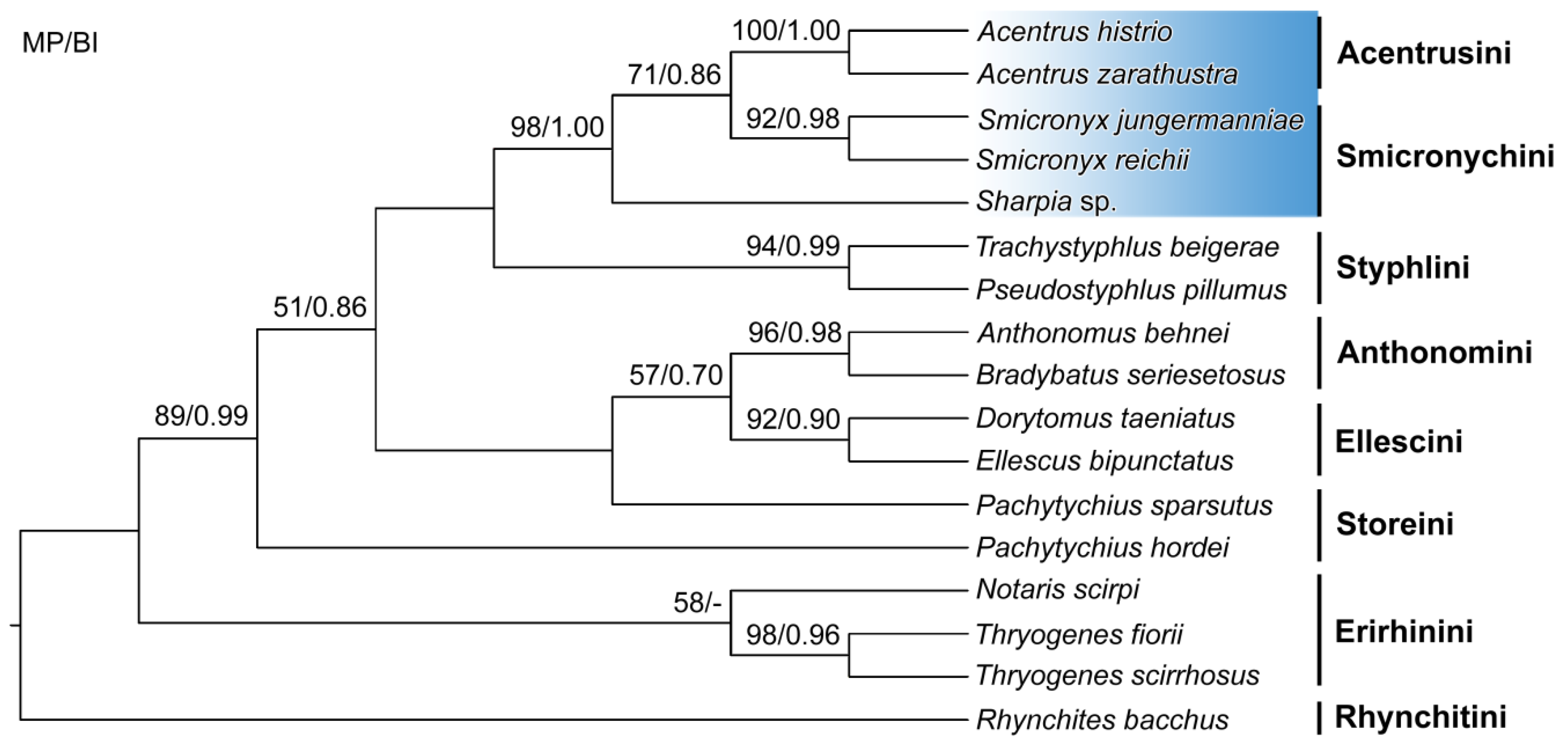
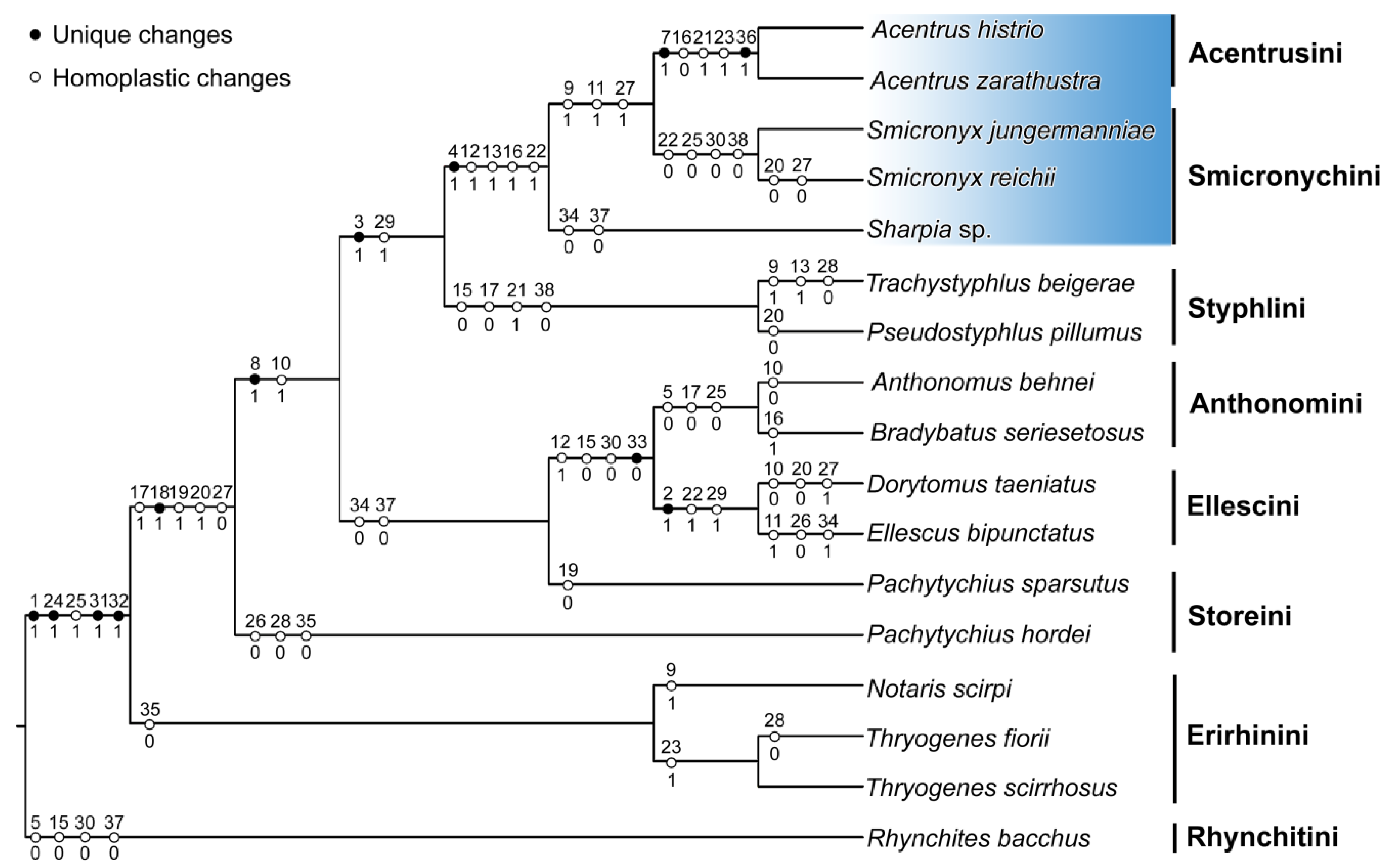
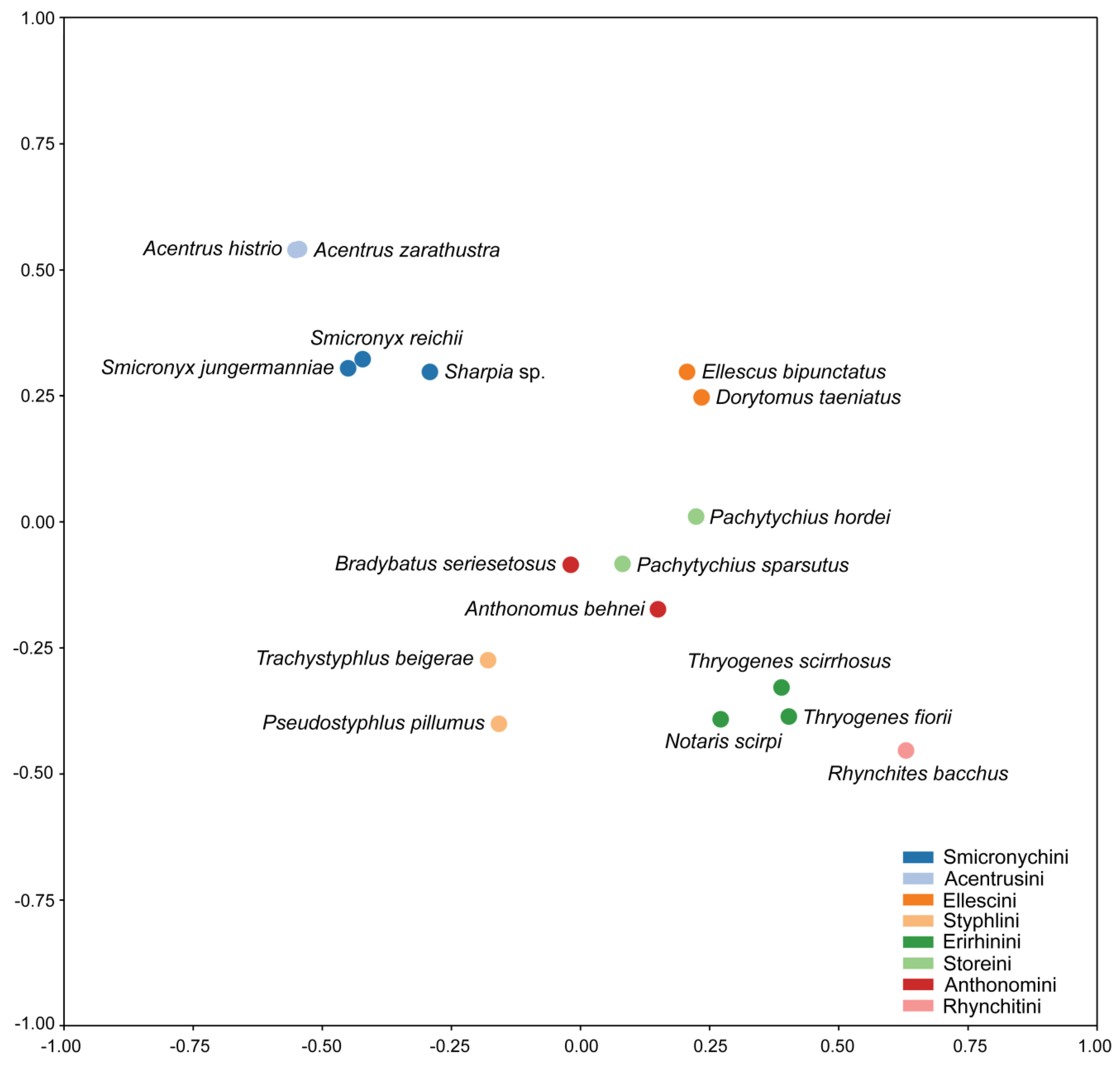
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alonso-Zarazaga, M.A. Diagnosis preliminares de nuevos táxones de Curculionidae (Coleoptera). Bol. SEA 2005, 37, 89–93. [Google Scholar]
- Košťál, M. Revision of the genus Acentrus Desmarest, 1839 (Coleoptera: Curculionidae). Koleopt. Rdsch. 2014, 84, 329–336. [Google Scholar]
- Caldara, R.; Franz, N.M.; Oberprieler, R.G. Curculioninae Latreille, 1802: Introduction, Phylogeny. In Handbook of Zoology Volume 3; Leschen, R.A.B., Beutel, R.G., Eds.; Walter de Gruyter GmbH: Berlin, Germany; Boston, MA, USA, 2014; Volume 3, pp. 589–628. [Google Scholar]
- Alonso–Zarazaga, M.A.; Barrios, H.; Borovec, R.; Bouchard, P.; Caldara, R.; Colonnelli, E.; Gültekin, L.; Hlaváč, P.; Korotyaev, B.; Lyal, C.H.C.; et al. Cooperative Catalogue of Palaearctic Coleoptera Curculionoidea; Monografias Electrónicas SEA 8; Sociedad Entomológica Aragonesa, S.E.A.: Zaragosa, Spain, 2017; p. 729. [Google Scholar]
- Oberprieler, R.G.; Anderson, R.S.; Marvaldi, A.E. Curculionoidea Latreille, 1802: Introduction, Phylogeny. In Handbook of Zoology Volume 3; Leschen, R.A.B., Beutel, R.G., Eds.; Walter de Gruyter GmbH: Berlin, Germany; Boston, MA, USA, 2014; Volume 3, pp. 285–300. [Google Scholar]
- Košťál, M.; Košťálová, Z. Description of the male of Acentrus boroveci Košťál, 2014 (Coleoptera: Curculionidae: Acentrusini). Zootaxa 2016, 4121, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Byng, J.W.; Chase, M.W.; Christenhusz, M.J.M.; Fay, M.F.; Judd, W.S.; Mabberley, D.J.; Sennikov, A.N.; Soltis, D.E.; Soltis, P.S.; Stevens, P.F.; et al. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar]
- Swofford, D.L. PAUP*. In Phylogenetic Analysis Using Parsimony (*and Other Methods), version 4; Sinauer Associates: Sunderland, MA, USA, 2002; p. 144. [Google Scholar]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Version 3.40. Available online: http://mesquiteproject.org (accessed on 30 April 2018).
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Subfamily | Tribe | Species | Character No. | |||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | |||||||||||||||||||||||||||||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 0 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 0 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 0 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | ||||
| Attalabidae | Rhynchitinae | Rhynchitini | Rhynchites bacchus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 1 |
| Curculionidae | Brachycerinae | Erirhinini | Notaris scirpi | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 |
| Curculionidae | Brachycerinae | Erirhinini | Thryogenes fiorii | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 0 | n | n | n | n | n | n | 0 | 0 | 1 | 1 |
| Curculionidae | Brachycerinae | Erirhinini | Thryogenes scirrhosus | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 |
| Curculionidae | Curculioninae | Acentrusini | Acentrus histrio | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Curculionidae | Curculioninae | Acentrusini | Acentrus zarathustra | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Curculionidae | Curculioninae | Anthonomini | Anthonomus behnei | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 1 |
| Curculionidae | Curculioninae | Anthonomini | Bradybatus seriesetosus | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 1 |
| Curculionidae | Curculioninae | Ellescini | Dorytomus taeniatus | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 1 |
| Curculionidae | Curculioninae | Ellescini | Ellescus bipunctatus | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 0 | 1 |
| Curculionidae | Curculioninae | Smicronychini | Sharpia sp. | 1 | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 1 |
| Curculionidae | Curculioninae | Smicronychini | Smicronyx jungermanniae | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 0 |
| Curculionidae | Curculioninae | Smicronychini | Smicronyx reichii | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 0 |
| Curculionidae | Curculioninae | Storeini | Pachytychius sparsutus | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 1 |
| Curculionidae | Curculioninae | Storeini | Pachytychius hordei | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | n | n | n | n | n | n | 0 | 0 | 1 | 1 |
| Curculionidae | Curculioninae | Styphlini | Trachystyphlus beigerae | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | n | n | n | n | n | n | 1 | 0 | 1 | 0 |
| Curculionidae | Curculioninae | Styphlini | Pseudostyphlus pillumus | 1 | 0 | 1 | 0 | 1 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Košťál, M.; Vďačný, P. On the Phylogenetic Position of the Weevil Tribe Acentrusini Alonso-Zarazaga, 2005 (Coleoptera: Curculionidae: Curculioninae). Diversity 2018, 10, 34. https://doi.org/10.3390/d10020034
Košťál M, Vďačný P. On the Phylogenetic Position of the Weevil Tribe Acentrusini Alonso-Zarazaga, 2005 (Coleoptera: Curculionidae: Curculioninae). Diversity. 2018; 10(2):34. https://doi.org/10.3390/d10020034
Chicago/Turabian StyleKošťál, Michael, and Peter Vďačný. 2018. "On the Phylogenetic Position of the Weevil Tribe Acentrusini Alonso-Zarazaga, 2005 (Coleoptera: Curculionidae: Curculioninae)" Diversity 10, no. 2: 34. https://doi.org/10.3390/d10020034
APA StyleKošťál, M., & Vďačný, P. (2018). On the Phylogenetic Position of the Weevil Tribe Acentrusini Alonso-Zarazaga, 2005 (Coleoptera: Curculionidae: Curculioninae). Diversity, 10(2), 34. https://doi.org/10.3390/d10020034