Sonar Surveys for Bat Species Richness and Activity in the Southern Kalahari Desert, Kgalagadi Transfrontier Park, South Africa

Abstract
:1. Introduction
2. Materials and Methods
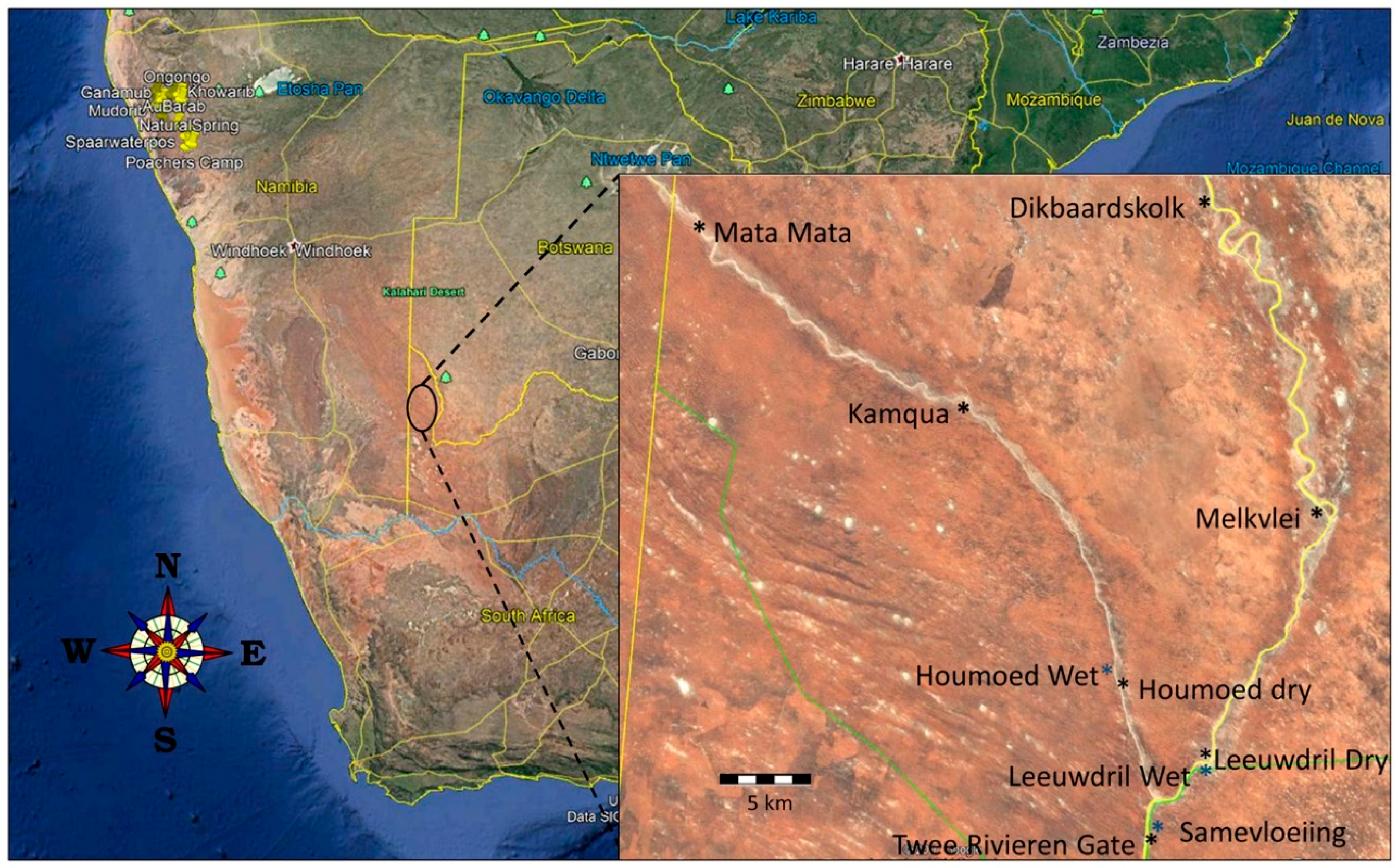
2.1. Study Area
2.2. Data Collection
2.3. Data Analysis
3. Results
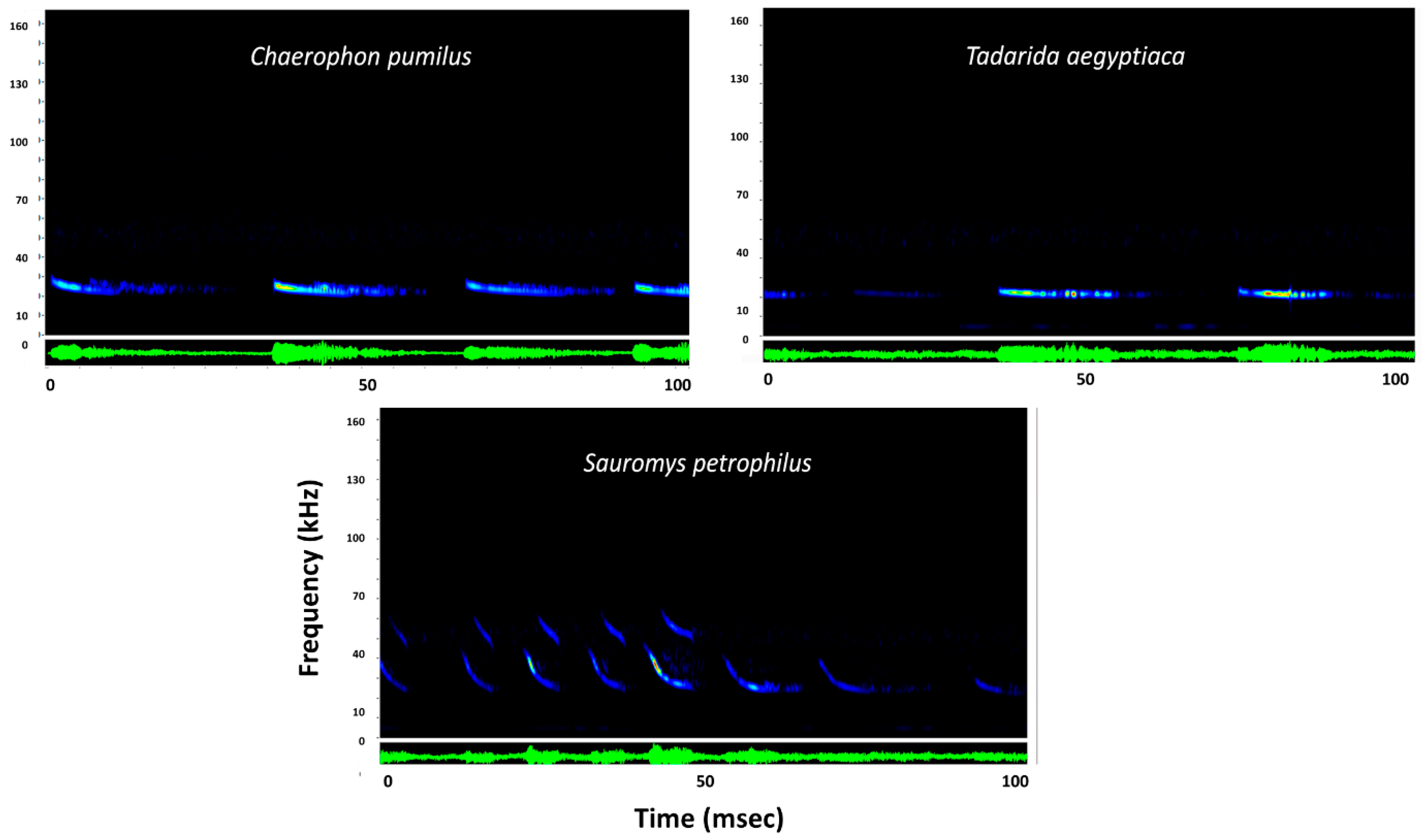
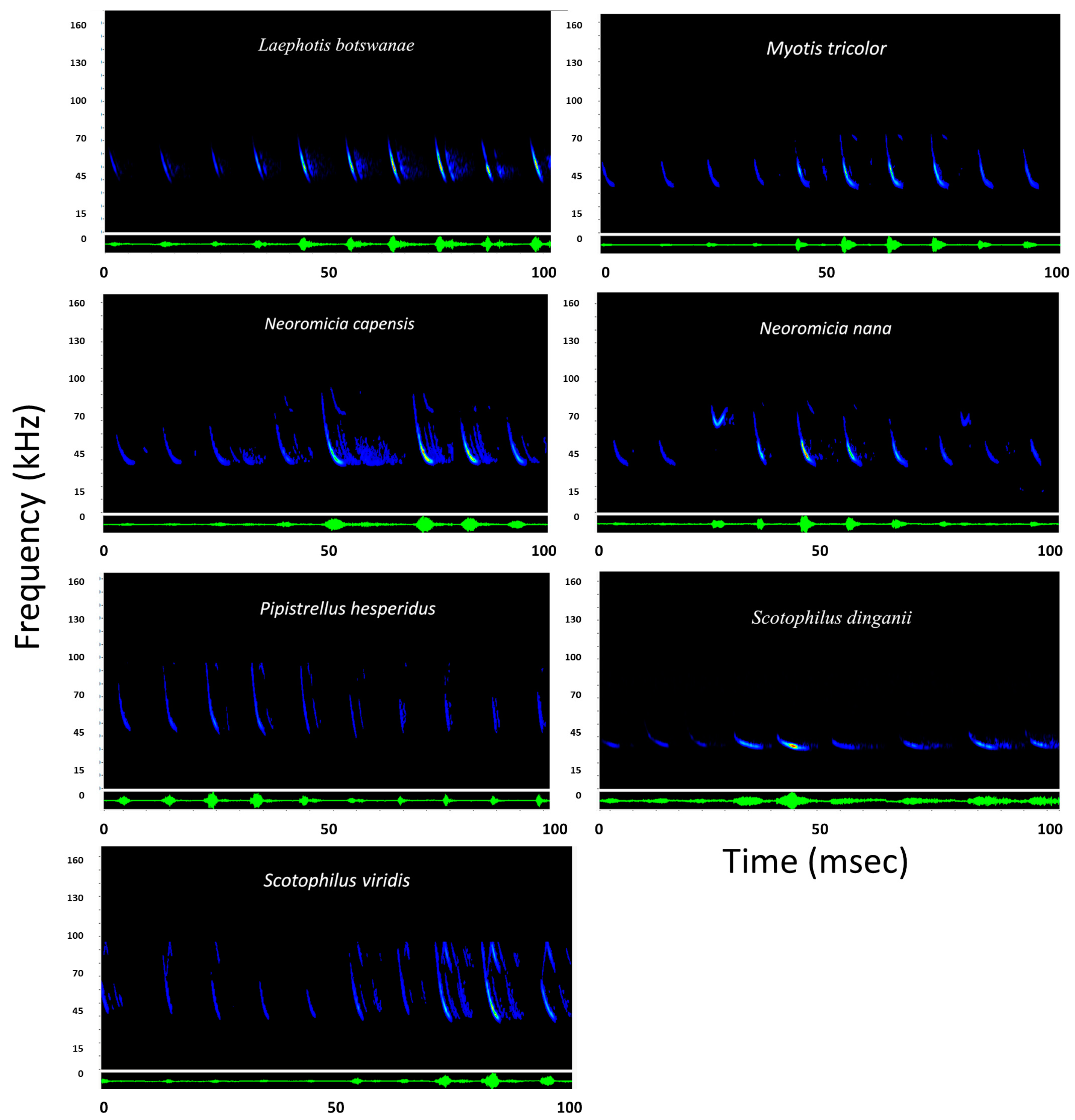
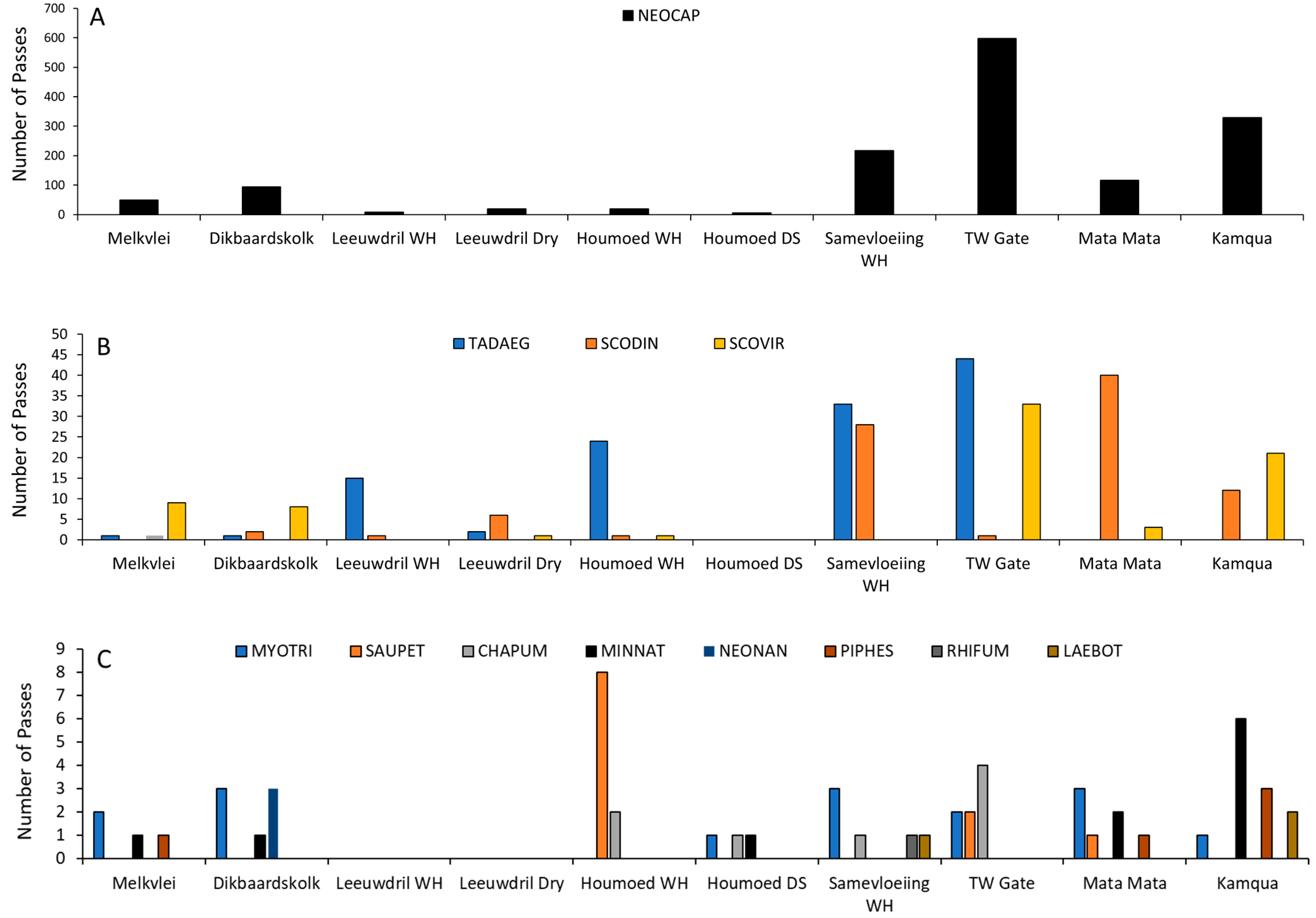
3.1. Sonar Capture Results
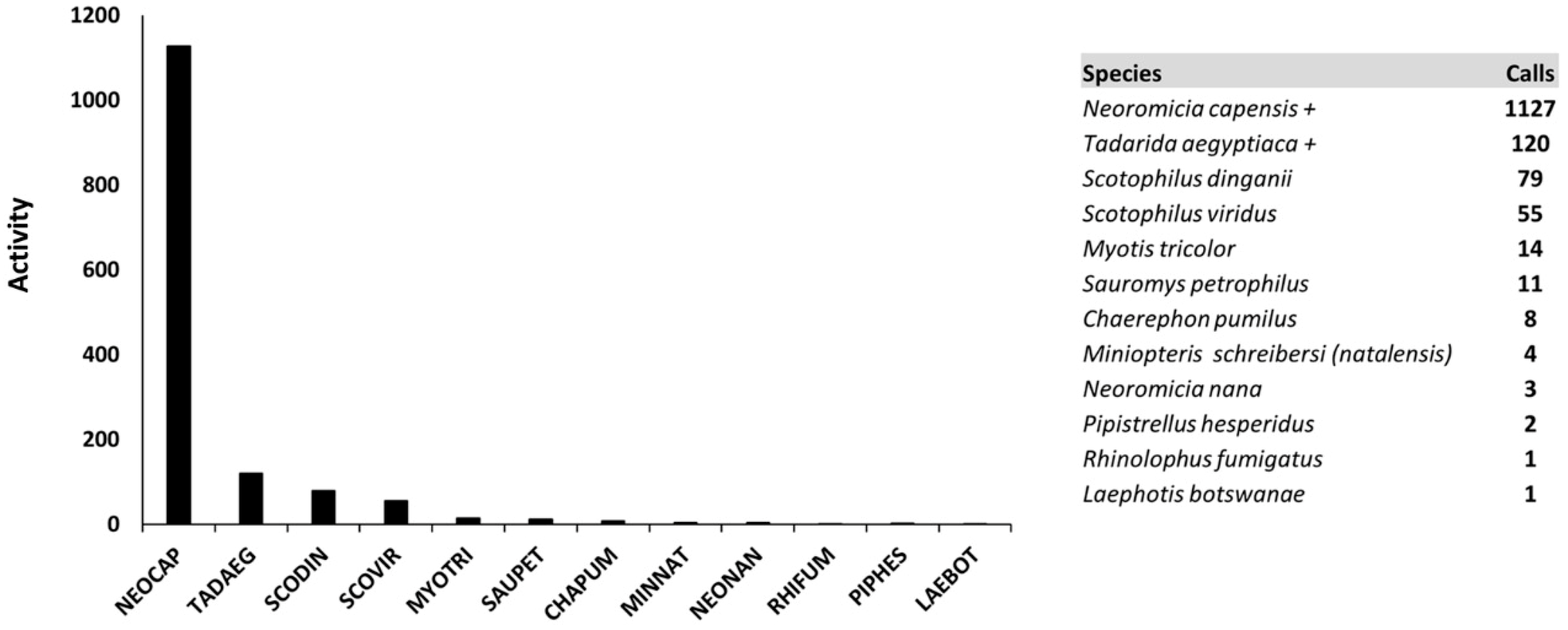
3.2. Species Richness and Activity
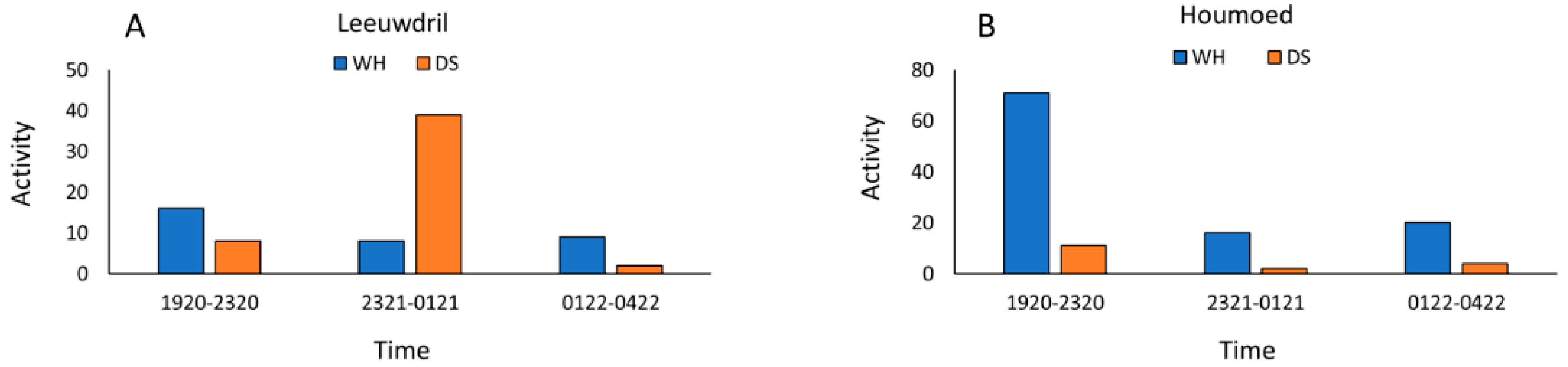
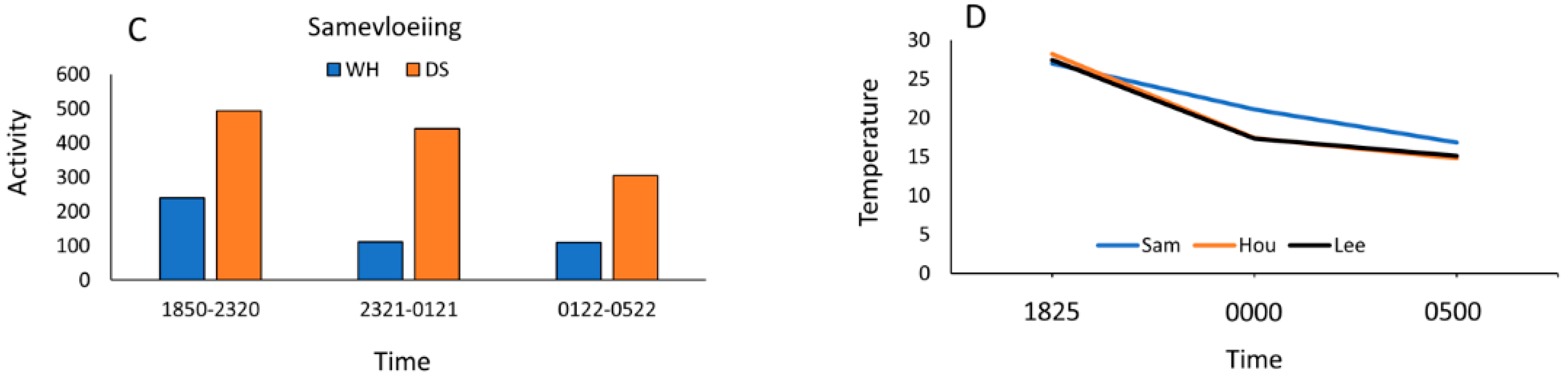
3.3. Wet versus Dry Site Comparisons
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kiker, G.A. South African study on climate change: Vulnerability and adaptation assessment section. In A National Climate Change Response Strategy for South Africa; Department of Environmental Affairs and Tourism: Pretoria, South Africa, 2004. [Google Scholar]
- Cowlings, R.M.; Rundel, P.W.; Desmet, P.G.; Esler, K.J. Extraordinary high regional-scale plant diversity in southern African arid lands: Subcontinental and global comparison. Divers. Distrib. 1998, 4, 27–36. [Google Scholar]
- Hanks, J. Transfrontier conservation areas (TFCAs) in South Africa. J. Sustain. For. 2003, 17, 127–148. [Google Scholar] [CrossRef]
- Bergström, R.; Skarpe, C. The abundance of large wild herbivores in a semi-arid savanna in relation to seasons, pans and livestock. Afr. J. Ecol. 1999, 37, 12–26. [Google Scholar] [CrossRef]
- Blaum, N.; Rossmanith, E.; Jeltsch, F. Land use affects rodent communities in Kalahari savannah rangelands. Afr. J. Ecol. 2007, 45, 189–195. [Google Scholar] [CrossRef]
- Monadjem, A.; Taylor, P.J.; Cotterill, F.P.D.; Schoeman, M.C. Bats of Southern and Central Africa; Wits University Press: Johannesburg, South Africa, 2001. [Google Scholar]
- Jones, G.; Jacobs, D.S.; Kunz, T.H.; Willig, M.R.; Racey, P.A. Carpe noctem: The importance of bats as bioindicators. Endanger. Spec. Res. 2009, 8, 93–115. [Google Scholar] [CrossRef]
- Adams, R.A. Bat reproduction declines when conditions mimic climate change projections for western North America. Ecology 2010, 91, 2437–2445. [Google Scholar] [CrossRef] [PubMed]
- Tews, J.; Blaum, N.; Jetsch, F. Structural and animal species diversity in arid and semi-arid savannas of the southern Kalahari. Ann. Arid Zone 2006, 43, 413–425. [Google Scholar]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.; Underwood, E.C.; D’amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial ecoregions of the world: A new map of life on Earth. Bioscience 2001, 15, 933–938. [Google Scholar] [CrossRef]
- White, F. The Vegetation of Africa; UNESCO Press: Paris, France, 1983. [Google Scholar]
- Taylor, P.J.; Monadjem, A.; Steyn, J.N. Seasonal patterns in habit use by insectivorous bats in a subtropical African agro-ecosystem dominated by macadamia orchards. Zoo. J. Linnaean Soc. 2013, 167, 191–207. [Google Scholar]
- Monadjem, A.; Shapiro, J.T.; Mtsetfwa, F.; Reside, A.E.; McCleery, R.A. Acoustic call library and detection distances for bats of Swaziland. Acta Chiropterol. 2017, 19, 175–187. [Google Scholar] [CrossRef]
- Geluso, K.N.; Geluso, K. Effects of environmental factors on capture rates of insectivorous bats, 1971–2005. J. Mammal. 2012, 93, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Korine, C.; Adams, R.A.; Russo, D.; Jacobs, D. Bats and water: Anthropogenic alterations threaten global bat populations. In Bats in the Anthropocene: Conservation of Bats in a Changing World; Voigt, C., Kingston, T., Eds.; Springer Press: New York, NY, USA, 2016; pp. 188–215. [Google Scholar]
- Erasmus, B.F.N.; van Jaarveld, A.S.; Chown, S.L.; Kshatriya, M.; Wessels, K.J. Vulnerability of South African animal taxa to climate change. Glob. Chang. Biol. 2002, 8, 679–693. [Google Scholar] [CrossRef]
- Biggs, R.; Bohensky, E.; Desanker, P.V.; Fabricius, C.; Lynam, T.; Misselhorn, A.A.; Musvoto, C.; Mutale, M.; Reyers, B.; Scholes, R.J.; et al. The Southern Africa Millennium Ecosystem Assessment; Council for Scientific and Industrial Research: Pretoria, South Africa, 2004. [Google Scholar]
- Adams, R.A.; Hayes, M.A. Water availability and successful lactation by bats as related to climate change in western regions of North America. J. Anim. Ecol. 2008, 77, 1115–1121. [Google Scholar] [CrossRef] [PubMed]
- Hayes, M.A.; Adams, R.A. Simulated bat populations erode when exposed to climate change projections for western North America. PLoS ONE 2017, 12, e0180693. [Google Scholar] [CrossRef] [PubMed]
- Adams, R.A.; Hayes, M.A. Assemblage-level analysis of sex-ratios in Coloradan bats in relation to climate variables: A model for future expectations. Glob. Ecol. Conserv. 2018, 14, e00379. [Google Scholar] [CrossRef]
- Adams, R.A. Climate change associated with disruption in altitudinal migration and population declines in some Rocky Mountain bat species. J. Zool. Lond. 2017, in press. [Google Scholar]
- Kunz, T.H.; Braun de Torrez, E.; Bauer, D.; Lobova, T.; Fleming, T.H. Ecosystem services provided by bats. Ann. N. Y. Acad. Sci. 2011, 1223, 1–38. [Google Scholar] [CrossRef] [PubMed]
- Maine, J.J.; Boyles, J.G. Bats initiate vital agroecological interactions in corn. Proc. Natl. Acad. Sci. USA 2015, 112, 12438–12443. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Survey Date | Latitude | Longitude | Elevation |
|---|---|---|---|---|
| Kamqua | 10 April 2016 | S 26°01′20.7″ | E 20°24′17.3″ | 904 m |
| Mata Camp | 10 April 2016 | S 25°49′26.98″ | E 20°00′25.13″ | 952 m |
| Samevloeiing Water Hole | 11 April 2016 | S 26°28′10.3″ | E 20°36′55.9″ | 869 m |
| Twee Rivieren Gate | 11 April 2016 | S 26°28′22.3″ | E 20°36′48.0″ | 870 m |
| Houmoed Water Hole | 12 April 2016 | S 26°20′22.5″ | E 20°35′27.6″ | 878 m |
| Houmoed Dry Site | 12 April 2016 | S 26°20′53.6″ | E 20°35′40.4″ | 873 m |
| Leeuwdril Water Hole | 13 April 2016 | S 26°23′07.5″ | E 20°41′51.6″ | 875 m |
| Leeuwdril Dry Site | 13 April 2016 | S 26°22′35.7″ | E 20°41′50.6″ | 878 m |
| Melkvlei | 14 April 2016 | S 26°08′19.4″ | E 20°51′25.8″ | 892 m |
| Dikbaardskolk | 14 April 2016 | S 26°46′13.7″ | E 20°43′46.9″ | 926 m |
| Rhinolophidae | Molossidae | Miniopteridae | Vespertilionidae |
|---|---|---|---|
| Rhinolophus fumigatus | Chaerephon pumilus Sauromys petrophilus Tadarida aegyptiaca | Miniopteris schreibersi (natalensis) | Laephotis botswanae Myotis tricolor Neoromicia capensis Neoromicia nana Pipistrellus hesperidus Scotophilus dinganii Scotophilus viridus |
| Species | Family | Fc | Dur | Fmax | Fmin | N | MR |
|---|---|---|---|---|---|---|---|
| Rhinolophus fumigatus | Rhinolophidae | 53.9 | 40.1 | 54.4 | 53.6 | 1 | 0.9 |
| Chaerophon pumilus | Molossidae | 22.9 ± 0.3 | 5.6 ± 1.5 | 25 ± 0.1 | 22.4 ± 0.4 | 2 | 0.9 |
| Sauromys petrophilus | Molossidae | 24.0 ± 0.2 | 4.3 ± 0.7 | 30.3 ± 3.2 | 23.7 ± 0.1 | 2 | 1 |
| Tadarida aegyptiacus | Molossidae | 21.6 ± 1.2 | 4.9 ± 0.6 | 25.2 ± 1.1 | 21.2 ± 1.1 | 5 | 1 |
| Miniopteris schreibersi | Miniopteridae | 47.6 ± 6.5 | 2.6 ± 0.4 | 76.7 ± 7.7 | 53.2 ± 4.9 | 3 | 1 |
| Laephotis botswanae | Vespertilionidae | 38.8 ± 0.7 | 2.2 ± 0.3 | 68.5 ± 0.5 | 37.9 ± 0.9 | 3 | 1 |
| Neoromicia capensis | Vespertilionidae | 36.9 ± 0.6 | 3.5 ± 0.5 | 39.6 ± 4.3 | 36.7 ± 0.6 | 10 | 1 |
| Neoromicia nana | Vespertilionidae | 61.5 ± 8.9 | 3.7 ± 0.3 | 74.3 ± 6.2 | 61.1 ± 3.5 | 2 | 0.8 |
| Scotophilus dinganii | Vespertilionidae | 33.9 ± 2.4 | 3.3 ± 0.9 | 40.9 ± 2.1 | 33.4 ± 2.0 | 4 | 1 |
| Scotophilis viridus | Vespertilionidae | 37.5 ± 1.6 | 4.3 ± 0.3 | 70.6 ± 6.6 | 36.7 ± 1.8 | 3 | 0.9 |
| Pipistrellus hesperidus | Vespertilionidae | 43.3 ± 3.1 | 2.3 ± 0.03 | 53.6 ± 1.6 | 42.9 ± 3.4 | 2 | 0.85 |
| Myotis tricolor | Vespertilionidae | 41.8 ± 4.3 | 2.9 ± 0.9 | 81.5 ± 6.2 | 38.8 ± 2.2 | 6 | 1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adams, R.A.; Kwiecinski, G. Sonar Surveys for Bat Species Richness and Activity in the Southern Kalahari Desert, Kgalagadi Transfrontier Park, South Africa. Diversity 2018, 10, 103. https://doi.org/10.3390/d10030103
Adams RA, Kwiecinski G. Sonar Surveys for Bat Species Richness and Activity in the Southern Kalahari Desert, Kgalagadi Transfrontier Park, South Africa. Diversity. 2018; 10(3):103. https://doi.org/10.3390/d10030103
Chicago/Turabian StyleAdams, Rick A., and Gary Kwiecinski. 2018. "Sonar Surveys for Bat Species Richness and Activity in the Southern Kalahari Desert, Kgalagadi Transfrontier Park, South Africa" Diversity 10, no. 3: 103. https://doi.org/10.3390/d10030103
APA StyleAdams, R. A., & Kwiecinski, G. (2018). Sonar Surveys for Bat Species Richness and Activity in the Southern Kalahari Desert, Kgalagadi Transfrontier Park, South Africa. Diversity, 10(3), 103. https://doi.org/10.3390/d10030103