A Combined Molecular and Morphological Approach to Explore the Higher Phylogeny of Entimine Weevils (Coleoptera: Curculionidae), with Special Reference to South American Taxa

Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimens and Taxon Sampling
2.2. Molecular Data
2.2.1. DNA Isolation, PCR Amplification and Sequencing
2.2.2. Alignment of Sequences
2.3. Morphological Characters
2.4. Phylogenetic Analyses
3. Results
3.1. Annotated Structural Alignment
3.2. Phylogenetic Trees
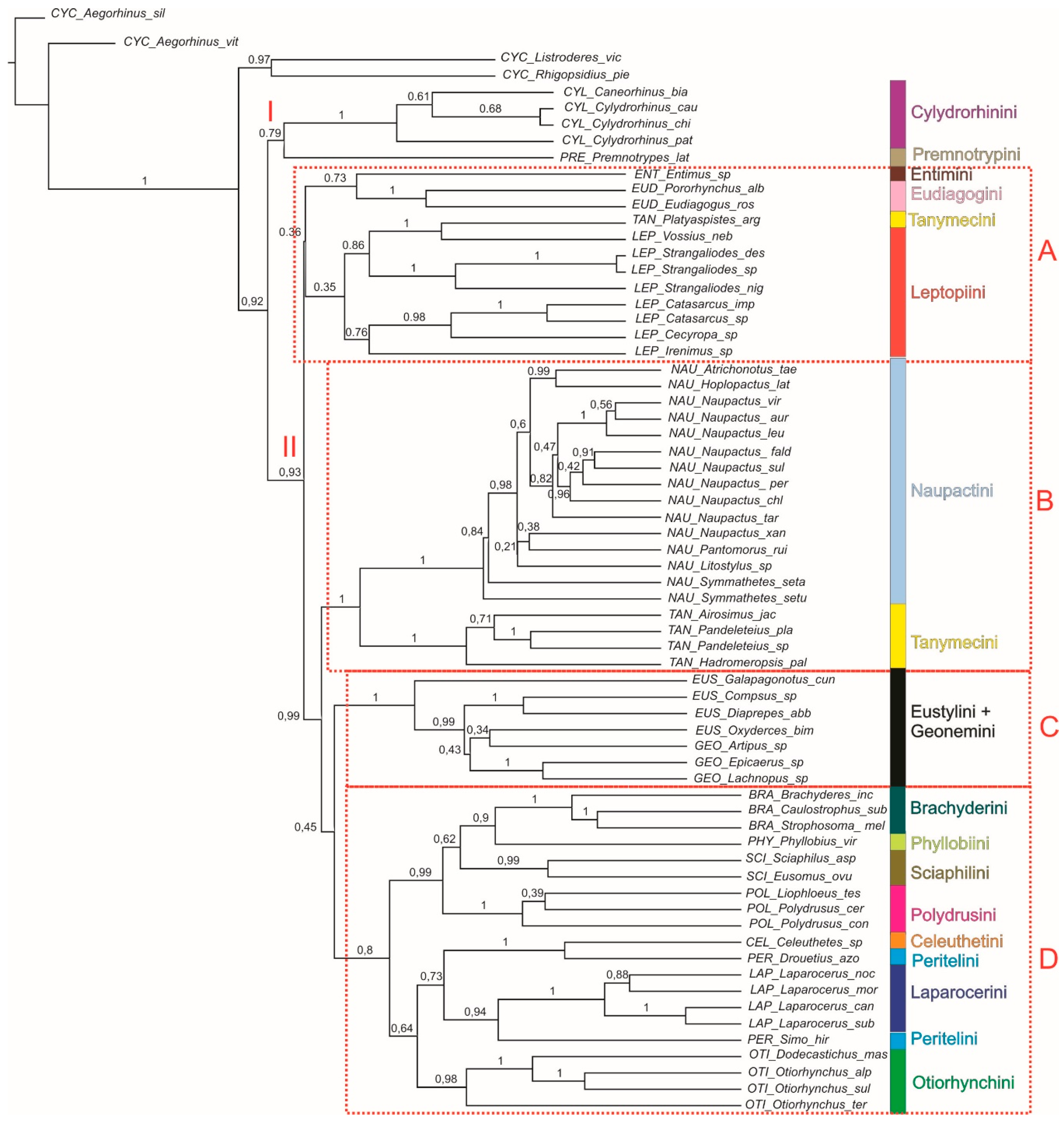
3.2.1. Bayesian Analysis
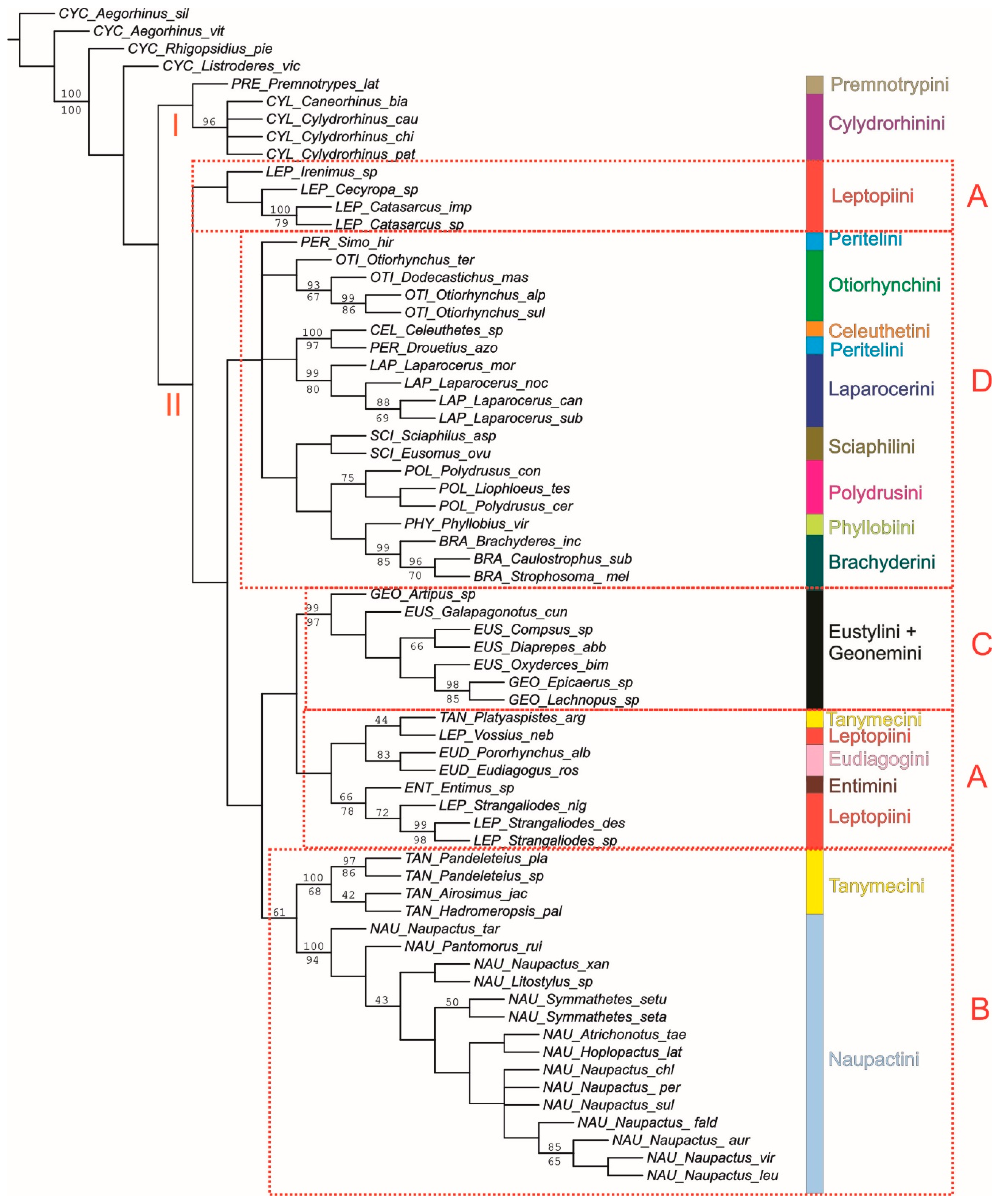
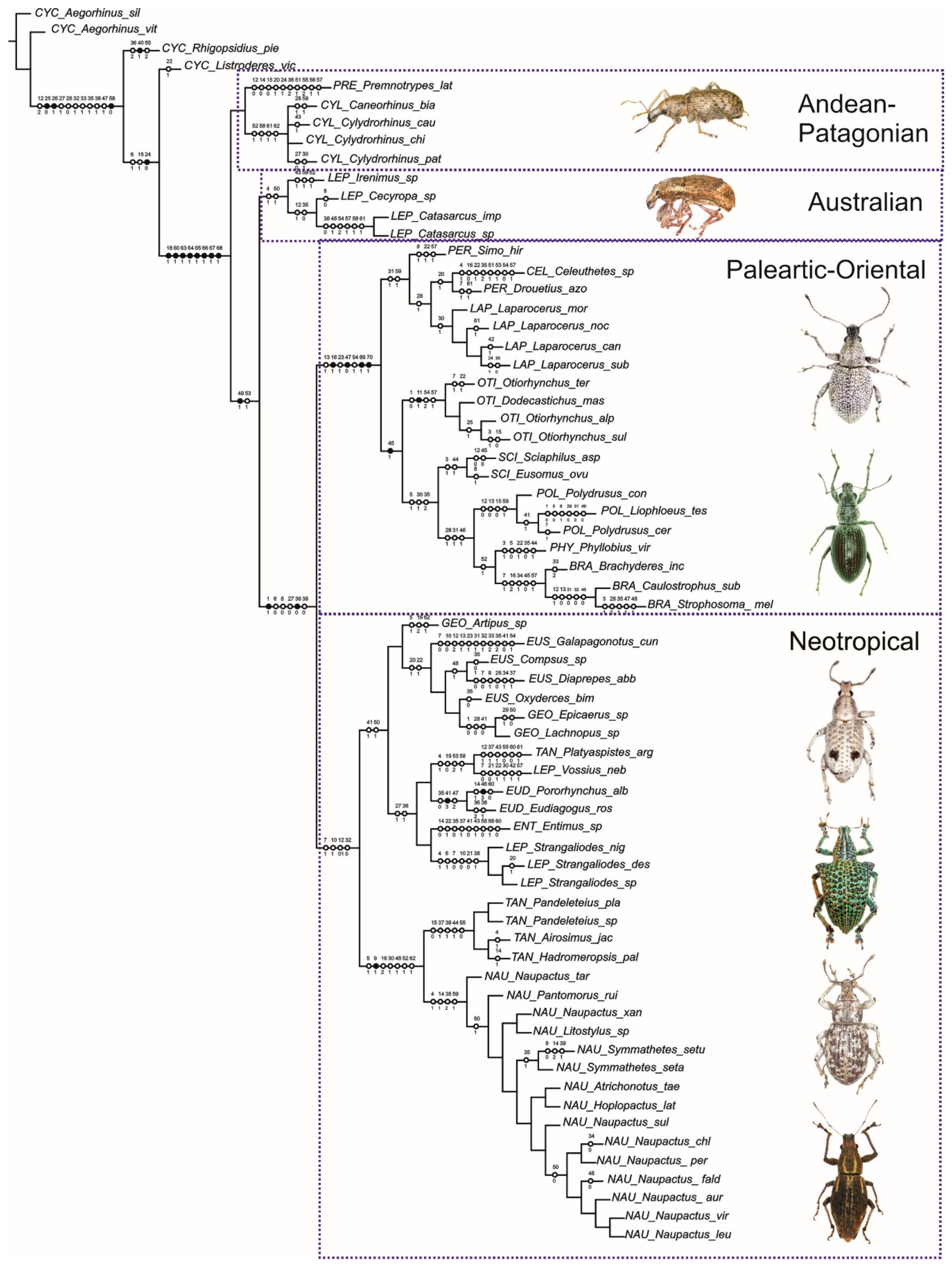
3.2.2. Combined Maximum-Parsimony Analysis and Synapomorphies
3.2.3. Separate Molecular Analyses (Bayesian and Parsimony)
4. Discussion
4.1. Southern South American Entiminae and Cyclominae
4.2. Neotropical Entiminae
4.3. Palaearctic, and Australian and Oriental Entiminae
4.4. General Remarks
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Thompson, R.T. Observations on the morphology and classification of weevils (Coleoptera, Curculionoidea) with a key to major groups. J. Nat. Hist. 1992, 26, 835–891. [Google Scholar] [CrossRef]
- Alonso-Zarazaga, M.A.; Lyal, C.H.C. A World Catalogue of Families and Genera of Curculionoidea: (Insecta: Coleoptera) Excepting Scolytidae and Platypodidae; Entomopraxis. D.C.P.: Barcelona, Spain, 1999; pp. 1–315. ISBN 978-84-605-9994-4. [Google Scholar]
- Marvaldi, A.E.; Lanteri, A.A.; del Río, M.G.; Oberprieler, R.G. 3.7.5. Entiminae Schoenherr, 1823. In Handbook of Zoology. Coleoptera, Beetles. Morphology and Systematics; Leschen, R.A., Beutel, R.G., Eds.; De Gruyter: Berlin, Germany; Boston, MI, USA, 2014; Volume 3, pp. 503–522. [Google Scholar]
- Marvaldi, A.E. Higher Level Phylogeny of Curculionidae (Coleoptera: Curculionoidea) based mainly on Larval Characters, with Special Reference to Broad-Nosed Weevils. Cladistics 1997, 13, 285–312. [Google Scholar] [CrossRef]
- Hundsdoerfer, A.K.; Rheinheimer, J.; Wink, M. Towards the phylogeny of the Curculionoidea (Coleoptera): Reconstructions from mitochondrial and nuclear ribosomal DNA sequences. Zool. Anz. 2009, 248, 9–31. [Google Scholar] [CrossRef]
- McKenna, D.D.; Sequeira, A.S.; Marvaldi, A.E.; Farrell, B.D. Temporal lags and overlap in the diversification of weevils and flowering plants. Proc. Natl. Acad. Sci. USA 2009, 106, 7083–7088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillett, C.P.; Crampton-Platt, A.; Timmermans, M.J.; Jordal, B.H.; Emerson, B.C.; Vogler, A.P. Bulk de novo mitogenome assembly from pooled total DNA elucidates the phylogeny of weevils (Coleoptera: Curculionoidea). Mol. Biol. Evol. 2014, 31, 2223–2237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillett, C.P.; Lyal, C.H.; Vogler, A.P.; Emerson, B.C. Statistical Evaluation of Monophyly in the ‘Broad-Nosed Weevils’ through Molecular Phylogenetic Analysis Combining Mitochondrial Genome and Single-Locus Sequences (Curculionidae: Entiminae, Cyclominae, and Hyperinae). Diversity 2018, 10, 21. [Google Scholar] [CrossRef]
- Gunter, N.L.; Oberprieler, R.G.; Cameron, S.L. Molecular phylogenetics of Australian weevils (Coleoptera: Curculionoidea): Exploring relationships in a hyperdiverse lineage through comparison of independent analyses. Austral Entomol. 2016, 55, 217–233. [Google Scholar] [CrossRef]
- Shin, S.; Clarke, D.J.; Lemmon, A.R.; Moriarty Lemmon, E.; Aitken, A.L.; Haddad, S.; Farrell, B.D.; Marvaldi, A.E.; Oberprieler, R.G.; McKenna, D.D. Phylogenomic Data Yield New and Robust Insights into the Phylogeny and Evolution of Weevils. Mol. Biol. Evol. 2018, 35, 823–836. [Google Scholar] [CrossRef] [PubMed]
- Haran, J.; Timmermans, M.J.; Vogler, A.P. Mitogenome sequences stabilize the phylogenetics of weevils (Curculionoidea) and establish the monophyly of larval ectophagy. Mol. Phylogenet. Evol. 2013, 67, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Oberprieler, R.G. 3.7.4 Cyclominae Schoenherr, 1826. In Handbook of Zoology. Coleoptera, Beetles. Morphology and Systematics; Leschen, R.A., Beutel, R.G., Eds.; De Gruyter: Berlin, Germany; Boston, MI, USA, 2014; Volume 3, pp. 483–502. [Google Scholar]
- Gaiger, F. Systematic revision and cladistic analysis of the genus Rhigus Schoenherr, 1823 (Coleoptera, Curculionidae). Rev. Bras. Entomol. 2001, 45, 43–85. [Google Scholar]
- Vanin, S.A.; Gaiger, F. A cladistic analysis of the genera of the tribe Entimini (Coleoptera, Curculionidae), with description of a new genus and species from the Amazonian region. Zootaxa 2005, 1053, 1–21. [Google Scholar]
- Gaiger, F.; Vanin, S.A. On the utility of female genitalia in the systematics of Curculionidae: Examples from Entimini and Hylobiina (Coleoptera: Curculionidae). Ann. Soc. Entomol. Fr. 2008, 44, 47–57. [Google Scholar] [CrossRef]
- Franz, N.M. Redescriptions of critical type species in the Eustylini Lacordaire (Coleoptera: Curculionidae: Entiminae). J. Nat. Hist. 2010, 44, 41–80. [Google Scholar] [CrossRef]
- Franz, N.M. Revision and phylogeny of the Caribbean weevil genus Apotomoderes Dejean, 1834 (Coleoptera, Curculionidae, Entiminae). ZooKeys 2010, 49, 33–75. [Google Scholar] [CrossRef]
- Franz, N.M. Phylogenetic reassessment of the Exophthalmus genus complex (Curculionidae: Entiminae: Eustylini, Geonemini). Zool. J. Linn. Soc. 2012, 164, 510–557. [Google Scholar] [CrossRef]
- Girón, J.C.; Franz, N.M. Revision, phylogeny and historical biogeography of the genus Apodrosus Marshall, 1922 (Coleoptera: Curculionidae: Entiminae). Insect Syst. Evol. 2010, 41, 339–414. [Google Scholar] [CrossRef]
- Girón, J.C.; Franz, N.M. Phylogenetic assessment of the Caribbean weevil genus Lachnopus Schoenherr (Coleoptera: Curculionidae: Entiminae). Invertebr. Syst. 2012, 26, 67–82. [Google Scholar] [CrossRef]
- Howden, A.T. The Pandeleteius subgenus Exmenetypus Voss in Central America (Coleoptera: Curculionidae, Tanymecini). Coleopt. Bull. 1974, 28, 7–16. [Google Scholar]
- Howden, A.T. Revision of the New World genus Hadromeropsis Pierce (Coleoptera, Curculionidae, Tanymecini). Contrib. Am. Entomol. Inst. 1982, 19, 1–180. [Google Scholar]
- Howden, A.T. Revision of the Mesoamerican species of Airosimus Howden (Coleoptera: Curculionidae). Coleopt. Bull. 1993, 47, 249–267. [Google Scholar]
- Howden, A.T. The species of Pandeleteius Schoenherr of coastal Chile and Peru (Coleoptera, Curculionidae). Zootaxa 2008, 1773, 55–62. [Google Scholar]
- Howden, A.T. On some species of Pandeleteius Schoenherr, 1834, in South America south of the tenth parallel (Coleoptera, Curculionidae: Entiminae: Tanymecini). Zootaxa 2011, 2977, 50–60. [Google Scholar]
- Scataglini, M.A.; Lanteri, A.A.; Confalonieri, V.A. Phylogeny of the Pantomorus–Naupactus complex based on morphological and molecular data (Coleoptera: Curculionidae). Cladistics 2005, 21, 131–142. [Google Scholar] [CrossRef]
- Rosas, M.V.; Morrone, J.J.; del Río, M.G.; Lanteri, A.A. Phylogenetic analysis of the Pantomorus-Naupactus complex (Coleoptera: Curculionidae: Entiminae) from North and Central America. Zootaxa 2011, 2780, 1–19. [Google Scholar]
- Lanteri, A.A.; del Río, M.G. Phylogeny of the tribe Naupactini (Coleoptera: Curculionidae) based on morphological characters. Syst. Entomol. 2017, 42, 429–447. [Google Scholar] [CrossRef]
- del Río, M.G.; Rodriguero, M.S.; Confalonieri, V.A.; Lanteri, A.A. Molecular and Morphological Phylogenetic Analysis of Naupactus Dejean (Curculionidae: Entiminae) and Allied Genera: The Dilemma of Classification. Diversity 2018, 10, 59. [Google Scholar] [CrossRef]
- Velazquez De Castro, A.J.; Alonso-Zarazaga, M.Á.; Outerelo, R. Systematics of Sitonini (Coleoptera: Curculionidae: Entiminae), with a hypothesis on the evolution of feeding habits. Syst. Entomol. 2007, 32, 312–331. [Google Scholar] [CrossRef]
- Korotyaev, B.A.; Meleshko, J.Y. On the systematics of the weevil genus Polydrusus (Coleoptera: Curculionidae). Zoosyst. Ross. 1997, 6, 275–286. [Google Scholar]
- Morimoto, K.; Kojima, H.; Miyakawa, S. The Insects of Japan. Vol. 3. Curculionoidea: General Introduction and Curculionidae: Entiminae (Part 1), Phyllobiini, Polydrusini and Cuphicerini (Coleoptera); Entomological Society of Japan, Touka Shobo Co. Ltd.: Fukuoka, Japan, 2006. [Google Scholar]
- Machado, A. El género Drouetius Méquignon, 1942 stat. prom., de las islas Azores (Coleoptera, Curculionidae, Entiminae). Graellsia 2009, 65, 19–46. [Google Scholar] [CrossRef]
- Machado, A. Revisión de los antiguos Lichenophagus (= Laparocerus) de la Macaronesia (Coleoptera, Curculionidae, Entiminae). Bol. SEA 2015, 57, 93–141. [Google Scholar]
- Machado, A.; Lopez, M.; Almeida, T.; Hernandez, M. Mitochondrial DNA phylogenetic analysis of the genus Laparocerus (Coleoptera, Curculionidae, Entiminae). I. The Madeiran clade. Zool. Scr. 2008, 37, 415–427. [Google Scholar] [CrossRef]
- Machado, A.; Rodríguez-Expósito, E.; López, M.; Hernández, M. Phylogenetic analysis of the genus Laparocerus, with comments on colonisation and diversification in Macaronesia (Coleoptera, Curculionidae, Entiminae). ZooKeys 2017, 1651, 1–77. [Google Scholar] [CrossRef] [PubMed]
- Yunakov, N.N. Species of the Subgenus Pseudocryphiphorus, Genus Otiorhynchus (Coleoptera, Curculionidae), in the Fauna of the Crimea. Entomol. Rev. 2003, 83, 221–229. [Google Scholar]
- Yunakov, N.N. On the taxonomy of the weevil genus Parameira Seidl. (Coleoptera, Curculionidae, Entiminae). Zool. Zhurnal 2004, 83, 1284–1289. [Google Scholar]
- Yunakov, N.N.; Korotyaev, B.A. On the systematics of the weevil genus Araxia Khnzorian with descriptions of new taxa from Transcaucasia, Turkey, and Turkmenistan (Coleoptera: Curculionidae: Entiminae). Insect Syst. Evol. 2008, 39, 39–54. [Google Scholar] [CrossRef]
- Yunakov, N.N.; Klass, K.D. Contribution to the taxonomy and nomenclature of Entiminae from the Palearctic region and South Africa (Coleoptera: Curculionidae). Beitr. Entomol. 2012, 62, 427–445. [Google Scholar] [CrossRef]
- Oberprieler, R. Revision of the Tanyrhynchini of continental Africa (Coleoptera, Curculionidae) I. Introduction and review of the genera, revision of the genus Brachytrachelus Schönherr and description of Afroleptops gen. nov. Entomol. Mem. Dep. Agric. Water Supply Repub. S. Afr. 1988, 71, 1–50. [Google Scholar]
- Oberprieler, R.G. Systematic position and composition of the tribes Tanyrhynchini and Myorhinini (Coleoptera: Curculionidae). Mem. Entomol. Soc. Wash. 1995, 14, 155–167. [Google Scholar]
- Thompson, R.T. Revision of the genus Catasarcus Schonherr (Coleoptera: Curculionidae). Bull. Br. Mus. (Nat. Hist.) Entomol. 1968, 22, 357–454. [Google Scholar]
- Posadas, P.E. Los Curculionoidea (Insecta: Coleoptera) de la Tierra del Fuego e Islas Malvinas. Ph.D. Thesis, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, La Plata, Argentina, 2002. [Google Scholar]
- Elgueta, M. Geosphaeropterus, nuevo género de Tropiphorini (Coleoptera: Curculionidae) de Chile, con descripción de tres nuevas especies. Bol. Mus. Nac. Hist. Nat. Chile 2013, 62, 203–217. [Google Scholar]
- Brown, S.D.J. Taxonomy and Evolution of Endemic New Zealand Broad-Nosed Weevils (Coleoptera: Curculionidae: Entiminae). Ph.D. Thesis, Lincoln University, Christchurch, New Zealand, 2016. [Google Scholar]
- Borovec, R. Revision of the Palaearctic supraspecific taxa of the tribe Trachyphloeini (Coleoptera: Curculionidae: Entiminae). Klapalekiana 2009, 45, 1–97. [Google Scholar]
- Kuschel, G. Terrestrial zoology in southern Chile. Proc. R. Soc. Lond. B 1960, 152, 540–550. [Google Scholar] [CrossRef]
- Kuschel, G. The Curculionidae of Gough Island and the relationships of the weevil fauna of the Tristan da Cunha group. Proc. Linn. Soc. Lond. 1962, 173, 69–78. [Google Scholar] [CrossRef]
- Kuschel, G. Biogeography and ecology of South American Coleoptera. In Biogeography and Ecology of South America; Fittkau, E.J., Illies, J., Klinge, H., Schwabe, G.H., Eds.; Monographiae Biologicae; Springer: The Hague, Switzerland, 1969; pp. 709–722. [Google Scholar]
- Morrone, J.J.; Roig-Juñent, S. Synopsis and cladistics of the American Aterpini (Coleoptera: Curculionidae, Cyclominae). Insect Syst. Evol. 2000, 30, 417–434. [Google Scholar] [CrossRef]
- Morrone, J.J. The subtribes and genera of the tribe Listroderini (Coleoptera, Curculionidae, Cyclominae): Phylogenetic analysis with systematic and biogeographical accounts. ZooKeys 2013, 273, 15–71. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Zarazaga, M.A.; Barrios, H.; Borovec, R.; Bouchard, P.; Caldara, R.; Colonnelli, E.; Gültekin, L.; Hlaváč, P.; Korotyaev, B.; Lyal, C.H. Cooperative catalogue of Palaearctic Coleoptera Curculionoidea. Monogr. Electrón. SEA 2017, 8, 1–729. [Google Scholar]
- Grebennikov, V.V. Neglected Trichalophus (Coleoptera: Curculionidae): DNA barcode and phylogeography of high-altitude flightless weevils rediscovered in Southwest China. Bonn. Zool. Bull. 2016, 64, 59–76. [Google Scholar]
- Pullen, K.R.; Jennings, D.; Oberprieler, R.G. Annotated catalogue of Australian weevils (Coleoptera: Curculionoidea). Zootaxa 2014, 3896, 1–481. [Google Scholar] [CrossRef] [PubMed]
- Morrone, J.J. Biogeographical regionalisation of the world: A reappraisal. Aust. Syst. Bot. 2015, 28, 81–90. [Google Scholar] [CrossRef]
- Kuschel, G. Nuevos Brachyderinae y Magdalinae chilenos (Coleoptera Curculionidae). Arthropoda 1950, 1, 181–195. [Google Scholar]
- Kuschel, G. The Curculionidae of the Cordillera Chileno-Argentina (1a. parte) (Aporte 13 Coleoptera Curculionidae). Rev. Chil. Entomol. 1952, 2, 271–273. [Google Scholar]
- Sunnucks, P.; Hales, D.F. Numerous transposed sequences of mitochondrial cytochrome oxidase I-II in aphids of the genus Sitobion (Hemiptera: Aphididae). Mol. Biol. Evol. 1996, 13, 510–524. [Google Scholar] [CrossRef] [PubMed]
- Duckett, C.N.; Gillespie, J.J.; Kjer, K.M. Relationships among the subfamilies of Chrysomelidae inferred from small subunit ribosomal DNA and morphology with special emphasis on the relationship among the flea beetles and the Galerucinae. In New Developments in the Biology of Chrysomelidae; Jolivet, P., Santiago-Blay, J.A., Schmitt, M., Eds.; SPB Academic Publishing BV.: The Hague, The Netherlands, 2004; pp. 3–18. [Google Scholar]
- Brown, S.D.J.; Armstrong, K.F.; Cruickshank, R.H. Molecular phylogenetics of a South Pacific sap beetle species complex (Carpophilus spp., Coleoptera: Nitidulidae). Mol. Phylogenet. Evol. 2012, 64, 428–440. [Google Scholar] [CrossRef] [PubMed]
- Filatov, D.A. ProSeq: A software for preparation and evolutionary analysis of DNA sequence data sets. Mol. Ecol. Notes 2002, 2, 621–624. [Google Scholar] [CrossRef]
- Kjer, K.M.; Roshan, U.; Gillespie, J.J. Structural and evolutionary considerations for multiple sequence alignment of RNA, and the challenges for algorithms that ignore them. In Sequence Alignment: Methods, Models, Concepts, and Strategies; Rosenberg, M.S., Ed.; University of California Press: Berkeley, CA, USA, 2009; 337p. [Google Scholar]
- Gillespie, J.J.; Johnston, J.S.; Cannone, J.J.; Gutell, R.R. Characteristics of the nuclear (18S, 58 S, 28S and 5S) and mitochondrial (12S and 16S) rRNA genes of Apis mellifera (Insecta: Hymenoptera): Structure, organization, and retrotransposable elements. Insect Mol. Biol. 2006, 15, 657–686. [Google Scholar] [CrossRef] [PubMed]
- Marvaldi, A.E.; Duckett, C.N.; Kjer, K.M.; Gillespie, J.J. Structural alignment of 18S and 28S rDNA sequences provides insights into phylogeny of Phytophaga (Coleoptera: Curculionoidea and Chrysomeloidea). Zool. Scr. 2009, 38, 63–77. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP* Phylogeny Analysis Using Parsimony (* and Other Methods); Version 40b10; Sinauer Assoc. Inc.: Sunderland, MA, USA, 2002. [Google Scholar]
- Halanych, K.M.; Lutz, R.A.; Vrijenhoek, R.C. Evolutionary origins and age of vestimentiferan tube-worms. Cah. Biol. Mar. 1998, 39, 355–358. [Google Scholar]
- Sequeira, A.S.; Lanteri, A.A.; Scataglini, M.A.; Confalonieri, V.A.; Farrell, B.D. Are flightless Galapaganus weevils older than the Galápagos Islands they inhabit? Heredity 2000, 85, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Dowton, M.; Austin, A.D. Phylogenetic relationships among the microgastroid wasps (Hymenoptera: Braconidae): Combined analysis of 16S and 28S rDNA genes and morphological data. Mol. Phylogenet. Evol. 1998, 10, 354–366. [Google Scholar] [CrossRef] [PubMed]
- Whiting, M.F.; Carpenter, J.C.; Wheeler, Q.D.; Wheeler, W.C. The Strepsiptera problem: Phylogeny of the holometabolous insect orders inferred from 18S and 28S ribosomal DNA sequences and morphology. Syst. Biol. 1997, 46, 1–68. [Google Scholar] [CrossRef] [PubMed]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Lanteri, A.A.; O’Brien, C.W. Taxonomic revision and cladistic analysis of Atrichonotus Buchanan (Coleoptera: Curculionidae). Trans. Am. Entomol. Soc. 1990, 116, 697–725. [Google Scholar]
- Marvaldi, A.E. Larvae of South American Entimini (Coleoptera: Curculionidae), and phylogenetics implications of certain characters. Rev. Chil. Entomol. 1998, 25, 21–44. [Google Scholar]
- Marvaldi, A.E. Larvae of South American Rhytirrhininae (Coleoptera: Curculionidae). Coleopt. Bull. 1998, 52, 71–89. [Google Scholar]
- Morrone, J.J.; Diaz, N.B.; Loiacono, M.S. Comparative morphology of mouthparts in the tribe Listroderini (Coleoptera: Curculionidae). Elytron 1992, 6, 47–59. [Google Scholar]
- Marvaldi, A.E.; Lanteri, A.A. Key to higher taxa of South American weevils based on adult characters (Coleoptera: Curculionoidea). Rev. Chil. Hist. Nat. 2005, 78, 65–87. [Google Scholar] [CrossRef]
- Buchanan, L.L.F. The Species of Pantomorus of America North of Mexico; U.S. Department Agriculture: Washinton, DC, USA, 1939; Volume 341, pp. 1–39.
- Marvaldi, A.E. Larvae of Entiminae (Coleoptera: Curculionidae): Tribal diagnoses and phylogenetic key, with a proposal about natural groups within Entimini. Insect Syst. Evol. 1998, 29, 89–98. [Google Scholar] [CrossRef]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.H.; Xie, D.; Suchard, M.A.; Drummond, A.J. BEAST 2: A Software Platform for Bayesian Evolutionary Analysis. PLoS Comput. Biol. 2014, 10, e1002537. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffe, W.; Schwartz, T. Creating the CIPRES Science Gate way for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010. [Google Scholar]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer Science & Business Media: New York, NY, USA, 2002. [Google Scholar]
- Drumond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A. FigTree v. 1.3.1. Computer Program and Documentation Distributed by the Author, 2006–2012. Available online: http://tree.bio.ed.ac.uk/software/ (accessed on 21 December 2017).
- Goloboff, P.A.; Catalano, S. TNT version 1.5, including full implementation of phylogenetic morphometrics. Cladistics 2016, 32, 221–238. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Kluge, A.G.; Farris, J.S. Quantitative phyletics and the evolution of anurans. Syst. Biol. 1969, 18, 1–32. [Google Scholar] [CrossRef]
- Farris, J.S. The retention index and the rescaled consistency index. Cladistics 1989, 5, 417–419. [Google Scholar] [CrossRef]
- Nixon, K.C. WinClada, ver. 1.00.08; Nixon, K.C.: Ithaca, NY, USA, 2002. [Google Scholar]
- Oberprieler, R. A reclassification of the weevil subfamily Cyclominae (Coleoptera: Curculionidae). Zootaxa 2010, 2515, 1–35. [Google Scholar]
- Meregalli, M.; Germann, C.; Bernasconi, M.V.; Cervella, P. Phylogeny of the Genus Dichotrachelus (Coleoptera: Curculionidae: Cyclominae). Diversity 2018, 10, 66. [Google Scholar] [CrossRef]
- Ashworth, A.C.; Kuschel, G. Fossil weevils (Coleoptera: Curculionidae) from latitude 85° S Antarctica. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003, 191, 191–202. [Google Scholar] [CrossRef]
- Kuschel, G. La subfamilia Aterpinae en América. Rev Chil. Entomol. 1951, 1, 205–244. [Google Scholar]
- Morrone, J.J. Distributional pattern of South American Aterpini (Coleoptera: Curculionidae). Rev. Soc. Entomol. Argent. 1996, 55, 131–141. [Google Scholar]
- Posadas, P.; Morrone, J.J. Biogeografía histórica de la familia Curculionidae (Coleoptera) en las subregiones Subantártica y Chilena Central. Rev. Soc. Entomol. Argent. 2003, 62, 75–84. [Google Scholar]
- Kuschel, G. Los “Curculionidae” del extremo Norte de Chile (Coleoptera, Curcul. Ap. 6). Acta Zool. Lilloana 1949, 8, 5–54. [Google Scholar]
- Kuschel, G. Entomology of the Aucklands and other islands south of New Zealand: (Coleoptera: Curculionidae). Pac. Insects Monogr. 1971, 27, 225–259. [Google Scholar]
- Morrone, J.J. Distributional patterns of species of Rhytirrhinini (Coleoptera: Curculionidae) and the historical relationships of the Andean provinces. Glob. Ecol. Biogeogr. Lett. 1994, 4, 188–194. [Google Scholar] [CrossRef]
- Morrone, J.J. Cladistics of the new world genera of Listroderina (Coleoptera: Curculionidae: Rhytirrhinini). Cladistics 1997, 13, 247–266. [Google Scholar] [CrossRef]
- Posadas, P. A preliminar overview of species composition and geographical distribution of Malvinian weevils (Insecta: Coleoptera: Curculionidae). Zootaxa 2008, 1704, 1–26. [Google Scholar]
- Posadas, P. Species composition and geographic distribution of Fuegian Curculionidae (Coleoptera: Curculionoidea). Zootaxa 2012, 3303, 1–36. [Google Scholar]
- Morrone, J.J.; Loiácono, M.S. Revision of the genus Rhigopsidius Heller (Insecta, Coleoptera, Curculionidae: Rhytirrhininae). Entomol. Abh 1992, 54, 129–139. [Google Scholar]
- Hustache, A. Contribution à l’étude des Curculionides de la République Argentine. An. Mus. Nac. Hist. Nat. Bernard. Rivadavia 1926, 34, 155–261. [Google Scholar]
- Schenkling, S.; Marshall, G.A.K. Coleopterorum Catalogus; Schenkling, S., Ed.; W. Junk: Berlín, Germany, 1931; Volume 28, Pars 114. Curculionidae: Eremninae, pp. 3–39; Leptopinae, pp. 1–83; Tanyrrhinchinae, pp. 1–10; Cylydrorrhininae, pp. 1–23; Thecesterninae (Suppl.), p. 1; Rhytitthininae (Suppl.), pp. 1–4; Ryparosominae (Suppl.) p. 4. [Google Scholar]
- Voss, E. Curculionidae (Col.). In Beiträge zur Fauna Perus; Titschack, E., Ed.; Gustav Fischer Verlag: Jena, Germany, 1954; Volume 4, pp. 163–376. [Google Scholar]
- Kuschel, G. Nuevos Cylydrorhininae de la Patagonia (Col. Curculionoidea, Aporte 18). Investig. Zool. Chil. 1958, 4, 231–252. [Google Scholar]
- Kuschel, G. Revisión de los Premnotrypini y adiciones a los Bagoini (Aporte 17 sobre Coleoptera Curculionoidea). Bol. Mus. Nac. Hist. Nat. Chile 1956, 26, 187–235. [Google Scholar]
- Alcázar, J.; Cisneros, F. Taxonomy and bionomics of the Andean potato weevil complex: Premnotrypes spp. and related genera. CIP Program Rep. 1997, 98, 141–151. [Google Scholar]
- Elgueta, M. Descripción de una nueva especie de Hybreoleptops Kuschel 1949 y comentarios sobre otros representantes del género (Insecta: Coleoptera: Curculionidae). Bol. Mus. Nac. Hist. Nat. Chile 2012, 61, 29–42. [Google Scholar]
- Pérez, M.E.; Posadas, P. Cladistics and redescription of Hybreoleptops Kuschel (Coleoptera: Curculionidae: Entiminae) with the description of two new species from the Central Chilean subregion. J. Nat. Hist. 2006, 40, 1775–1791. [Google Scholar] [CrossRef]
- Alonso-Zarazaga, M.A.; Perrin, H.; Sanchez-Ruiz, M. Sur l’identité du genre Ctenolobus Desbrochers des Loges, 1892 (Coleoptera, Curculionidae). Bull. Société Entomol. Fr. 2010, 115, 479–483. [Google Scholar]
- del Río, M.G.; Morrone, J.J.; Lanteri, A.A. Evolutionary biogeography of South American weevils of the tribe Naupactini (Coleoptera: Curculionidae). J. Biogeogr. 2015, 42, 1293–1304. [Google Scholar] [CrossRef]
- Vaurie, P. Revision of the genus Entimus with notes on other genera of Entimini (Coleoptera Curculionidae). Rev. Chil. Entomol. 1952, 1, 147–171. [Google Scholar]
- Vanin, S.A. Revision of Phaedropus Schoenherr with notes on the male genitalia of Entimini (Coleoptera: Curculionidae: Leptopiinae). Coleopt. Bull. 1983, 37, 283–298. [Google Scholar]
- Warner, R.E. The genus Eudiagogus (Coleoptera: Curculionidae: Leptopiinae), with two new species on the weed Sesbania (Leguminosae). Proc. Entomol. Soc. Wash. 1979, 81, 304–320. [Google Scholar]
- O’Brien, C.W.; Kovarik, P.W. A new species of Eudiagogus Schoenherr from South America (Coleoptera: Curculionidae). Coleopt. Bull. 2000, 54, 313–317. [Google Scholar] [CrossRef]
- Girón, J.C.; O’Brien, C.W.; Rose-Smyth, M.C. On the West Indian weevil genus Lachnopus Schönherr, 1840 (Coleoptera: Curculionidae: Entiminae): Descriptions of six new species, a proposal for species-groups, and an annotated checklist. Zootaxa 2018, 4423, 1–85. [Google Scholar] [CrossRef]
- Zhang, G.; Basharat, U.; Matzke, N.; Franz, N.M. Model selection in statistical historical biogeography of Neotropical insects—The Exophthalmus genus complex (Curculionidae: Entiminae). Mol. Phylogenet. Evol. 2017, 109, 226–239. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.S.; Lanteri, A.A. New genera and species of weevils from the Galapagos Islands, Ecuador, and Cocos Island, Costa Rica (Coleoptera; Curculionidae; Entiminae; Entimini). Am. Mus. Novit. 2000, 3299, 1–15. [Google Scholar] [CrossRef]
- Van Emden, F.I. On the taxonomy of Rhynchophora larvae: Adelognatha and Alophinae (Insecta: Coleoptera). Proc. Zool. Soc. Lond. 1952, 122, 651–795. [Google Scholar] [CrossRef]
- Jackson, S.D. Observaciones sobre la biologia de Platyaspistes glaucus Fahraeus, 1840 (Coleoptera: Curculionidae). Rev. Chil. Entomol. 1987, 15, 85–87. [Google Scholar]
- Marvaldi, A.E. Eggs and oviposition habits in Entimini (Coleoptera: Curculionidae). Coleopt. Bull. 1999, 53, 115–126. [Google Scholar]
- Marshall, G.A.K. The Otiorrhynchine Curculionidae of the Tribe Celeuthetini (Col.); Trustees of the British Museum: London, UK, 1956; pp. 1–134. [Google Scholar]
- Toussaint, E.F.; Tänzler, R.; Rahmadi, C.; Balke, M.; Riedel, A. Biogeography of Australasian flightless weevils (Curculionidae, Celeuthetini) suggests permeability of Lydekker’s and Wallace’s Lines. Zool. Scr. 2015, 44, 632–644. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tribe | Species | Acronyms | Locality | Accesion Codes | |||
|---|---|---|---|---|---|---|---|
| 18S | 28S | 16S | COI-5P | ||||
| *Aterpini | Aegorhinus silvicola Kuschel | CYC_Aegorhinus_sil | Argentina, Neuquén, San Martín de los Andes | MH746278 | MH746328 | MH746378 | MH734198 |
| Aegorhinus vitulus (Fabricius) | CYC_Aegorhinus_vit | Argentina, Tierra del Fuego, Ushuaia | MH746279 | - | MH746379 | MH734199 | |
| *Listroderini | Listroderes victus Germain | CYC_Listroderes_vic | Chile, Cartagena, El Tabo, coastal dunes | MH746280 | MH746329 | MH746380 | MH734200 |
| Rhigopsidius piercei Heller | CYC_Rhigopsidius_pie | Argentina, Jujuy | MH746281 | MH746330 | MH746381 | MH734201 | |
| Cylydrorhinini | Caneorhinus biangulatus (Champion) | CYL_Caneorhinus_bia | GenBank source (Arg. Falklands Is.) | - | - | EF213994 | - |
| Cylydrorhinus caudiculatus (Fairmaire) | CYL_Cylydrorhinus_cau | GenBank source (Arg. Falklans Is.) | - | - | EF214094 | - | |
| Cylydrorhinus chilensis (Blanchard) | CYL_Cylydrorhinus_chi | Argentina, La Rioja, Dto. Vinchina | MH746286 | MH746335 | - | MH734202 | |
| Cylydrorhinus aff. patagonicus (Kuschel) | CYL_Cylydrorhinus_pat | Argentina, Jujuy, Dto. Susques | MH746287 | MH746336 | MH746385 | MH734203 | |
| Premnotrypini | Premnotrypes latithorax (Pierce) | PRE_Premnotrypes_lat | Perú, Cuzco | MH746316 | MH746365 | MH746416 | MH734204 |
| Leptopiini | Catasarcus impresipennis (Boisduval) | LEP_Catasarcus_imp | GenBank source (Australia) | - | KF016401 | KF016266 | - |
| Catasarcus sp. | LEP_Catasarcus_sp | GenBank source (Australia) | FJ867744 | FJ867671 | AJ495489 | FJ867847 | |
| Cecyropa sp. | LEP_Cecyropa_sp | New Zealand, Waikuku Beach, Surf Club | MH746322 | MH746371 | - | MH734205 | |
| Irenimus sp. | LEP_Irenimus_sp | New Zealand, Ahuriri Scenic Reserve, Port Hills, Christchurch | MH746324 | MH746373 | MH746422 | MH734206 | |
| Strangaliodes deserticola Kuschel | LEP_Strangaliodes_des | Chile, Atacama | MH746325 | MH746374 | - | MH734207 | |
| Strangaliodes sp. (aff. deserticola) | LEP_Strangaliodes_sp | Chile, Pichilemos | - | MH746375 | MH746423 | MH734208 | |
| Strangaliodes niger (Blanchard) | LEP_Strangaliodes_nig | Argentina, San Juan | MH746326 | MH746376 | MH746424 | MH734209 | |
| Vossius nebulosus (Gyllenhal) | LEP_Vossius_neb | Argentina, Misiones, on way to Salto Tabai, close to Jardín America | MH746323 | MH746372 | - | MH734210 | |
| Entimini | Entimus sp. | ENT_Entimus_sp | Brazil, Rio de Janeiro, Coastal Restingas | MH746327 | MH746377 | MH746425 | MH734211 |
| Eudiagogini | Eudiagogus rosenschoeldi Fahraeus | EUD_Eudiagogus_ros | GenBank source (United States of America) | AF250081 | - | - | - |
| Pororhynchus aff. albolateralis Hustache | EUD_Pororhynchus_alb | Argentina, San Juan, near Telteca | MH746288 | MH746337 | MH746387 | MH734212 | |
| Eustylini | Compsus sp. | EUS_Compsus_sp | GenBank source (South America) | - | KT350862 | - | KT350650 |
| Diaprepes abbreviatus (Linnaeus) | EUS_Diaprepes_abb | GenBank source (Central America) | AY157729 | KT350865 | CN475651 | HQ891432 | |
| Galapagonotus cuneiformis Waterhouse | Galapagonotus_cun | Ecuador, Islas Galápagos, San Cristóbal, El Junco | MH746282 | MH746331 | MH746386 | - | |
| Oxyderces bimaculatus (Hustache) | EUS_Oxyderces_bim | Argentina, Corrientes, Empedrado | MH746289 | MH746338 | MH746388 | MH734213 | |
| Geonemini | Artipus sp. | GEO_Artipus sp. | GenBank source (Caribbean) | - | KT350859 | KP253881 | HQ891428 |
| Epicaerus sp | GEO_Epicaerus_sp | United States of America | MH746290 | MH746339 | MH746389 | MH734214 | |
| Lachnopus sp. | GEO_Lachnopus_sp | GenBank source (Caribbean) | AF389052 | KT350910 | - | HQ891465 | |
| Naupactini | Atrichonotus taeniatulus (Berg) | NAU_Atrichonotus_tae | Argentina, Mendoza, Guaymallén | MH746296 | MH746345 | MH746395 | MH537919 |
| Hoplopactus lateralis Arrow | NAU_Hoplopactus_lat | Brazil, San Pablo | MH746297 | MH746346 | MH746396 | MH537920 | |
| Litostylus sp. | NAU_Litostylus_sp | GenBank source (Caribbean) | - | KT350916 | - | HQ891470 | |
| Naupactus auricinctus Boheman | NAU_Naupactus_ aur | Brazil, San Pablo | MH746298 | MH746347 | MH746397 | MH537921 | |
| Naupactus aff chloraspis Erichson | NAU_Naupactus_chl | Brazil, Bahia | MH746299 | MH746348 | MH746398 | MH734215 | |
| Naupactus aff faldermanni Boheman | NAU_Naupactus_ fald | Brazil, Mato Grosso | MH746300 | MH746350 | MH746399 | - | |
| Naupactus leucoloma Boheman | NAU_Naupactus_leu | Argentina, Mendoza | MH746301 | MH746349 | MH746400 | MH537922 | |
| Naupactus aff peruvianus Hustache | NAU_Naupactus_ per | Perú, Cuzco, Dto. Machu Pichu, Puerta Ciudad Antigua | MH746302 | MH746351 | MH746401 | - | |
| Naupactus sulphurifer Pascoe | NAU_Naupactus_sul | Argentina, Mendoza, Reserva Divisadero Largo | MH746303 | MH746352 | MH746402 | - | |
| Naupactus tarsalis Boheman | NAU_Naupactus_tar | Argentina, Formosa, Dto. Matacos, 20 Km Ing. Juárez | MH746304 | MH746353 | MH746403 | - | |
| Naupactus viridisquamosus Boheman | NAU_Naupactus_vir | Brazil, Mato Grosso | MH746305 | MH746354 | MH746404 | ||
| Naupactus xanthographus (Germar) | NAU_Naupactus_xan | Argentina, Mendoza [except 16S, Buenos Aires, La Plata] | FJ867775 | FJ867695 | MH746405 | NC0180354 | |
| Pantomorus ruizi (Brèthes) | NAU_Pantomorus_rui | Argentina, Chubut, Trelew INTA | MH746306 | MH746355 | MH746407 | MH537925 | |
| Symmathetes setarius (Boheman) | NAU_Symmathetes_seta | Brazil, Mato Grosso | MH746308 | MH746357 | MH746408 | MH537923 | |
| Symmathetes setulosus Hustache | NAU_Symmathetes_setu | Argentina, Catamarca, Las Esquinas, Dpto. Valle Viejo | MH746307 | MH746356 | MH746406 | MH537924 | |
| Tanymecini | Airosimus jacobi (Hustache) | TAN_Airosimus_jac | Argentina, Misiones, Urugua-i | MH746318 | MH746367 | MH746418 | MH734216 |
| Hadromeropsis aff. pallida Howden | TAN_Hadromeropsis_pal | Argentina, Entre Ríos, Cerrito | MH746319 | MH746368 | MH746419 | MH734217 | |
| Pandeleteius platensis Brèthes | TAN_Pandeleteius_pla | Argentina, Mendoza, Reserva Ñacuñán | MH746320 | MH746369 | MH746420 | MH734218 | |
| Pandeleteius sp. | TAN_Pandeleteius_sp | GenBank source (United States of America) | - | HQ891475 | - | HQ891475 | |
| Platyaspistes argentinensis Kuschel | TAN_Platyaspistes_arg | Argentina, San Juan, Dto. Sarmiento-Pedernal, Camino de los Berros | MH746321 | MH746370 | MH746421 | MH734219 | |
| Brachyderini | Brachyderes incanus Linnaeus | BRA_Brachyderes_inc | GenBank source (Europe) | - | KX551463 | AJ495503 | KJ962877 |
| Caulostrophus subsulcatus (Boheman) | BRA_Caulostrophus_sub | Italy, Genova, Voltri, Fiorino | MH746283 | MH746332 | MH746382 | MH734220 | |
| Strophosoma (Strophosoma) melanogrammum (Forster) | BRA_Strophosoma_ mel | Italy, Vercelli, Val Sesia, Monte Tovo | MH746284 | MH746333 | MH746383 | MH734221 | |
| Laparocerini | Laparocerus (Atlantis) noctivagans Wollaston | LAP_Laparocerus_noc | Portugal, Madeira, Rabaçal | MH746293 | MH746341 | MH746391 | MH734222 |
| Laparocerus (Fernandezius) subcalvus Wollaston | LAP_Laparocerus_sub | Spain, Islas Canarias, Isla El Hierro, NE San Andrés | MH746292 | MH746344 | MH746394 | - | |
| Laparocerus (Guanchotrox) canariensis Boheman | LAP_Laparocerus_can | Spain, Islas Canarias, Isla de Tenerife, El Portillo | MH746294 | MH746342 | MH746392 | MH734223 | |
| Laparocerus (Laparocerus) morio Boheman | LAP_Laparocerus_mor | Portugal, Madeira, Encumeada, | MH746295 | MH746343 | MH746393 | MH734224 | |
| Otiorhynchini | Dodecastichus mastix Olivier | OTI_Dodecastichus_mas | Italy, Pescara, La Maielletta | MH746309 | MH746358 | MH746409 | MH734225 |
| Otiorhynchus (Dorymerus) sulcatus Fabricius | OTI_Otiorhynchus_sul | GenBank source (Europe) | AF250084 | KU041875 | AJ495482/ JN163969 | KJ962464 | |
| Otiorhynchus (Metopiorrhynchus) teretirostris Stierling | OTI_Otiorhynchus_ter | Italy, Vercelli, Val Sesia, Monte Tovo | MH746310 | MH746359 | MH746410 | MH734226 | |
| Otiorhynchus (Provadilus) alpicola Boheman | OTI_Otiorhynchus_alp | Italy, Pescara, La Maielletta | MH746311 | MH746360 | MH746411 | MH734227 | |
| Peritelini | Drouetius azoricus parallelirostris Machado | PER_Drouetius_azo | Portugal, Islas Azores, Isla Terceira, Angra, Monte Brazil | MH746291 | MH746340 | MH746390 | MH734228 |
| Simo hirticornis Herbst | PER_Simo_hir | Italy, Lecco, Lombardia, Lagi di Annone | MH746312 | MH746361 | MH746412 | MH734229 | |
| Phyllobiini | Phyllobius (Parnemoicus) viridicollis Fabricius | PHY_Phyllobius_vir | Italy, Verona, Monti Lessini, Monte Castelberto | MH746313 | MH746362 | MH746413 | MH734230 |
| Polydrusini | Liophloeus (Liophloeus) tessulatus O.F. Müller | POL_Liophloeus_tes | Italy, Como, Lombardia, Piano del Tivano | MH746314 | MH746363 | MH746414 | MH734231 |
| Polydrusus (Eurodrusus) cervinus Linnaeus | POL_Polydrusus_cer | GenBank source (Europe) | - | HQ883568 | AJ495494 | HQ883653 | |
| Polydrusus (Eurodrusus) confluens Stephens | POL_Polydrusus_con | Italy, Como, Lombardia, Piano del Tivano | MH746315 | MH746364 | MH746415 | MH734232 | |
| Sciaphilini | Eusomus ovulum Germar | SCI_Eusomus_ovu | Italy, Milan, Zelo Buon Persico, Fiume Adda [except COI, GenBank source] | MH746317 | MH746366 | MH746417 | MH734233 |
| Sciaphilus asperatus Bonsdorff | SCI_Sciaphilus_asp | GenBank source (Europe) | JN619105 | - | AJ495502 | KJ962310 | |
| Celeuthetini | Celeuthetes sp. | CEL_Celeuthetes_sp | Vanuatu, Efate, Tagabe, Agr. Research Stn, Port Vila | MH746285 | MH746334 | MH746384 | MH734234 |
| Marker | Primer | Sequence (5′ > 3′) | Sense | Reference |
|---|---|---|---|---|
| 18S | E | CTG GTT GAT CCT GCC ACG T | F | [67] |
| R1138 | CGC CTT CGA ACC TCT AAC | R | [68] | |
| F1094 | GGA TCG TCG CAA GAC GGA CAG AAG | F | [68] | |
| P | TAA TGA TCC TTC CGC AGG TTC ACC T | R | [67] | |
| 18S a07 | ATT AAA GTT GTT GCG GTT | F | [60] | |
| 18S b25 | TCT TTG GCA AAT GCT TTC GC | R | [60] | |
| V7V9up | TCC GAT AAC GAA CGA GAC TC | F | [60] | |
| V7V9dwn | GTT ACG ACT TTT ACT TCC TC | R | [60] | |
| 28S | S3660 | GAG AGT TMA ASA GTA CGT GAA AC | F | [69] |
| A335 | TCG GAR GGA ACC AGC TAC TA | R | [70] | |
| 16S | N13398 | CGC CTG TTT AWC AAA AAC AT | F | [5] |
| J12887 | CTC CGG TYT GAA CTC AGA TCA AGT | R | [5] | |
| COI | LCO | GGT CAA CAA ATC ATA AAG ATA TTG G | F | [71] |
| HCO | TAA ACT TCA GGG TGA CCA AAA ATC A | R | [71] | |
| A3014 | TCC AAT GCA CTA ATC TGC CAT ATT A | R | [72] |
| 1 | Rostrum, length: more than 1.25 × as long as wide (minimum width, measured across apex without scrobes) (0) (Figure 1a); about as long as wide (slightly longer to slightly shorter) (1) (Figure 1b). |
| 2 | Rostrum, anterior portion of epistome: not raised and not projecting beyond anterior margin of rostrum (0); raised and projecting beyond anterior margin of rostrum (1) (Figure 1c). |
| 3 | Rostrum, outline of apex: slightly emarginated (0) (Figure 1a); V-shaped (1) (Figure 1b). |
| 4 | Rostrum, posterior part of epistome: denuded (0); covered with vestiture different from that of posterior part of rostrum (1) (Figure 5 in [28]). |
| 5 | Rostrum, expansion at apex (including pterigia): rostrum distinctly expanded at apex, ahead of antennal insertion (0) (Figure 1b); not to slightly expanded apex (sides subparallel or convergent towards apex) (1) (Figure 1d). |
| 6 | Rostrum, dorsal surface: impressed or flat (0) (Figure 1e); convex (1) (Figure 1c). |
| 7 | Rostrum, groove along midline: absent or indistinct (0); present (1) (Figure 1d). |
| 8 | Rostrum unicarinate or tricarinate (with central and lateral longitudinal carinae): not unicarinate or tricarinate (0); unicarinate or tricarinate (1) (Figure 1f). |
| 9 | Rostrum bicarinate (with pair of dorso-lateral carinae): not bicarinate (0); bicarinate (1) (Figure 1d). |
| 10 | Rostrum, scrobes in dorsal view: completely exposed, visible along their whole length (0) (Figure 3b); not completely exposed, only apical part partially visible (1) (Figure 3a). |
| 11 | Rostrum, sulcus along pterigia in lateral view: absent to slight (0); deep (1). |
| 12 | Rostrum, curvature of scrobes in lateral view: strongly curved ventrad of head (0) (Figure 1e); curved downwards and passing below eyes (1) (Figure 2a); slightly curved and orientated towards eyes (2) (Figure 2b). |
| 13 | Rostrum, scrobes in lateral view: well-defined and slightly widened posteriad (0) (Figure 2a); vaguely defined, shallow and very widened posteriad (1) (Figure 2b). |
| 14 | Rostrum, anteocular impression: longitudinal, latero-dorsal (0) (Figure 1 in [14]); triangular, lateral (1) (Figure 5 in [14]); indistinct (2). |
| 15 | Rostrum, transversal sulcus between rostrum and head (ventral view): present (0); indistinct (1). |
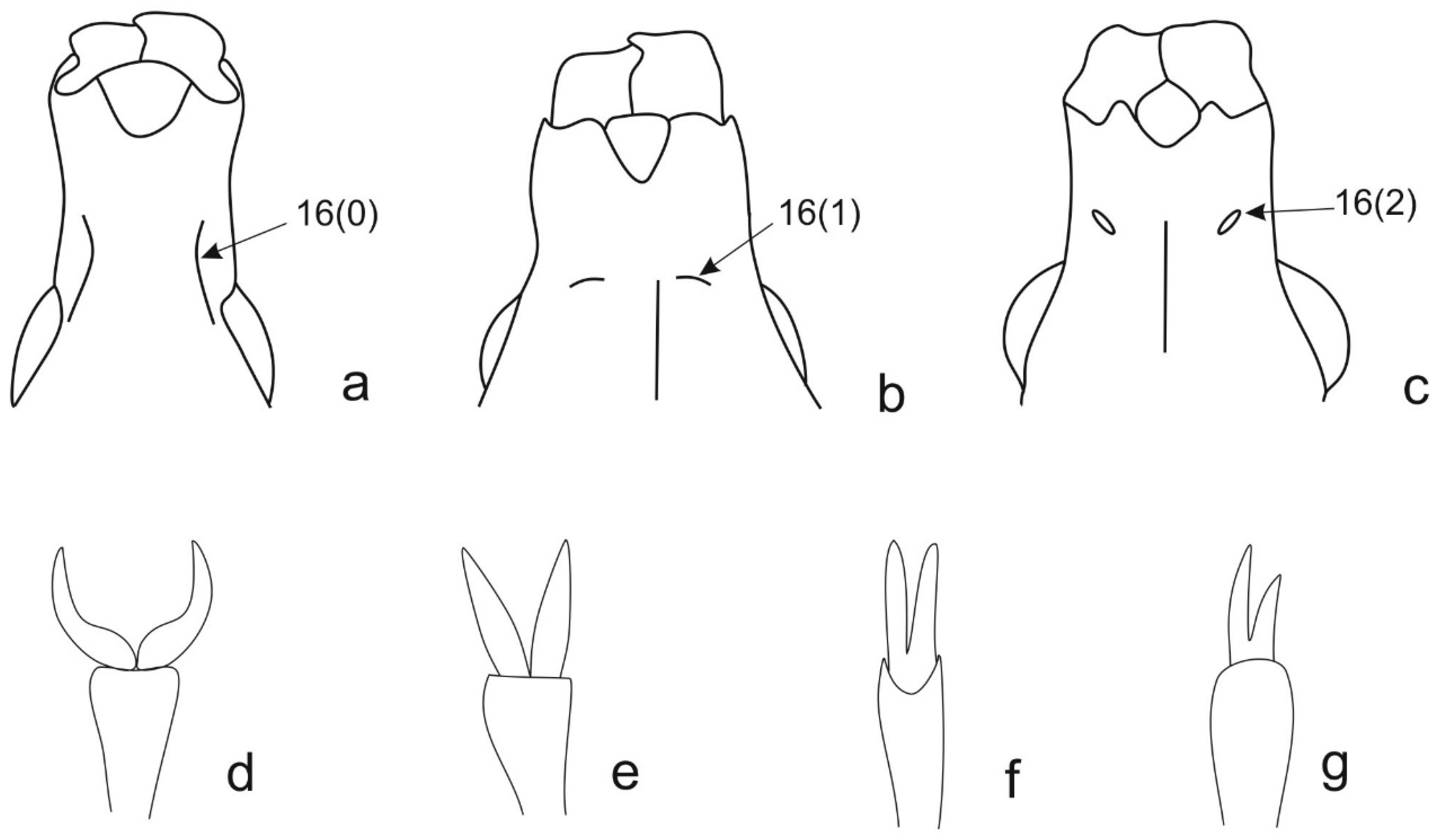
| 16 | Rostrum, occipital sutures (ventral view): longitudinal, curved, usually connected with lower edge of eye (0) (Figure 4a); transversal relative to longitudinal axis of rostrum, not connected with lower edge of eye (1) (Figure 4b); reduced to a fovea (2) (Figure 4c). |
| 17 | Mouthparts, inner side on mandibles: with two or more teeth (0) (Figures 1 and 2 [76]); with a single tooth on distal end or lacking tooth (1) (Figures 7 and 8 in [18]) |
| 18 | Mouthparts, scar or lasting appendage on apical surface of mandibles: absent (0); present (1) (Figure 1b and Figure 2e). |
| 19 | Mouthparts, prementum relative to maxillae: prementum incompletely concealing maxillae (phanerognathous condition) (0); prementum completely concealing maxillae (adelognathous condition) (1) (Figure 2f) (Figure 4d,e in [77]). |
| 20 | Mouthparts, long setae on prementum: absent (0) (Figure 2f); present (1) (Figures 9 and 10 in [18]). |
| 21 | Head, transversal furrow or impression between rostrum and forehead (dorsal view): present (0) (Figure 1c); absent (1). |
| 22 | Head, width of forehead relative to minimum width of rostrum (without pterigia): forehead wider than rostrum (0); forehead about same width to narrower than rostrum (1). |
| 23 | Head, orientation of setae behind forehead (vertex): setae anteriorly oriented (0); posteriorly oriented (1). |
| 24 | Head, superciliar arches: absent (0); present (1) (Figure 1e). |
| 25 | Head, circumocular sulcus: absent (0); present (1) (Figure 1e). |
| 26 | Head, large fovea on forehead: present (0); absent or very small (1). |
| 27 | Head, shape and position of eyes: rounded to slightly oval, oriented antero-posteriad (Figure 2b) (0); oval, oriented dorso-ventrad (1) (Figure 1f). |
| 28 | Head, convexity of eyes: flat (slightly protruding) (0) (Figure 1b); convex (protruding) (1) (Figure 1d); conical (strongly protruding) (2) (Figure 3a). |
| 29 | Eyes, position relative to anterior margin of pronotum: slightly separated from pronotum (by 1× or less than 1X the diameter of eyes) (0) (Figure 3c); largely separated from pronotum (by 1× of more than 1× the diameter of eyes) (1) (Figure 3b). |
| 30 | Antennae, shape and width of scape: clavate to spatulate, broad (0) (Figure 2b); clavate to capitate, slender (1) (Figure 3c). |
| 31 | Antennae, curvature of scape: straight (0) (Figure 1a); curved (1) (Figure 3b). |
| 32 | Antennae, scape in resting position: passing below eyes (0); passing across dorsal half of eyes or over eyes (1) (Figure 2c). |
| 33 | Antennae, length of scape: not reaching anterior margin of eyes (0); exceeding anterior margin of eye, but not exceeding anterior margin of pronotum (1); largely exceeding anterior margin of pronotum (2). |
| 34 | Antennae, relative length of funicle antennomeres 1 and 2: funicle antennomere 2 slightly shorter than 1 or both subequal (0); funicle antennomer 2 longer than 1 (1). |
| 35 | Antennae, length/ width ratio of funicle antennomeres 4–7: wider than long (0); about as long as wide (1); distinctly longer than wide (2). |
| 36 | Pronotum, postocular lobes: absent (0); present, not covering eyes (1) (Figure 1e); present, covering eyes (2) (Figure 1c). |
| 37 | Pronotum, long vibrissae on lateral margins: absent (0); present (1) (Figure 2d). |
| 38 | Pronotum, shape: subcylindrical to subconical, not impressed behind anterior margin (0); subhexagonal, impressed behind anterior margin (1) (Figure 3d). |
| 39 | Pronotum, basal margin: not constricted (0); strongly constricted (1) (Figure 3d in [28]). |
| 40 | Prosternum, channel to accommodate the rostrum in repose: absent (0); present (1). |
| 41 | Scutellum, shape: subtriangular (0); quadrate to rounded (1); suboval to subrectangular (longitudinal) (2) (Figure 3d); subcircular (3). |
| 42 | Elytra, striae 9 and 10: closer to each other on posterior 2/3 (0) (Figure 3f); equally separated along their extension (1) (Figure 3e); closer to each other on anterior 1/3 (2). |
| 43 | Elytra, outline of apex: entire (0); bifid (1) (Figure 3a,d in [28]). |
| 44 | Procoxae, position relative to anterior and posterior margins of prosternum: about equally close to both margins of prosternum (0); distinctly closer to anterior margin (1). |
| 45 | Femora, large tooth on ventral edge: absent (0); present on all femora (1) (Figure 3c). |
| 46 | Femora, width of profemora relative to width of metafemora: profemora about as wide as metafemora (0); 1.25–1.5× as wide as metafemora (1); more than 1.5× as wide as metafemora (2); narrower than metafemora (3) (Figures 2a and 3a,d,e in [28]). |
| 47 | Tibiae, mucro on protibiae: reduce to indistinct (0) (Figure 3b); moderate to large, not forming forceps with tuft of stiff setae (1) (Figure 3a); large, forming forceps with tuft of stiff setae (2). |
| 48 | Tibiae, line of denticles along inner edge: absent in all tibiae (0); present at least on protibiae (1) (Figure 3a). |
| 49 | Tibiae, spurs at metatibial apex: present (0); absent (1). |
| 50 | Tibiae, true corbel at metatibial apex: absent (metatibial apex simple or corbel open); present (1) (Figures 5A, 5B–D in [3]). |
| 51 | Tibiae, false corbel (inner flange or corbel semi-enclosed): absent (0); present (1) (Figure 5E,F in [3]). |
| 52 | Tibiae, dorsal comb at metatibial apex (corbel ascending): absent (0); present (1) (Figures A–B and E–J in [78]). |
| 53 | Tarsal claws: free, widely separated (0) (Figure 4d); free, moderately separated (1) (Figure 4e); connate (2) (Figure 4f–g). |
| 54 | Metepisternum and metepisteral-metasternal suture: metepisternum wide, suture complete (0) (Figure 3f); metepisternum narrow, suture complete (1); metepisternum indistinct, suture incomplete, partially fused with metasternum (2) (Figure 3e). |
| 55 | Metasternum, length relative to metacoxae: about 2X as long as metacoxae (0); 1–2× as long as metacoxae (1) (Figure 3f); less than 1× as long as metacoxae (2) (Figure 3e). |
| 56 | Venter, convexity of ventrites: flat to slightly convex (0); ventrites 1 and 2 more convex than remaining ventrites (Figure 3e) (1); ventrites 1–4 very convex (2). |
| 57 | Venter, separation between metacoxae: equal to slightly narrower than width of each metacoxa (0); much wider than width of each metacoxae (1). |
| 58 | Venter, length of ventrite 2 relative to ventrites 3 + 4 (female): longer than 3+ 4 (0); about as long as to slightly shorter than 3 + 4 (1). |
| 59 | Female genitalia, shape of plate of sternite VIII: subtriangular (0); not subtriangular (1). (Figure 6a–g in [28]) |
| 60 | Female genitalia, length of apodeme of sternite VIII: about as long as plate (0); distinctly longer than plate (1). |
| 61 | Female genitalia, distal gonocoxites and styli: gonocoxites membranous to slightly sclerotized, styli present (0); gonocoxites strongly sclerotized, styli absent (1) (Figure 7c,d in [28]) |
| 62 | Female genitalia, pair of baculi along ovipositor: absent (0); present (1) (Figure 7b–e in [28]). |
| 63 | Larva, length and shape of antennal sensorium: longer than wide, subconical (0) (Figure 53 [75]; wider than long, cushion like (1) (Figure 2 in [74]). |
| 64 | Larva, shape of antennal sensorium in apical view: subcircular (0) (Figure 10 in [4]); elliptical (1) (Figure 13 in [4]). |
| 65 | Larva, number of ventral setae on maxillary mala: five (0) (Figure 56 in [75]); four (1) (Figures 7, 52, and 74 in [74]). |
| 66 | Larva, shape of posterior extension of premental sclerite of labium: acute at apex (0) (Figure 28 in [4]); truncate at apex (1) (Figures 33 and 34 in [4]). |
| 67 | Larva, accessory teeth on intermediate portion of mandibular cutting-edge: present (0) (Figure 20 in [4]); absent (1) (Figure 6 in [79]). |
| 68 | Larva, mandibular scrobe: sclerotized, similar to the rest of mandible (0) (Figure 55 in [75]); slightly sclerotized, paler than the rest of mandible (1) (Figure 6 in [79]). |
| 69 | Larva, labral rods: subparallel (0)(Figures 3–5 in [79]); divergent in their distal half (1) (Figures 1 and 2 in [79]); U-shaped (2) (Figures 50 and 60 in [74]). |
| 70 | Larva, number of setae on alar area: two (0) (Figure 9 in [74]); one (1) (Figure 7 in [79]). |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marvaldi, A.E.; Del Río, M.G.; Pereyra, V.A.; Rocamundi, N.; Lanteri, A.A. A Combined Molecular and Morphological Approach to Explore the Higher Phylogeny of Entimine Weevils (Coleoptera: Curculionidae), with Special Reference to South American Taxa. Diversity 2018, 10, 95. https://doi.org/10.3390/d10030095
Marvaldi AE, Del Río MG, Pereyra VA, Rocamundi N, Lanteri AA. A Combined Molecular and Morphological Approach to Explore the Higher Phylogeny of Entimine Weevils (Coleoptera: Curculionidae), with Special Reference to South American Taxa. Diversity. 2018; 10(3):95. https://doi.org/10.3390/d10030095
Chicago/Turabian StyleMarvaldi, Adriana E., María Guadalupe Del Río, Vanina A. Pereyra, Nicolás Rocamundi, and Analía A. Lanteri. 2018. "A Combined Molecular and Morphological Approach to Explore the Higher Phylogeny of Entimine Weevils (Coleoptera: Curculionidae), with Special Reference to South American Taxa" Diversity 10, no. 3: 95. https://doi.org/10.3390/d10030095
APA StyleMarvaldi, A. E., Del Río, M. G., Pereyra, V. A., Rocamundi, N., & Lanteri, A. A. (2018). A Combined Molecular and Morphological Approach to Explore the Higher Phylogeny of Entimine Weevils (Coleoptera: Curculionidae), with Special Reference to South American Taxa. Diversity, 10(3), 95. https://doi.org/10.3390/d10030095