Reconnecting Amphibian Habitat through Small Pond Construction and Enhancement, South Okanagan River Valley, British Columbia, Canada


Abstract
:1. Introduction
2. Methods
2.1. Site Selection
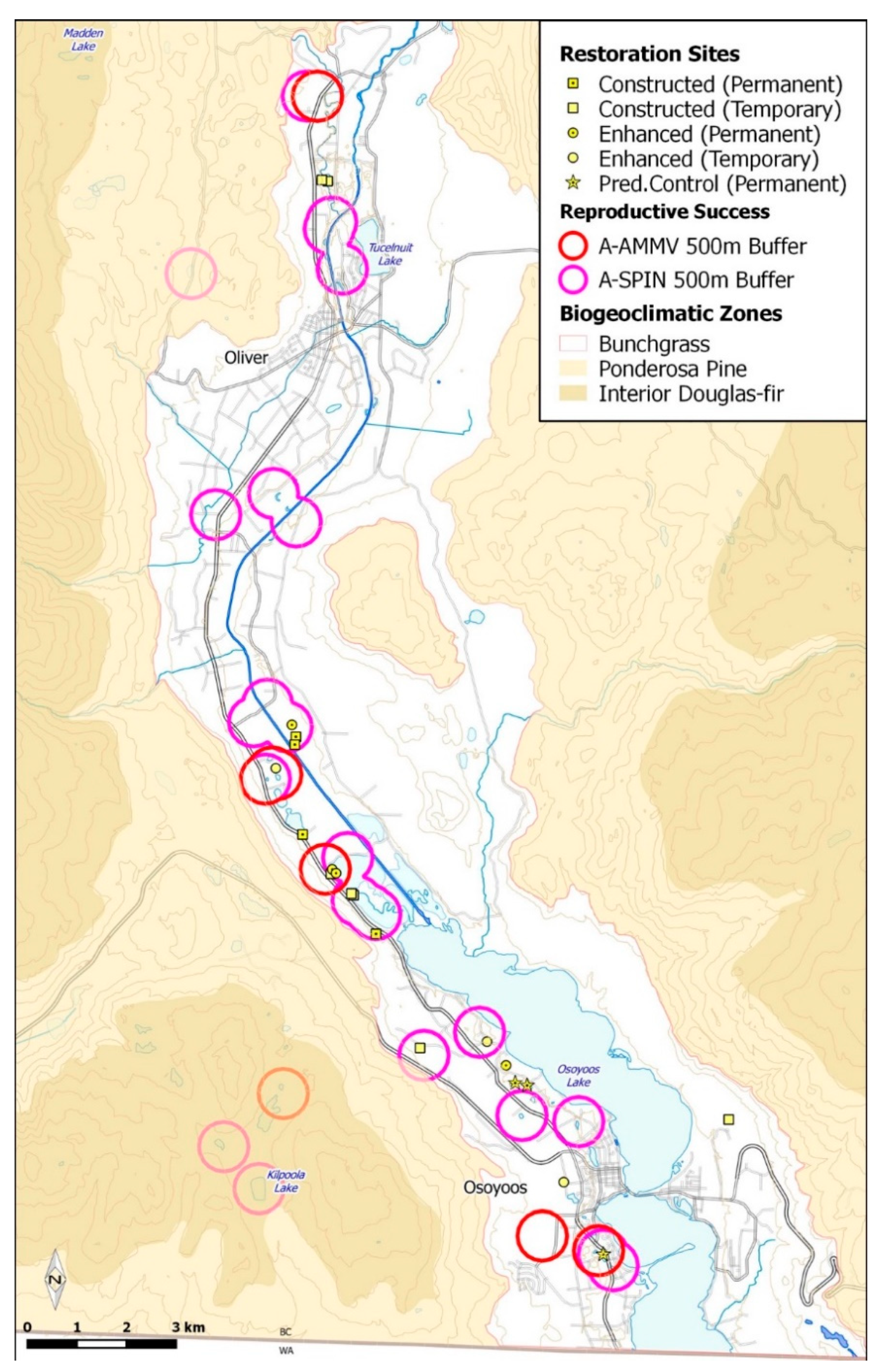
2.2. Pond Habitat Construction and Enhancement
- Construction (N = 10): No pre-existing pond in the locale since at least 1938 [12],
- Enhancement (N = 8): Pond existed historically and was in filled,
- Invasive predatory species removal (N = 3): Pond exists with predatory non-native species present.
2.3. Species Monitoring
2.4. Terrestrial Soil Characteristics
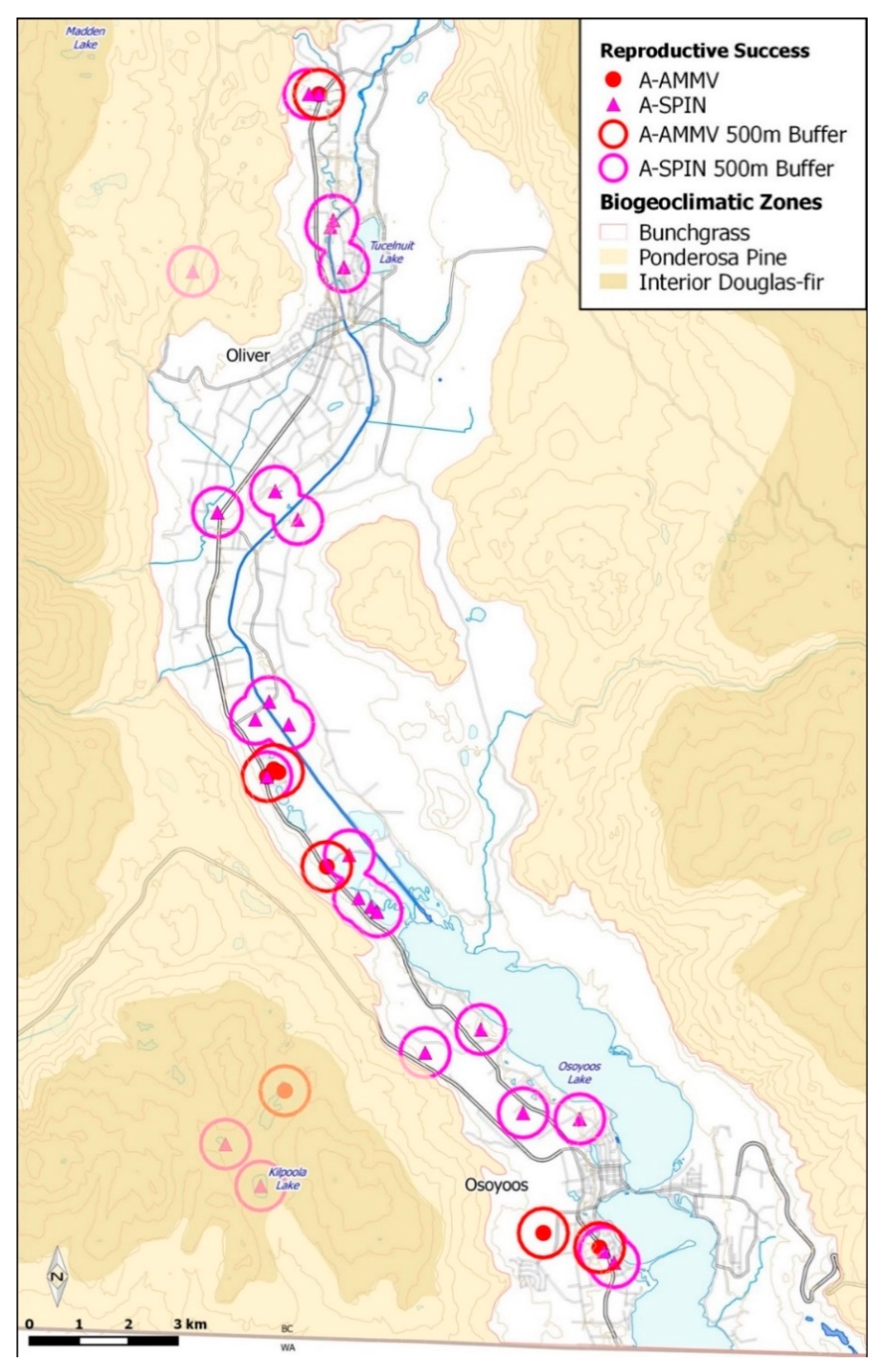
3. Results
3.1. Species Monitoring
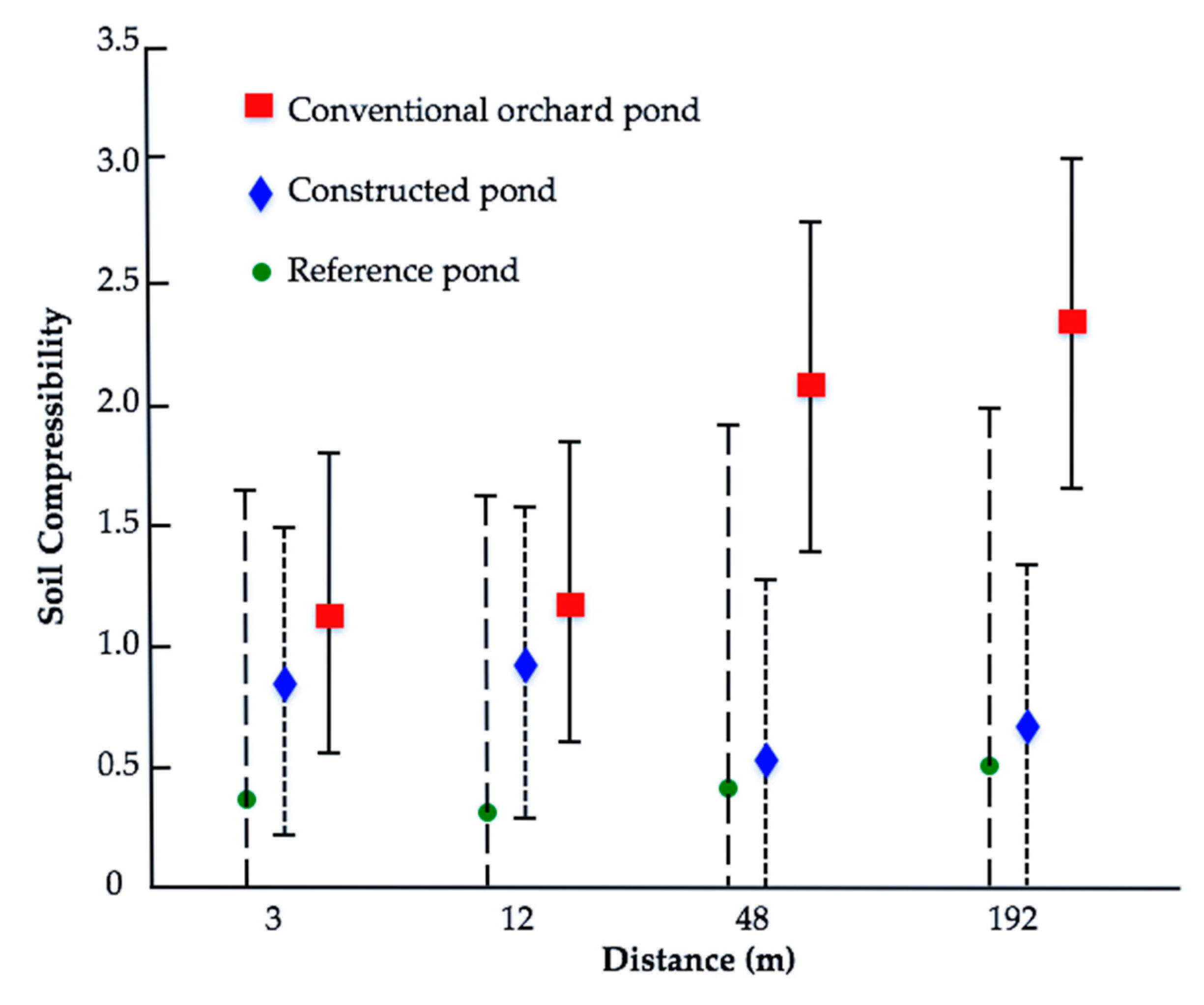
3.2. Substrate Characteristics
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stuart, S.N.; Chanson, J.S.; Cox, N.A.; Young, B.E.; Rodrigues, A.S.L.; Fishman, D.L.; Waller, R.W. Status and trends of amphibian and extinctions worldwide. Science 2004, 306, 1783–1786. [Google Scholar] [CrossRef] [PubMed]
- Wake, D.B.; Vredenburg, V.T. Are we in the midst of the sixth mass extinction? A view from the world of amphibians. Proc. Natl. Acad. Sci. USA 2008, 105, 11466–11473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaustein, A.R.; Han, B.A.; Relyea, R.A.; Johnson, P.T.; Buck, J.C.; Gervasi, S.S.; Kats, L.B. The complexity of amphibian population declines: Understanding the role of cofactors in driving amphibian losses. Ann. N. Y. Acad. Sci. 2011, 1223, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Calhoun, A.J.K.; Arrigoni, J.; Brooks, R.P.; Hunter, M.L.; Richter, S.C. Creating successful vernal pools: A literature review and advice for practitioners. Wetlands 2014, 34, 1027–1038. [Google Scholar] [CrossRef]
- Pechmann, J.H.K.; Estes, R.A.; Scott, D.E.; Gibbons, J.W. Amphibian colonization and use of ponds created for trial mitigation of wetland loss. Wetlands 2000, 21, 93–111. [Google Scholar] [CrossRef]
- Shulse, C.D.; Semlitsch, R.D.; Trauth, K.M.; Williams, A.D. Influences of design and landscape placement parameters on amphibian abundance in constructed wetlands. Wetlands 2010, 30, 915–928. [Google Scholar] [CrossRef]
- Lesbarrères, D.; Ashpole, S.L.; Bishop, C.A.; Blouin-Demers, G.; Brooks, R.J.; Echaubard, P.; Govindarajulu, P.; Green, D.M.; Hecnar, S.J.; Herman, T.; et al. Conservation of herpetofauna in northern landscapes: Threats and challenges from a Canadian perspective. Biol. Conserv. 2014, 170, 48–55. [Google Scholar] [CrossRef]
- British Columbia. Statistics. Available online: http://www.bcstats.gov.bc.ca/StatisticsBySubject/EnvironmentalStatistics.aspx. (accessed on 4 September 2016).
- Ashpole, S.L. Restoring a wetland complex for amphibian populations, South Okanagan Valley, British Columbia, Canada (2003 to 2014). Ph.D. Thesis, University of Waterloo, Waterloo, ON, Canada, 2016. [Google Scholar]
- Committee on the Status of Endangered Wildlife in Canada. COSEWIC Assessment and Update Status Report on the Great Basin Spadefoot Spea Intermontana in Canada; COSEWIC: Ottawa, ON, Canada, 2007; Available online: www.sararegistry.gc.ca/status/status_e.cfm (accessed on 4 September 2015).
- Committee on the Status of Endangered Wildlife in Canada. COSEWIC Assessment and Status Report on the Tiger Salamander Ambystoma Tigrinum in Canada; COSEWIC: Hull, QC, Canada, 2011. [Google Scholar]
- Lea, T. Historical (pre-settlement) ecosystems of the Okanagan Valley and Lower Similkameen Valley of British Columbia—Pre European contact to the present. Davidsonia 2008, 19, 3–33. [Google Scholar]
- South Okanagan Similkameen Conservation Program. Keeping Nature in Our Future: A Biodiversity Conservation Strategy for the South Okanagan Similkameen. Available online: http://www.rdosmaps.bc.ca/min_bylaws/contract_reports/CorpBd/2012/11June07/PlanningDev/2_BiodiversityStrategyExecutiveSummary2012.pdf (accessed on 25 May 2012).
- Knutson, M.G.; Richardson, W.B.; Reineke, D.M.; Gray, B.R.; Parmelee, J.R.; Wick, S.E. Agricultural ponds support amphibian populations. Ecol. Appl. 2004, 14, 669–684. [Google Scholar] [CrossRef]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Diamond, J.M. The Island Dilemma: Lessons of modern biogeographic studies for the design of natural reserves. Biol. Conserv. 1975, 7, 129–146. [Google Scholar] [CrossRef]
- Levins, R. Some demographic and genetic consequences of environmental heterogeneity for biological control. Bull. Entomol. Soc. Am. 1969, 15, 237–240. [Google Scholar] [CrossRef]
- Sutter, G.; Francisco, R. Vernal pool creation in the Sacramento Valley: A review of issues surrounding its role as a conservation tool. In Ecology, Conservation, and Management of Vernal Pool Ecosystems, Proceedings from 1996 Conference; Witham, C.W., Bauder, E.T., Belk, D., Ferren, W.R., Jr., Ornduff, R., Eds.; California Native Plant Society: Sacramento, CA, USA, 1998; pp. 190–194. [Google Scholar]
- Scheffers, B.R.; Paszkowski, C.A. Amphibians use of urban storm wetlands: The role of natural habitat features. Landsc. Urban Plan. 2013, 113, 139–149. [Google Scholar] [CrossRef]
- Brown, D.J.; Street, G.M.; Nairn, R.W.; Forstner, M.R. A place to call home: Amphibian use of created and restored wetlands. Int. J. Ecol. 2012, 2012, 989872. [Google Scholar] [CrossRef]
- Porej, D.; Hetherington, T.E. Designing wetlands for amphibians: The importance of predatory fish and shallow littoral zones in structuring of amphibian communities. Wetl. Ecol. Manag. 2005, 13, 445–455. [Google Scholar] [CrossRef]
- Lehtinen, R.M.; Galatowitsch, S.M. Colonization of restored wetlands by amphibians in Minnesota. Am. Midl. Nat. 2001, 145, 388–396. [Google Scholar] [CrossRef]
- Petranka, J.W.; Holbrook, C.T. Wetland restoration for amphibians: Should local sites be designed to support metapopulations or patchy populations? Restor. Ecol. 2006, 14, 404–411. [Google Scholar] [CrossRef]
- Semlitsch, R.D. Principles for management of aquatic-breeding amphibians. J. Wildl. Manag. 2000, 64, 615–631. [Google Scholar] [CrossRef]
- Semlitsch, R.D.; Brodie, J.R. Biological criteria for buffer zones around wetlands and riparian habitats for amphibians and reptiles. Conserv. Biol. 2003, 17, 1219–1228. [Google Scholar] [CrossRef]
- Environment and Climate Change Canada. Recovery Strategy for the Great Basin Spadefoot (Spea intermontana) in Canada [Proposed]; Species at Risk Act Recovery Strategy Series; Environment and Climate Change Canada: Ottawa, ON, Canada, 2017. [Google Scholar]
- Environment and Climate Change Canada. Recovery Strategy for the Tiger Salamander (Ambystoma tigrinum) Southern Mountain Population in Canada; Species at Risk Act Recovery Strategy Series; Environment and Climate Change Canada: Ottawa, ON, Canada, 2017. [Google Scholar]
- Crosby, J. Amphibian Occurrence on South Okanagan Roadways (2010–2012): Investigating Movement Patterns, Crossing Hotspots, & Roadkill Mitigation Structure Use at the Landscape Scale. Master’s Thesis, University of Waterloo, Waterloo, ON, Canada, 2014. [Google Scholar]
- Biebighauser, T.R. Guide to Creating Vernal Ponds; USDA Forest Service, United States Department of Agriculture: Washington, DC, USA, 2003. [Google Scholar]
- Moncada, K.M.; Sheaffer, C.C. Chapter 13: Tools for Managing Pest and Environmental Risks to Organic Crops in The Upper Midwest. In Organic Risk Management Handbook; University of Minnesota: Minneapolis, MN, USA, 2010; Available online: http://www.organicriskmanagement.umn.edu/default.htm (accessed on 31 December 2014).
- Ashpole, S.L.; Bishop, C.A.; Elliott, J.E. Unexplained die-off of larval tiger salamanders (Ambystoma tigrinum) in an agricultural pond in the South Okanagan Valley, British Columbia, Canada. Northwest. Nat. 2011, 92, 221–224. [Google Scholar] [CrossRef]
- Lukey, N.A.; Cunnington, D.; Ashpole, S.L.; Murphy, S.D.; Govindarajulu, P.P. Where Do We Go from Here? Long-Term, Adaptive Management of Invasive American Bullfrogs (Lithobates catesbeianus) in the South Okanagan Valley, British Columbia, August 2012; World Congress of Herpetology: Vancouver, BC, Canada, 2012. [Google Scholar]
- British Columbia Ministry of Environment. Guidelines for Amphibian and Reptile Conservation during Urban and Rural Land Development in British Columbia; B.C. Ministry of Environment: Victoria, BC, Canada, 2014.
- Olfert, E.D.; Cross, B.M.; McWilliam, A.A. (Eds.) Guide to the Care and Use of Experimental Animals; Canadian Council on Animal Care: Ottawa, ON, Canada, 1993; Volume 1, No. 2. [Google Scholar]
- StatSoft, Inc. Statistica for Windows [Computer Program Manual]; TIBCO Software Inc.: Tulsa, OK, USA, 2003. [Google Scholar]
- Shulse, C.D.; Semlitsch, R.D.; Trauth, K.M.; Gardner, J.E. Testing wetland features to increase amphibian reproductive success and species richness for mitigation and restoration. Ecol. Appl. 2012, 22, 1675–1688. [Google Scholar] [CrossRef] [PubMed]
- Bellakhal, M.; Neveu, A.; Aleya, L. Artificial wetlands as a solution to the decline in the frog population: Estimation of their suitability through the study of population dynamics of Sahara Frogs in hill lakes. Ecol. Eng. 2014, 63, 114–121. [Google Scholar] [CrossRef]
- Morey, S.R. Pool duration influence age and body mass at metamorphosis in the Western Spadefoot toad: Implications for vernal pool conservation. In Ecology, Conservation, and Management of Vernal Pool Ecosystems, Proceedings from 1996 Conference; Witham, C.W., Bauder, E.T., Belk, D., Ferren, W.R., Jr., Ornduff, R., Eds.; California Native Plant Society: Sacramento, CA, USA, 1998. [Google Scholar]
- Richardson, J.S.; Oaten, D. Critical Breeding, Foraging, and Overwintering Habitats of Great Basin Spadefoot Toads (Spea intermontana) and Western Toads (Anaxyrus boreas) within Grassland Ecosystems; Final Report; Canadian Wildlife Federation Kanata: Ottawa, ON, Canada, 2013. [Google Scholar]
- Ashpole, S.L.; Bishop, C.A.; Elliott, J.E. Use of anthropogenic breeding sites and clutch size in the Great Basin Spadefoot (Spea intermontana), South Okanagan Valley, British Columbia. Canada. Northwest. Nat. 2014, 95, 35–40. [Google Scholar] [CrossRef]
- Richardson, J.S.; Klenner, W.; Shatford, J. The tiger salamander in British Columbia: An amphibian in an endangered desert environment. In Proceedings of the Conference on the Biology and Management of Species and Habitats at Risk, Kamloops, BC, Canada, 15–19 February 1999; Darling, L.M., Ed.; University College of the Cariboo: Kamloops, BC, Canada, 2000; pp. 407–412. [Google Scholar]
- Porej, D.; Micacchion, M.; Hetherington, T.E. Core terrestrial habitat for conservation of local populations of salamanders and wood frogs in agricultural landscapes. Biol. Conserv. 2004, 120, 399–409. [Google Scholar] [CrossRef]
- Shoo, L.P.; Olson, D.H.; McMenamin, S.K.; Murray, K.A.; Van Sluys, M.; Donnelly, M.A.; Stratford, D.; Terhivuo, J.; Merino-Viteri, A.; Herbert, S.M.; et al. Engineering a future for amphibians under climate change. J. Appl. Ecol. 2011, 48, 487–492. [Google Scholar] [CrossRef] [Green Version]
- Ngo, A.; MCKay, V.L.; Murphy, R.W. Recovery Strategy for Tiger Salamander (Ambystoma tigrinum) (Great Lakes Population) in Canada [Proposed]; Species at Risk Act Recovery Strategy Series; Parks Canada Agency: Ottawa, ON, Canada, 2009. [Google Scholar]
- Trenham, P.C.; Koenig, W.D.; Shaffer, H.B. Spatially autocorrelated demography and interpond dispersal in the salamander Ambystoma californiense. Ecology 2001, 82, 3519–3530. [Google Scholar] [CrossRef]
- Greenberg, C.H.; Tanner, G.W. Breeding pond selection and movement patterns by eastern spadefoot toads (Scaphiopus holbrookii) in relation to weather and edaphic conditions. J. Herpetol. 2004, 38, 569–577. [Google Scholar] [CrossRef]
- Nyström, P.; Hansson, J.; Månsson, J.; Sundstedt, M.; Reslow, C.; Broström, A. A documented amphibian decline over 40 years: Possible causes and implications for species recovery. Biol. Conserv. 2007, 138, 399–411. [Google Scholar] [CrossRef]
- Baughman, B.; Todd, B.D. Role of substrate cues in habitat selection by recently metamorphosed Bufo terrestris and Scaphiopus holbrookii. J. Herpetol. 2007, 41, 154–157. [Google Scholar] [CrossRef]
- Iverson, K.; Curran, D.; Fleming, T.; Haney, A. Sensitive Ecosystems Inventory—Okanagan Valley: Vernon to Osoyoos, 2000–2007. Methods, Ecological Descriptions, Results and Conservation Tools; Technical Report Series No. 495; Canadian Wildlife Service, Pacific and Yukon Region: Delta, BC, Canada, 2008. [Google Scholar]
- Gray, M.J.; Smith, L.M.; Brenes, R. Effects of agriculture cultivation on demographics of Southern high plains amphibians. Conserv. Biol. 2004, 18, 1368–1377. [Google Scholar] [CrossRef]
- Carisio, L.; Sacchi, R.; Seglie, D.; Sindaco, R. Habitat selection in the fossorial toad Pelobates fuscus insbricus (Amphibia: Pelobatidae). Acta Herpetol. 2014, 9, 51–59. [Google Scholar]
- Hobbs, R.J. Setting effective and realistic restoration goals: Key direction for research. Restor. Ecol. 2007, 15, 354–357. [Google Scholar] [CrossRef]
- Janin, A.; Lena, J.-P.; Deblois, S.; Joly, P. Use of stress-hormone levels and habitat selection to assess functional connectivity of a landscape for an amphibian. Conserv. Biol. 2012, 26, 923–931. [Google Scholar] [CrossRef] [PubMed]
- Jansen, K.P.; Summers, A.P.; Delis, P.R. Spadefoot toads (Scaphiopus holbrookii holbrookii) in an urban landscape: Effects of nonnatural substrates on burrowing in adults and juveniles. J. Herpetol. 2001, 35, 141–145. [Google Scholar] [CrossRef]
- Garner, J.L. Movement and Habitat-Use of the Great Basin Spadefoot (Spea intermontana) at Its Northern Range Limit. Master’s Thesis, Thompson Rivers University, Kamloops, BC, Canada, 2012; pp. 1–8. [Google Scholar]
- Newman, R.A.; Dunham, A.E. Size at metamorphosis and water loss in a desert anuran (Scaphiopus couchii). Copeia 1994, 1994, 372–381. [Google Scholar] [CrossRef]
- Roe, J.H.; Georges, A. Heterogeneous wetlands complexes, buffer zones, and travel corridors: Landscape management for freshwater reptiles. Biol. Conserv. 2007, 135, 67–76. [Google Scholar] [CrossRef]
- Lindenmayer, D.; Hobbs, R.J.; Montague-Drake, R.; Alexandra, J.; Bennett, A.; Burgman, M.; Cale, P.; Calhoun, A.; Cramer, V.; Cullen, P.; et al. A checklist for ecological management of landscapes for conservation. Ecol. Lett. 2008, 11, 78–91. [Google Scholar] [CrossRef] [PubMed]
- Ghermandi, A.; Van Den Bergh, J.C.; Brander, L.M.; de Groot, H.L.; Nunes, P.A. Values of natural and human-made wetlands: A meta-analysis. Water Resour. Res. 2010, 46, W12516. [Google Scholar] [CrossRef]
- Parrott, L.; Kyle, C. The Value of Natural Capital in the Okanagan. Okanagan Institute for Biodiversity, Resilience, and Ecosystem Services (BRAES); The University of British Columbia: Kelowna, BC, Canada, 2014. [Google Scholar]
- Allan, D.J.; McIntyre, P.B.; Smith, S.D.P.; Halpern, B.S.; Boyer, G.L.; Buchsbaum, A.; Burton, G.A., Jr.; Campbell, L.M.; Chadderton, W.L.; Ciborowski, J.J.H.; et al. Joint analysis of stressors and ecosystem services to enhance restoration effectiveness. Proc. Natl. Acad. Sci. USA 2013, 110, 327–377. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Land Tenure | Site # | Year * | Activity Type | Pond Dimensions (m) | Pond Shape | Pond Water Sub-Class | Type of Closest Water Body | Distance to Nearest Water Bodies (m) | Distance to Closest Known Breeding Pond ^ (m) | Distance to Highway (m) |
|---|---|---|---|---|---|---|---|---|---|---|
| Private | 1 | 2006 | Enhancement | 22 × 28 | Half moon | Permanent | Creek | 118 | 668 | 243 |
| NGO | 2 | 2006 + | Enhancement | 20 × 30 | Oval | Permanent | Isolated oxbow | 35 | 498 | 973 |
| NGO | 3 | 2006 + | Constructed | 27 × 32 | Oval | Permanent | Isolated oxbow | 110 | 287 | 828 |
| NGO | 4 | 2006 | Constructed | 6 × 11 | Oval | Ephemeral | Isolated oxbow | 332 | 277 | 185 |
| NGO | 5 | 2006 + | Constructed | 7 × 14 | Oval | Ephemeral | Isolated oxbow | 365 | 116 | 118 |
| Private | 6 | 2006 | Enhancement | 14 × 37 | Half moon | Ephemeral | Permanent pond | 128 | 629 | 344 |
| Private | 7 | 2007 | Constructed | 6 × 8 | Oval | Ephemeral | Oxbow | 15 | 385 | 310 |
| NGO | 8 | 2007 | Constructed | 8 × 9 | Oval | Permanent | Oxbow | 35 | 210 | 905 |
| NGO | 9 | 2007 + | Enhancement | 17 × 31 | Oval | Permanent | Permanent pond | 100 | 55 | 110 |
| NGO | 10 | 2007 + | Enhancement | 25 × 32 | Oval | Permanent | Permanent pond | 155 | 75 | 110 |
| Private | 11 | 2007 | Enhancement; 2010 liner | 7 × 55; liner 7 × 21 | Teardrop | Ephemeral | Lake | 110 | 1300 | 125 |
| NGO | 12 | 2007 + | Constructed-liner | 30 × 50 | Square | Ephemeral | Ornamental pond | 65 | 65 | 185 |
| NGO | 13 | 2007 + | Constructed-liner | 4 × 8 | Oval | Ephemeral | Ornamental pond | 405 | 405 | 676 |
| Private | 14 | 2008 * | Non-native Fish species | 40 × 100 | Oval | Permanent | Permanent pond | 210 | 377 | 60 |
| 2009 * | ||||||||||
| Private | 15 | 2009 * | Enhancement | 17 × 19 | Oval | Ephemeral | Ephemeral pond | 297 | 1000 | 335 |
| Private | 16 | 2009 * | Enhancement | 50 × 70 | Horseshoe | Permanent | Lake | 239 | 664 | 260 |
| GO | 17 | 2010 | Constructed | 17 × 42 | Teardrop | Permanent | Permanent pond | 79 | 140 | 82 |
| GO | 18 | 2011 | Constructed | 40 × 40 | Oval | Permanent | Ephemeral pond | 240 | 240 | 71 |
| GO | 19 | 2010 | Constructed | 32 × 51 | Oval | Ephemeral | Permanent pond | 75 | 840 | 25 |
| Private | 20 | 2004 | Non-native species | 62 × 84 | Oval | Permanent | Permanent pond | 317 | 360 | 170 |
| 2011 * | ||||||||||
| Private | 21 | 2004 | Non-native species | 76 × 130 | Oval | Permanent | Permanent pond | 317 | 560 | 402 |
| 2011 * |
| Year | 2007 | 2008 | 2009 | 2010 | 2011 | 2012 | 2013 | 2014 | |
|---|---|---|---|---|---|---|---|---|---|
| Number constructed or enhanced ponds | 6 | 14 | 16 | 21 | 21 | 21 | 21 | 21 | |
| Auditory search effort (h) | 2.25 | 4.18 | 4.35 | 5.13 | 7.43 | 5.05 | 6.11 | 7.32 | |
| Active search effort (h) | 4.53 | 12.33 | 10.51 | 10.13 | 9.24 | 9.58 | 8.59 | 13.44 | |
| Great Basin spadefoot | colonized | 2 | 10 | 12 | 15 | 10 | 13 | 15 | 13 |
| metamorph | 1 | 10 | 8 | 5 | 3 | 5 | 5 | 7 | |
| Pacific treefrog | colonized | 7 | 8 | 9 | 9 | 4 | 6 | 8 | |
| metamorph | 1 | 0 | 4 | 3 | 2 | 2 | 2 | 3 | |
| Number sites colonized | 3 | 11 | 14 | 16 | 12 | 14 | 16 | 14 | |
| Number sites with metamorphic success | 1 | 10 | 11 | 7 | 4 | 7 | 6 | 8 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ashpole, S.L.; Bishop, C.A.; Murphy, S.D. Reconnecting Amphibian Habitat through Small Pond Construction and Enhancement, South Okanagan River Valley, British Columbia, Canada. Diversity 2018, 10, 108. https://doi.org/10.3390/d10040108
Ashpole SL, Bishop CA, Murphy SD. Reconnecting Amphibian Habitat through Small Pond Construction and Enhancement, South Okanagan River Valley, British Columbia, Canada. Diversity. 2018; 10(4):108. https://doi.org/10.3390/d10040108
Chicago/Turabian StyleAshpole, Sara L., Christine A. Bishop, and Stephen D. Murphy. 2018. "Reconnecting Amphibian Habitat through Small Pond Construction and Enhancement, South Okanagan River Valley, British Columbia, Canada" Diversity 10, no. 4: 108. https://doi.org/10.3390/d10040108
APA StyleAshpole, S. L., Bishop, C. A., & Murphy, S. D. (2018). Reconnecting Amphibian Habitat through Small Pond Construction and Enhancement, South Okanagan River Valley, British Columbia, Canada. Diversity, 10(4), 108. https://doi.org/10.3390/d10040108