Chromosome Level Genome Assembly and Comparative Genomics between Three Falcon Species Reveals an Unusual Pattern of Genome Organisation

 ,
, 
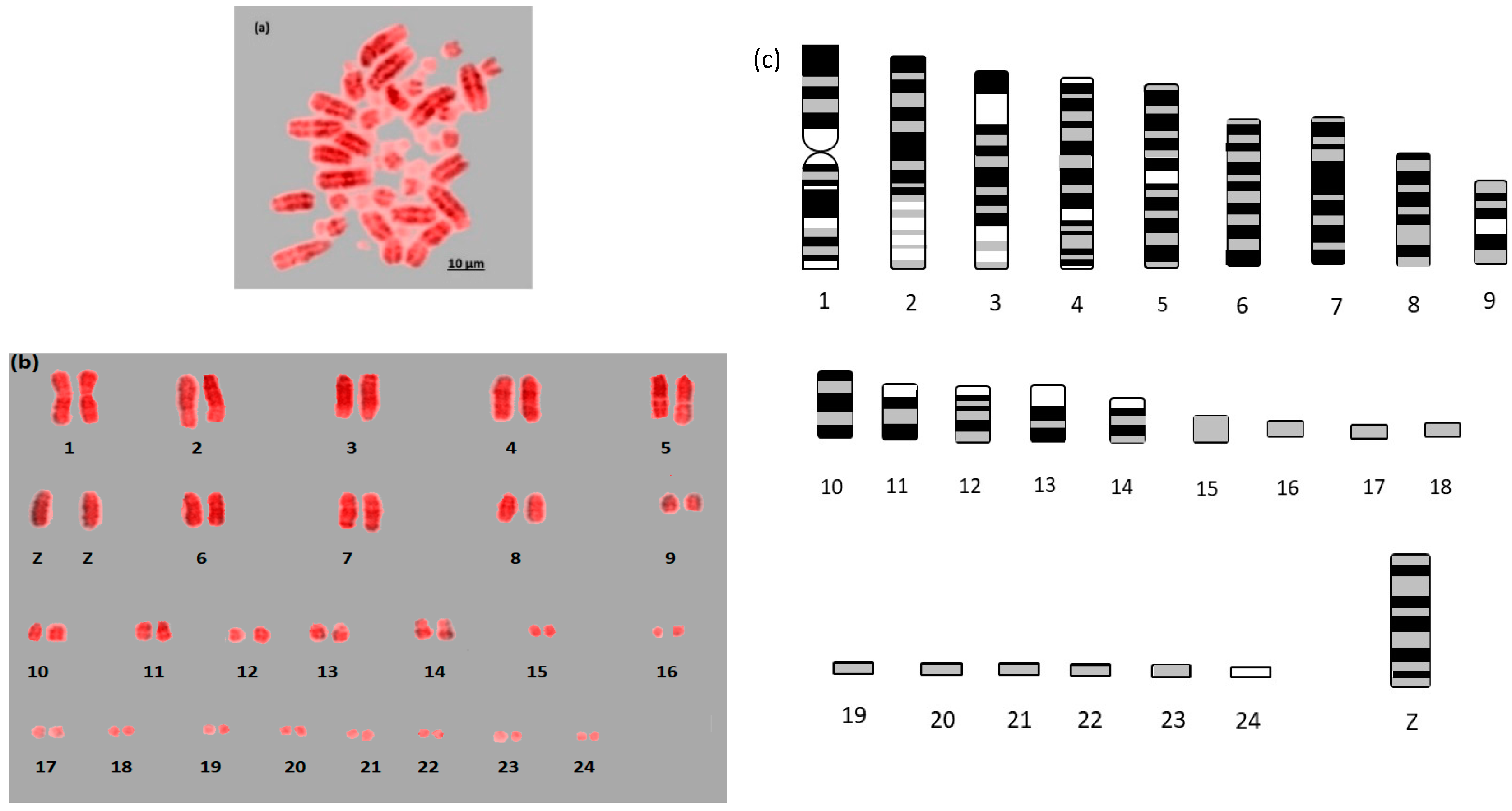{kind=link}
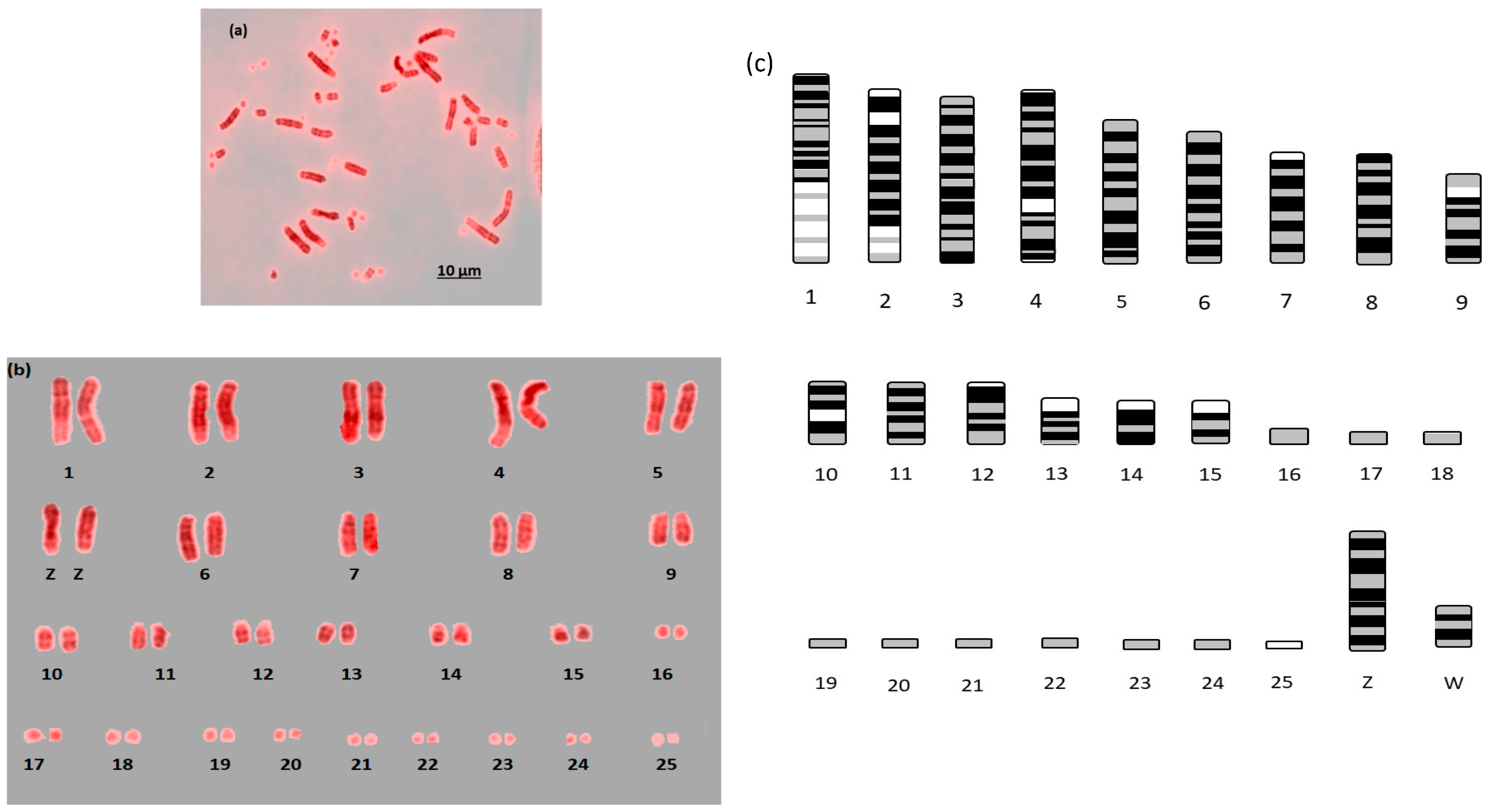{kind=link}
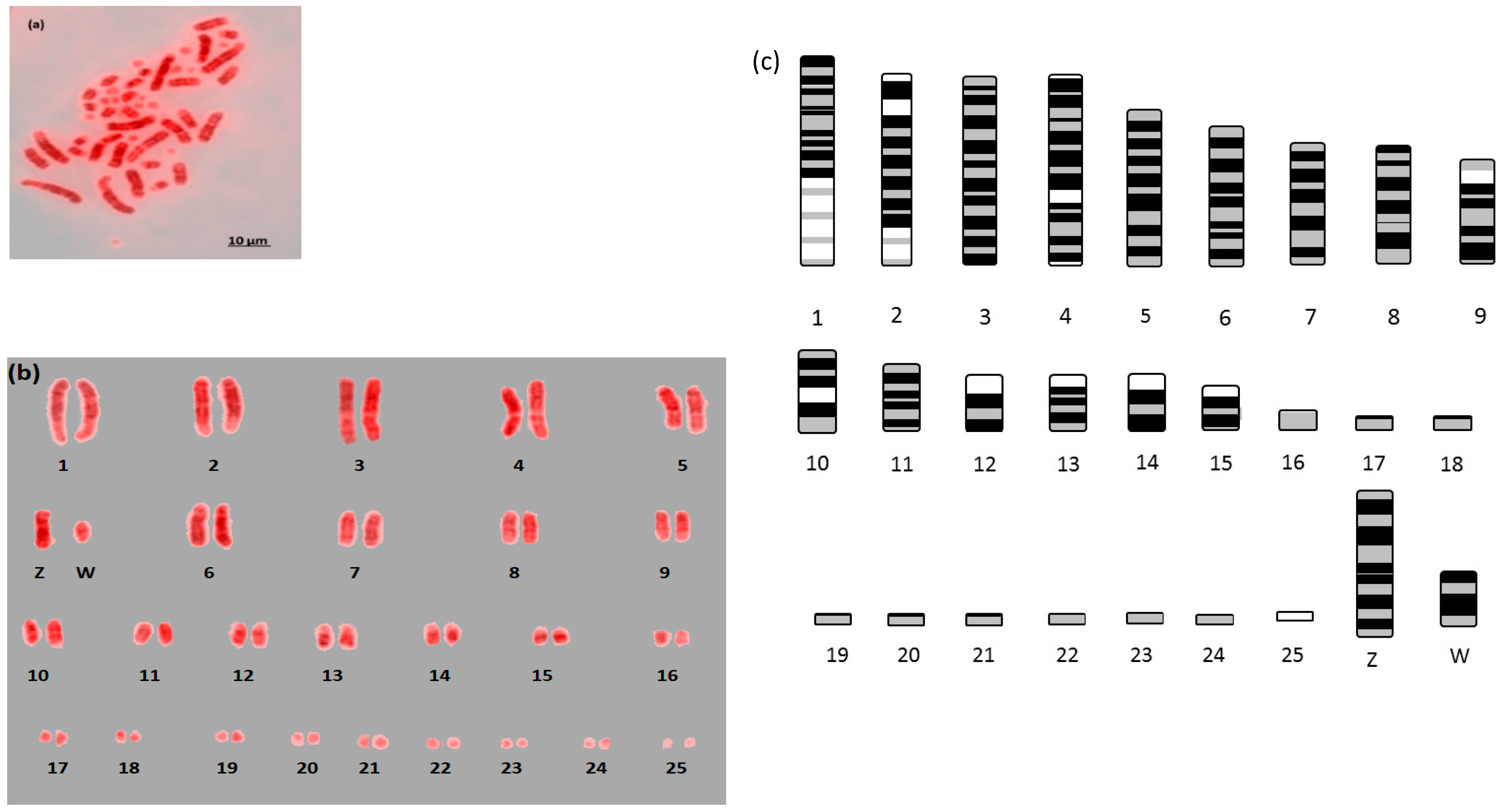{kind=link}
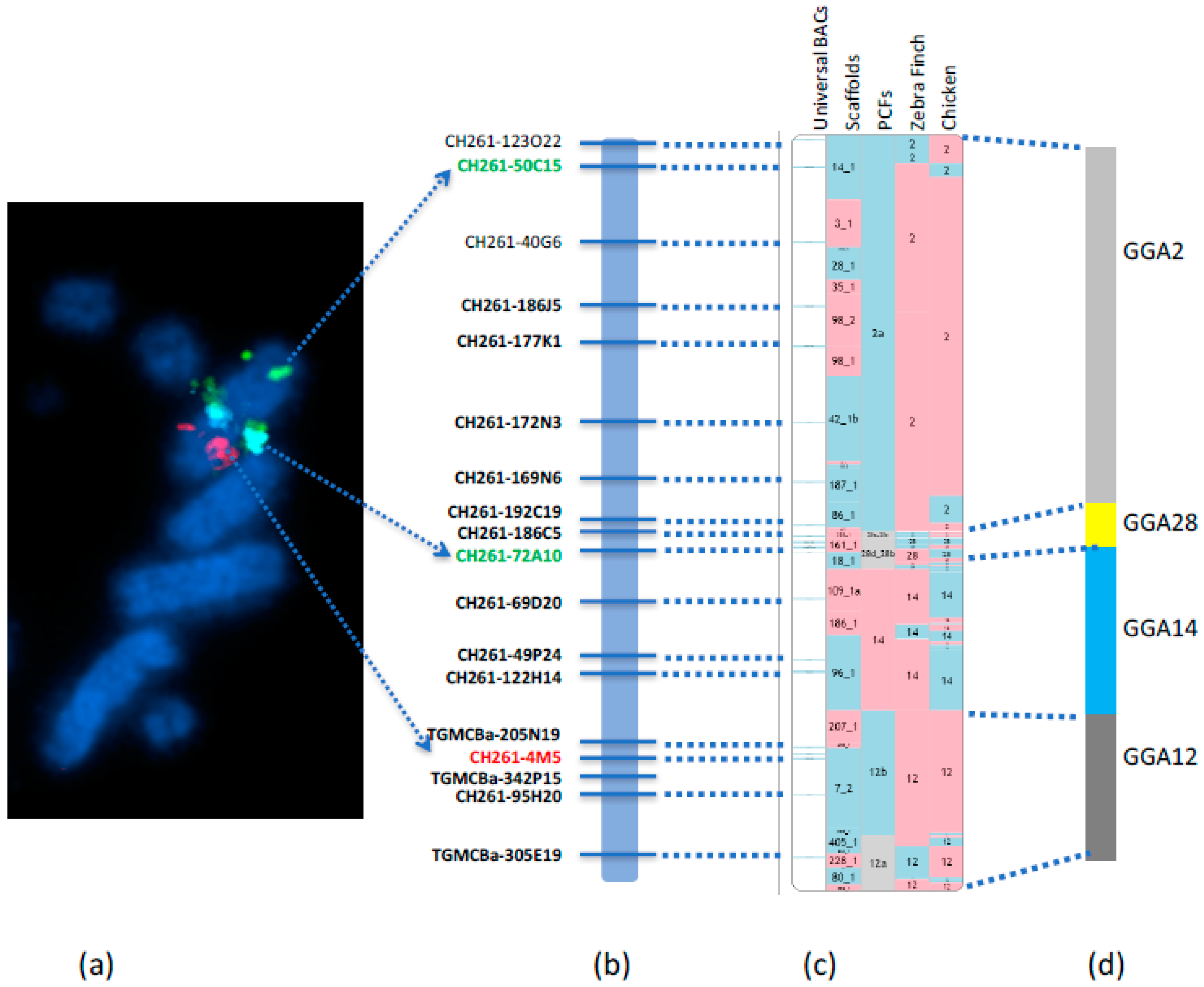{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Chromosome Preparation
2.2. Fluorescence In-Situ Hybridisation (FISH)
2.3. Genome Mapping
2.4. Gyr Falcon Genome Sequencing
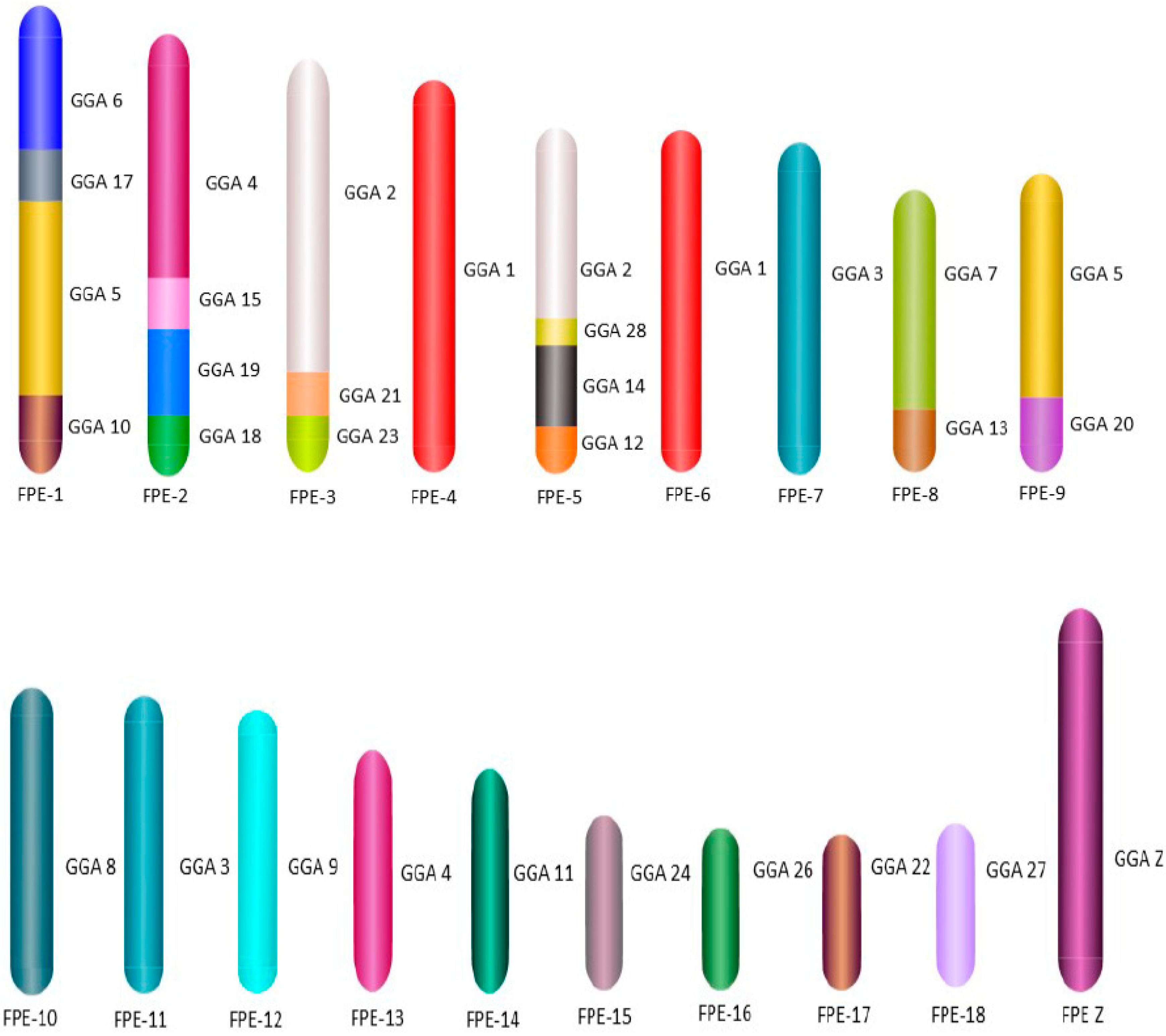
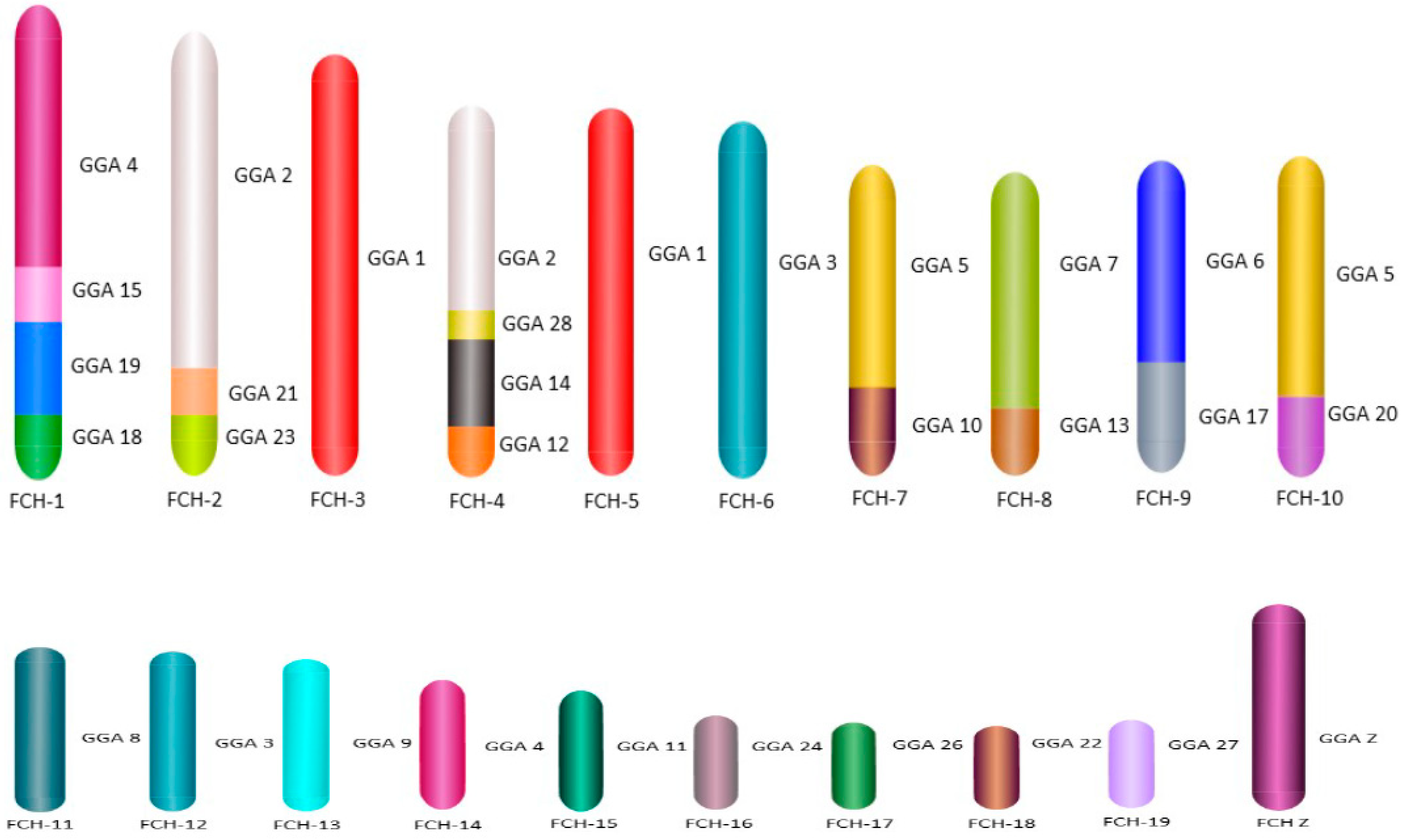
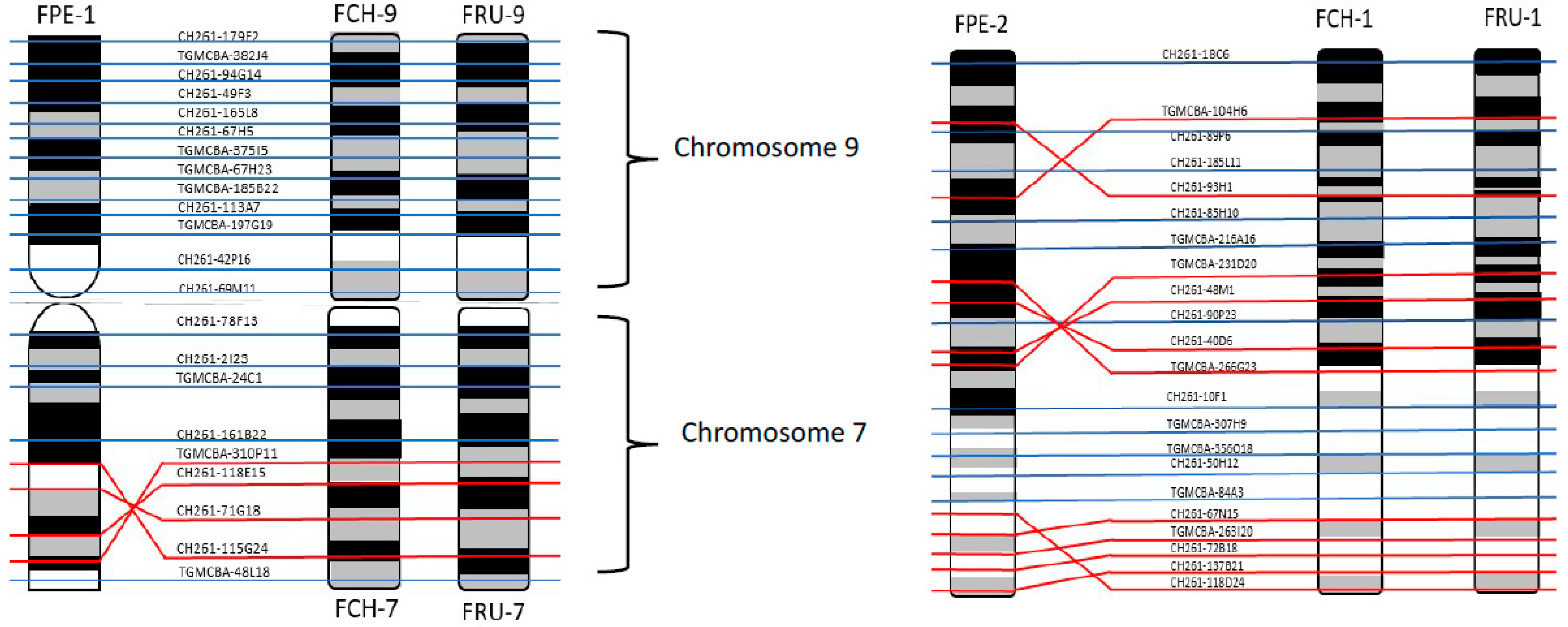
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kim, J.; Larkin, D.M.; Cai, Q.; Zhang, Y.; Ge, R.L.; Ma, J. Reference-assisted chromosome assembly. Proc. Natl. Acad. Sci. USA 2013, 110, 1785–1790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- França, L.T.C.; Carrilho, E.; Kist, T.B.L. A review of DNA sequencing techniques. Q. Rev. Biophys. 2002, 35, 169–200. [Google Scholar] [CrossRef] [PubMed]
- Masabanda, J.S.; Burt, D.W.; O’Brien, P.C.M.; Vignal, A.; Fillon, V.; Walsh, P.S.; Habermann, F. Molecular cytogenetic definition of the chicken genome: Te first complete avian karyotype. Genetics 2004, 166, 1367–1373. [Google Scholar] [CrossRef] [PubMed]
- Lewin, H.A.; Larkin, D.M.; Pontius, J.; O’Brien, S.J. Every genome sequence needs a good map. Genome Res. 2009, 19, 1925–1928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, L.; Georges, M. Domestic-animal genomics: Deciphering the genetics of complex traits. Nat. Rev. Genet. 2004, 5, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Damas, J.; O’Connor, R.; Farré, M.; Lenis, V.P.E.; Martell, H.J.; Mandawala, A.; Larkin, D.M. Upgrading short-read animal genome assemblies to chromosome level using comparative genomics and a universal probe set. Genome Res. 2017, 27, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Griffin, D.K.; Robertson, L.B.W.; Tempest, H.G.; Skinner, B.M. The evolution of the avian genome as revealed by comparative molecular cytogenetics. Cytogenet. Genome Res. 2007, 117, 64–77. [Google Scholar] [CrossRef] [PubMed]
- De Boer, L.E.M. Karyological heterogeneity in the Falconiformes (aves). Experientia 1975, 31, 1138–1139. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, S.M.; Oliphant, L.W. Chromosome study of peregrine, prairie, and gyrfalcons with implications for hybrids. J. Hered. 1987, 78, 388–390. [Google Scholar] [CrossRef]
- Amaral, K.; Felipe, K.; Jorge, W. The chromosomes of the Order Falconiformes: A review. Ararajuba 2003, 11, 65–73. [Google Scholar]
- Sibley, C.G.; Charles, G.; Ahlquist, J.E. Phylogeny and Classification of Birds: A study in Molecular Evolution; Yale University Press: New Haven, CT, USA, 1990. [Google Scholar]
- Del Hoyo, J.; Elliott, A.; Christie, D. Cotingas to Pipits and Wagtails. In Handbook of the Birds of the World; Oxford University Press: Oxford, UK, 1992. [Google Scholar]
- Itoh, Y.; Arnold, A.P. Chromosomal polymorphism and comparative painting analysis in the zebra finch. Chromosome Res. 2005, 13, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Belterman, R.H.R.; De Boer, L.E.M. A karyological study of 55 species of birds, including karyotypes of 39 species new to cytology. Genetica 1984, 65, 39–82. [Google Scholar] [CrossRef]
- De Boer, L.E.M. The somatic chromosome complements of 16 species of falconiformes (Aves) and the karyological relationships of the order. Genetica 1990, 46, 77–113. [Google Scholar] [CrossRef]
- Longmire, J.L.; Lewis, A.K.; Brown, N.C.; Buckingham, J.M.; Clark, L.M.; Jones, M.D.; Ray, F.A. Isolation and molecular characterization of a highly polymorphic centromeric tandem repeat in the family Falconidae. Genomics 1988, 2, 14–24. [Google Scholar] [CrossRef]
- Nishida, C.; Ishijima, J.; Kosaka, A.; Tanabe, H.; Habermann, F.A.; Griffin, D.K.; Matsuda, Y. Characterization of chromosome structures of Falconinae (Falconidae, Falconiformes, Aves) by chromosome painting and delineation of chromosome rearrangements during their differentiation. Chromosome Res. 2008, 16, 171–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, X.; Pan, S.; Wang, J.; Dixon, A.; He, J.; Muller, M.G.; Bruford, M.W. Peregrine and saker falcon genome sequences provide insights into evolution of a predatory lifestyle. Nat. Genet. 2013, 45, 563–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connor, R.E.; Farre, M.; Joseph, S.; Damas, J.; Kiazim, L.G.; Jennings, R.; Griffin, D.K. Chromosome-level assembly reveals extensive rearrangement in saker falcon, budgerigar but not ostrich genomes. Genome Biol. 2018, in press. [Google Scholar]
- Li, R.; Zhu, H.; Ruan, J.; Qian, W.; Fang, X.; Shi, Z.; Wang, J. De novo assembly of human genomes with massively parallel short read sequencing. Genome Res. 2010, 20, 265–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Rahbek, C.; Graves, G.R.; Lei, F.; Jarvis, E.D.; Gilbert, M.T.P. Genomics: Bird sequencing project takes off. Nature 2015, 522, 34. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, R.E.E.; Fonseka, G.; Frodsham, R.; Archibald, A.L.L.; Lawrie, M.; Walling, G.A.A.; Griffin, D.K.K. Isolation of subtelomeric sequences of porcine chromosomes for translocation screening reveals errors in the pig genome assembly. Anim. Genet. 2017, 48, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Gao, Y.; Feng, C.; Liu, Q.; Wang, X.; Du, Z.; Li, N. Advanced technologies for genomic analysis in farm animals and its application for QTL mapping. Genetica 2009, 136, 371–386. [Google Scholar] [CrossRef] [PubMed]
- Farré, M.; Narayan, J.; Slavov, G.T.; Damas, J.; Auvil, L.; Li, C.; Larkin, D.M. Novel Insights into Chromosome Evolution in Birds, Archosaurs, and Reptiles. Genome Biol. Evol. 2016, 8, 2442–2451. [Google Scholar] [CrossRef] [PubMed]
- Helbig, A.J.; Seibold, I.; Bednarek, W.; Brüning, H.; Gaucher, P.; Ristow, D.; Wink, M. Phylogenetic relation-ships among Falcon species (genus Falco) according to DNA sequence variation of the cytochrome b gene. Raptor Conserv. Today 1994, 593–599. [Google Scholar]
- Nittinger, F.; Haring, E.; Pinsker, W.; Wink, M.; Gamauf, A. Out of Africa? Phylogenetic relationships between Falco biarmicus and the other hierofalcons (Aves: Falconidae). J. Zool. Syst. Evol. Res. 2005, 43, 321–331. [Google Scholar] [CrossRef]
- Nittinger, F.; Gamauf, A.; Pinsker, W.; Wink, M.; Haring, E. Phylogeography and population structure of the saker falcon (Falco cherrug) and the influence of hybridization: Mitochondrial and microsatellite data. Mol. Ecol. 2007, 16, 1497–1517. [Google Scholar] [CrossRef] [PubMed]
- Eastham, C.P.; Nicholls, M.K. Morphometric Analysis of Large Falco Species and Their Hybrids With Implications for Conservation. J. Raptor Res. 2005, 39, 386–393. [Google Scholar]
- Heidenreich, M. Birds of Prey: Medicine and Management; Blackwell Science: Oxford, UK, 1997. [Google Scholar]
- de Oliveira Furo, I.; Kretschmer, R.; O’Brien, P.C.; Ferguson-Smith, M.A.; de Oliveira, E.H.C. Chromosomal Diversity and Karyotype Evolution in South American Macaws (Psittaciformes, Psittacidae). PLoS ONE 2015, 10, e0130157. [Google Scholar] [CrossRef] [PubMed]
- Nie, W.; O’Brien, P.C.M.; Fu, B.; Wang, J.; Su, W.; He, K.; Baker, R. Multidirectional chromosome painting substantiates the occurrence of extensive genomic reshuffling within Accipitriformes. BMC Evol. Biol. 2015, 15, 205. [Google Scholar] [CrossRef] [PubMed]
- Seibold-Torres, C.; Owens, E.; Chowdhary, R.; Ferguson-Smith, M.A.; Tizard, I.; Raudsepp, T. Comparative Cytogenetics of the Congo African Grey Parrot (Psittacus erithacus). Cytogenet. Genome Res. 2016, 147, 144–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Li, C.; Li, Q.; Li, B.; Larkin, D.M.; Lee, C.; Froman, D.P. Comparative genomics reveals insights into avian genome evolution and adaptation. Science 2014, 346, 1311–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romanov, M.N.; Farré, M.; Lithgow, P.E.; Fowler, K.E.; Skinner, B.M.; O’Connor, R.; Nishida, C. Reconstruction of gross avian genome structure, organization and evolution suggests that the chicken lineage most closely resembles the dinosaur avian ancestor. BMC Genom. 2014, 15, 1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joseph, S.; O’Connor, R.E.; Al Mutery, A.F.; Watson, M.; Larkin, D.M.; Griffin, D.K. Chromosome Level Genome Assembly and Comparative Genomics between Three Falcon Species Reveals an Unusual Pattern of Genome Organisation. Diversity 2018, 10, 113. https://doi.org/10.3390/d10040113
Joseph S, O’Connor RE, Al Mutery AF, Watson M, Larkin DM, Griffin DK. Chromosome Level Genome Assembly and Comparative Genomics between Three Falcon Species Reveals an Unusual Pattern of Genome Organisation. Diversity. 2018; 10(4):113. https://doi.org/10.3390/d10040113
Chicago/Turabian StyleJoseph, Sunitha, Rebecca E. O’Connor, Abdullah F. Al Mutery, Mick Watson, Denis M. Larkin, and Darren K. Griffin. 2018. "Chromosome Level Genome Assembly and Comparative Genomics between Three Falcon Species Reveals an Unusual Pattern of Genome Organisation" Diversity 10, no. 4: 113. https://doi.org/10.3390/d10040113
APA StyleJoseph, S., O’Connor, R. E., Al Mutery, A. F., Watson, M., Larkin, D. M., & Griffin, D. K. (2018). Chromosome Level Genome Assembly and Comparative Genomics between Three Falcon Species Reveals an Unusual Pattern of Genome Organisation. Diversity, 10(4), 113. https://doi.org/10.3390/d10040113