Influence of Landscape Factors on Amphibian Roadkills at the National Level



Abstract
:1. Introduction
2. Methods
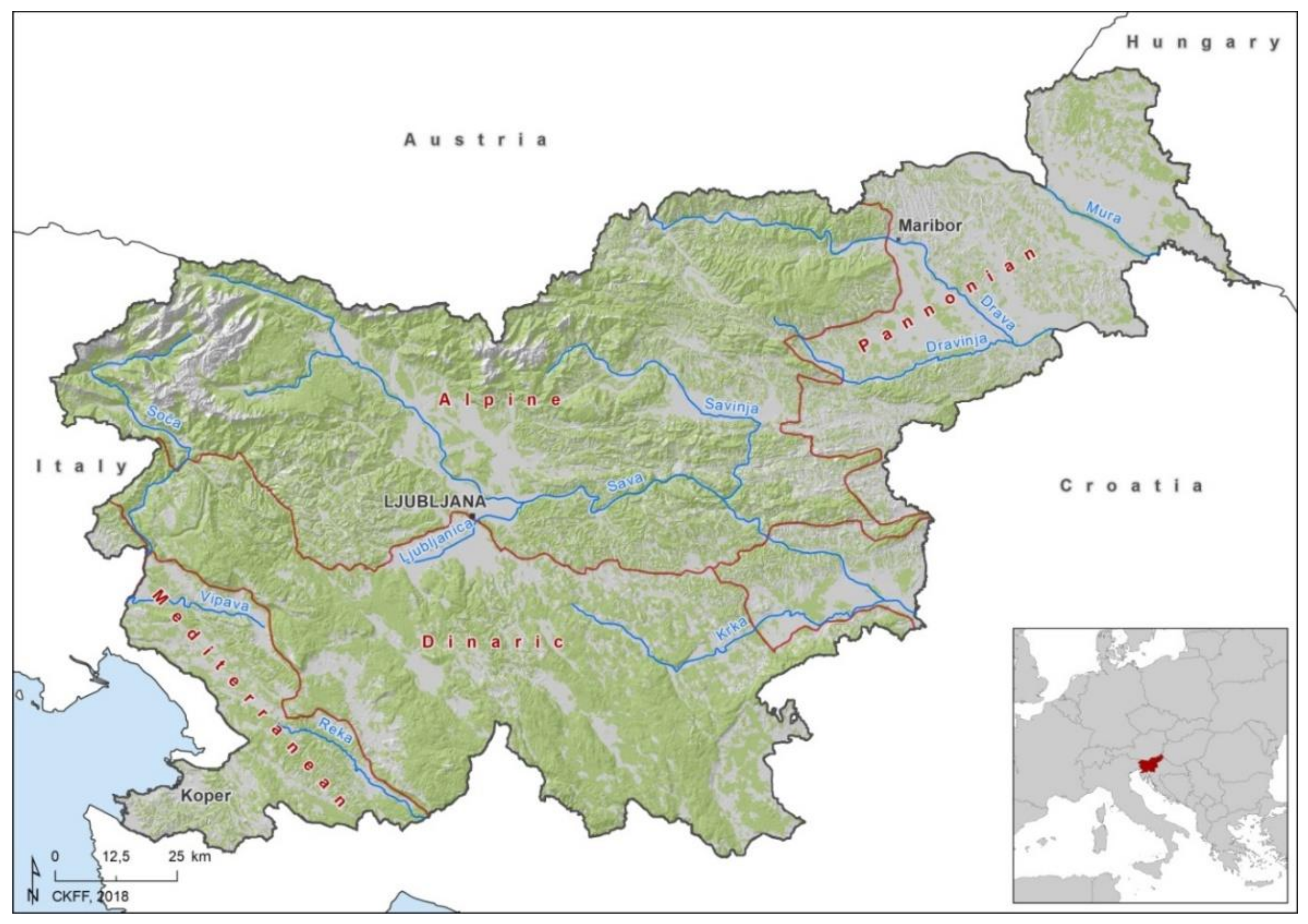
2.1. Study Area
2.2. Data Collection
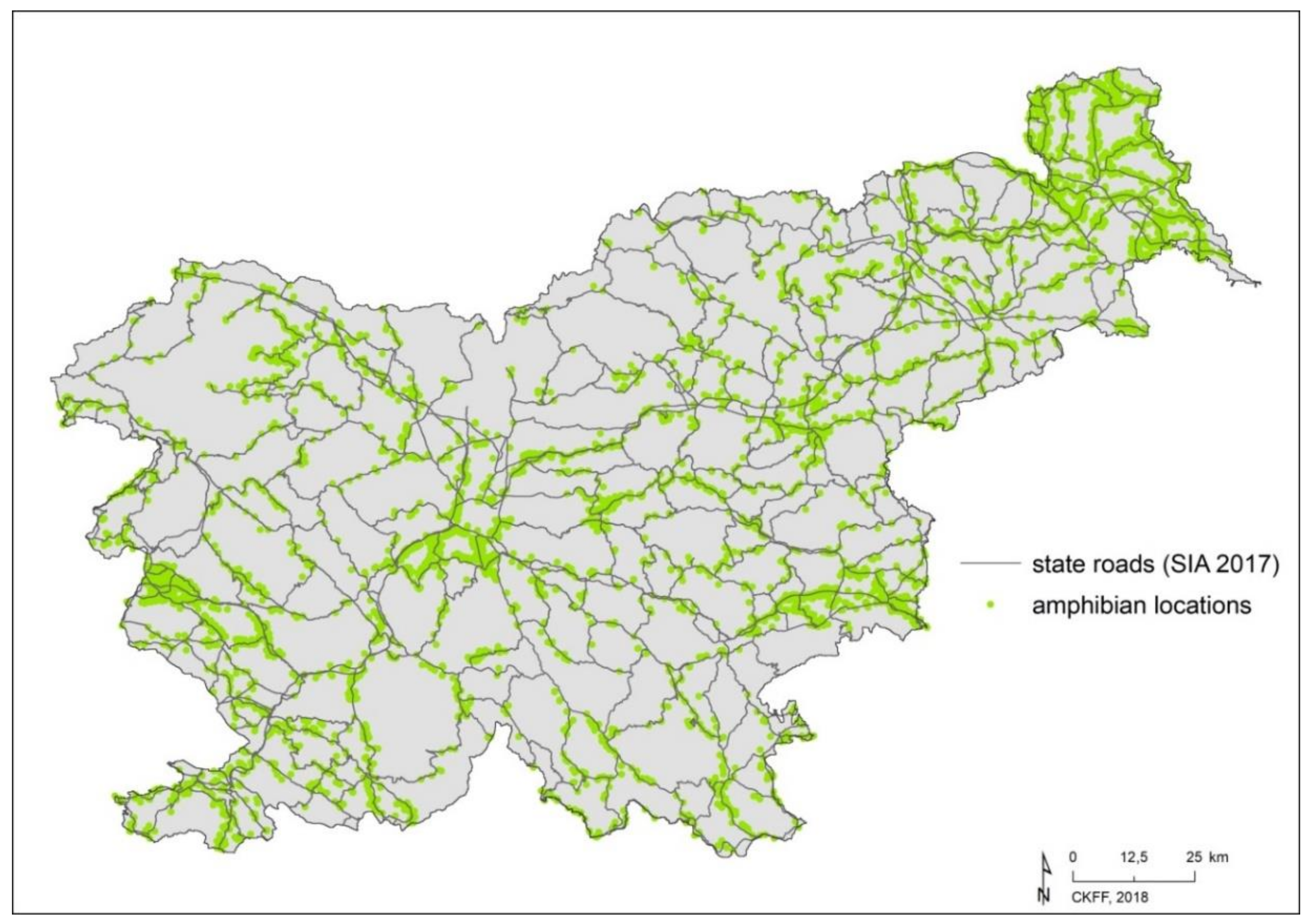
2.2.1. Amphibian Data
2.2.2. State Roads and Land Use
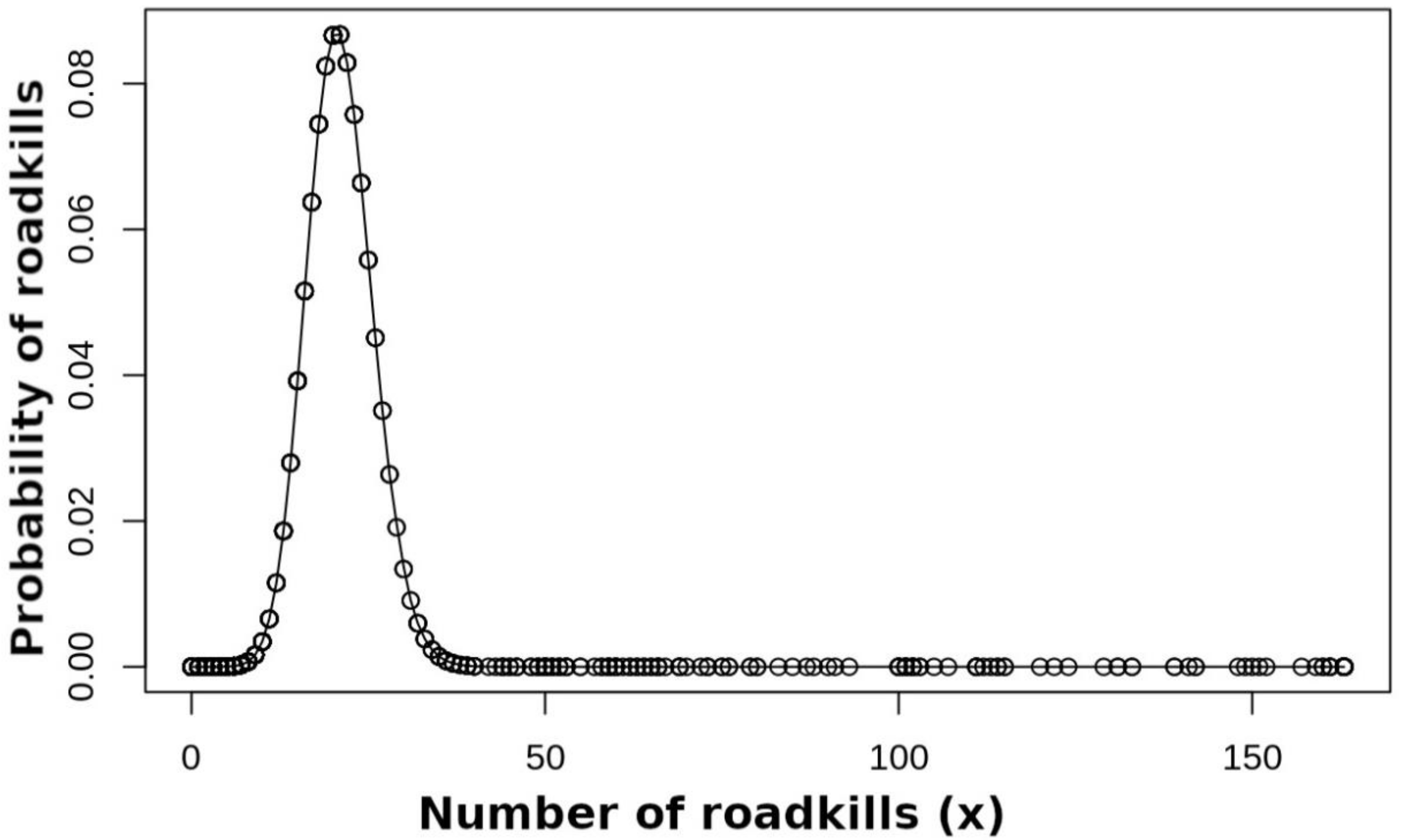
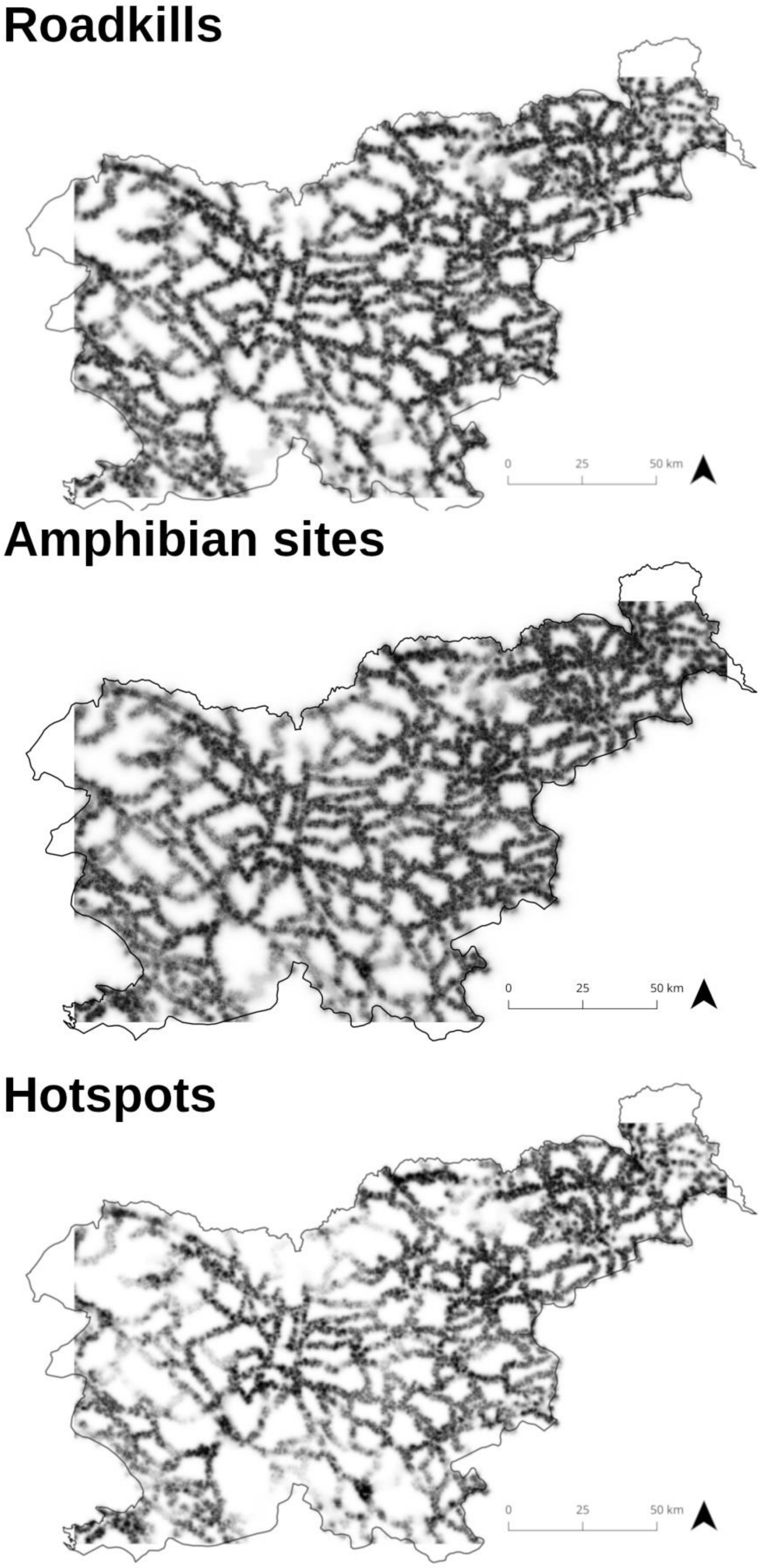
2.2.3. Determination of Roadkill Hotspots
2.3. Ecological Niche Models
2.4. GLM for Number of Roadkills
3. Results
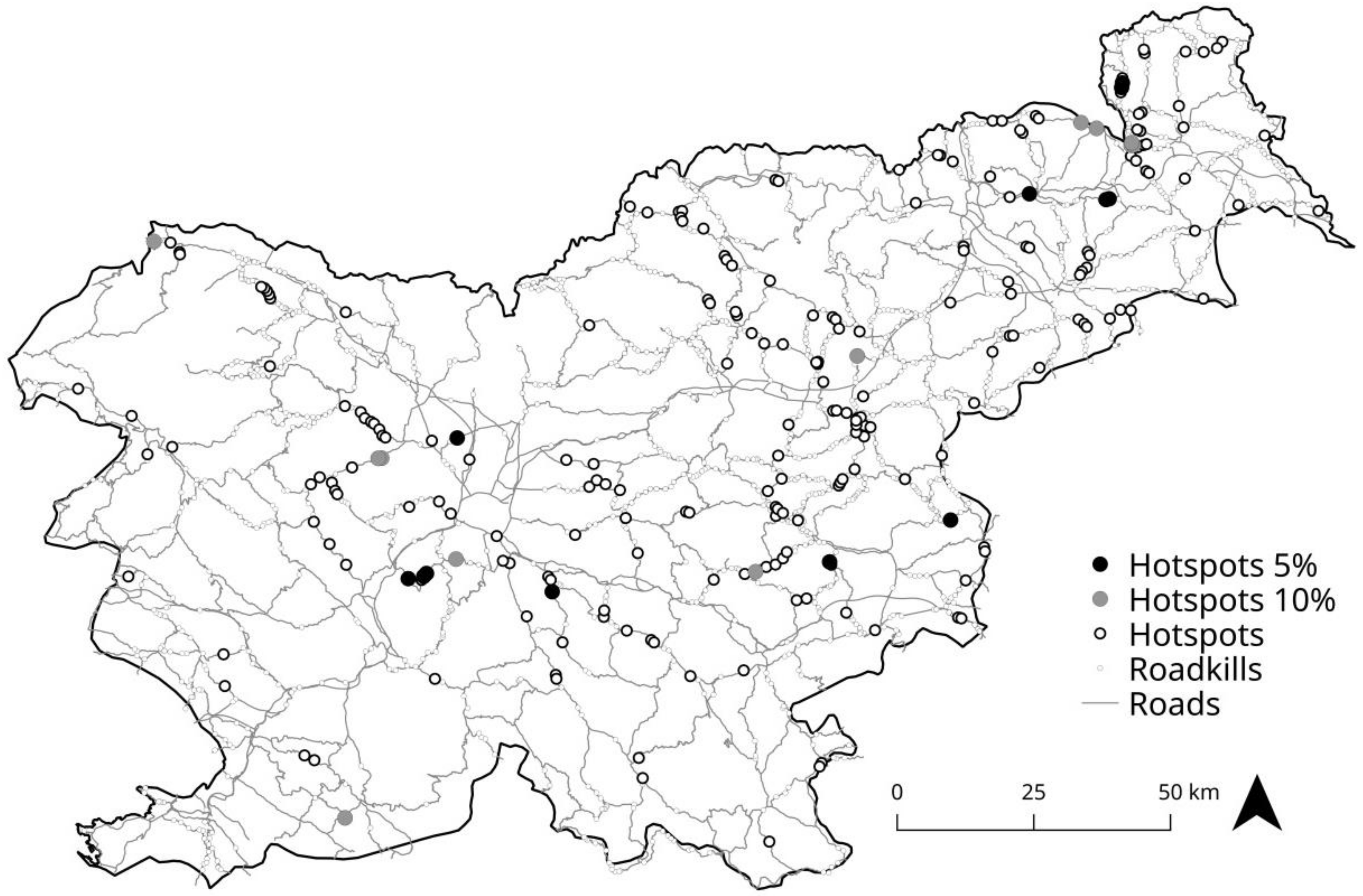
3.1. Determination of Roadkill Hotspots
3.2. Ecological Niche Models
3.3. Generalised Linear Models
4. Discussion
4.1. Determination of Roadkill Hotspots
4.2. Ecological Niche Modelling of Roadkills and Hotspots
4.3. Modelling of Number Of Roadkills and Hotspots
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Torres, A.; Jaeger, J.A.G.; Carlos, J. Assessing large-scale wildlife responses to human infrastructure development. Proc. Natl. Acad. Sci. USA 2016, 113, 8472–8477. [Google Scholar] [CrossRef] [PubMed]
- Clarke, G.P.; White, P.C.L.; Harris, S. Effects of roads on badger Meles meles populations in south-west England. Biol. Conserv. 1998, 86, 117–124. [Google Scholar] [CrossRef]
- Forman, R.T.T.; Alexander, L.E. Roads and their major ecological effects. Annu. Rev. Ecol. Syst. 1998, 29, 207–231. [Google Scholar] [CrossRef]
- Fahrig, L.; Pedlar, J.H.; Pope, S.E.; Taylor, P.D.; Wegner, J.F. Effect of road traffic on amphibian density. Biol. Conserv. 1995, 73, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Glista, D.J.; Devault, T.L.; Dewoody, J.A. Vertebrate road mortality predominantly impacts amphibians. Herpetol. Conserv. Biol. 2008, 3, 77–87. [Google Scholar]
- Gunther, K.A.; Biel, M.J.; Robison, H.L. Factors influencing the frecuency of road-killed wildlife in Yellowstone National Park. ICOWET 1998, 12, 32–42. [Google Scholar]
- Hels, T.; Buchwald, E. The effect of road kills on amphibian populations. Biol. Conserv. 2001, 3, 331–340. [Google Scholar] [CrossRef]
- Clevenger, A.P.; Chruszcz, B.; Gunson, K.E. Spatial patterns and factors influencing small vertebrate fauna road-kill aggregations. Biol. Conserv. 2003, 109, 15–26. [Google Scholar] [CrossRef]
- Saeki, M.; Macdonald, D.W. The effects of traffic on the raccoon dog (Nyctereutes procyonoides viverrinus) and other mammals in Japan. Biol. Conserv. 2004, 5, 559–571. [Google Scholar] [CrossRef]
- Santos, X.; Llorente, G.A.; Montori, A.; Carretero, M.A.; Franch, M.; Garriga, N.; Richter-Boix, A. Evaluating factors affecting amphibian mortality on roads: The case of the Common Toad, Bufo bufo near a breeding place. Anim. Biodiv. Conserv. 2008, 1, 97–104. [Google Scholar]
- Matos, C.; Sillero, N.; Argaña, E. Spatial analysis of amphibian road mortality levels in northern Portugal country roads. Amphib. Rept. 2012, 33, 469–483. [Google Scholar] [CrossRef]
- Langley, W.M.; Lipps, H.W.; Theis, J.F. Responses of Kansas Motorists to Snake Models on a Rural Highway. Trans. Kans. Acad. Sci. 1989, 92, 43–48. [Google Scholar] [CrossRef]
- Paul Ashley, E.; Kosloski, A.; Petrie, S.A. Incidence of Intentional Vehicle–Reptile Collisions. Hum. Dimens. Wildl. 2007, 12, 137–143. [Google Scholar] [CrossRef]
- Secco, H.; Ratton, P.; Castro, E.; da Lucas, P.S.; Bager, A. Intentional Snake Road-Kill: A Case Study using Fake Snakes on a Brazilian Road. Trop. Conserv. Sci. 2014, 7, 561–571. [Google Scholar] [CrossRef] [Green Version]
- Garriga, N.; Santos, X.; Montori, A. Are protected areas truly protected? The impact of road traffic on vertebrate fauna. Biodiv. Conser. 2012, 21, 2761–2774. [Google Scholar] [CrossRef]
- Crist, M.R.; Wilmer, B.; Aplet, G.H. Assessing the value of roadless areas in a conservation reserve strategy: Biodiversity and landscape connectivity in the northern Rockies. J. Appl. Ecol. 2005, 42, 181–191. [Google Scholar] [CrossRef]
- Chen, X.; Roberts, K. Roadless areas and biodiversity: A case study in Alabama, USA. Biodiv. Conserv. 2008, 17, 2013–2022. [Google Scholar] [CrossRef]
- Selva, N.; Kreft, S.; Kati, V.; Schluck, M.; Jonsson, B.-G.; Mihok, B.; Okarma, H.; Ibisch, P.L. Roadless and Low-Traffic Areas as Conservation Targets in Europe. Env. Manag. 2011, 48, 865–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibisch, P.; Hoffmann, M.T.; Kreft, S.; Pe’er, G.; Kati, V.; Biber-Freudenberger, L.; DellaSala, D.A.; Vale, M.M.; Hobson, P.R.; Selva, N. A global map of roadless areas and their conservation status. Science 2016, 354, 1423–1427. [Google Scholar] [CrossRef]
- Santos, S.M.; Lourenço, R.; Mira, A.; Beja, P. Relative effects of road risk, habitat suitability, and connectivity on wildlife roadkills: The case of tawny owls (Strix aluco). PLoS ONE 2013, 8, e79967. [Google Scholar] [CrossRef]
- Sillero, N. Amphibian mortality levels on Spanish country roads: Descriptive and spatial analysis. Amphib. Rept. 2008, 29, 337–347. [Google Scholar] [CrossRef]
- Ha, H.; Shilling, F. Modelling potential wildlife-vehicle collisions (WVC) locations using environmental factors and human population density: A case-study from 3 state highways in Central California. Ecol. Inform. 2018, 43, 212–221. [Google Scholar] [CrossRef]
- Carr, L.W.; Fahrig, L. Effect of Road Traffic on Two Amphibian Species of Differing Vagility. Conserv. Biol. 2001, 4, 1071–1078. [Google Scholar] [CrossRef]
- Mazerolle, M.J.; Huot, M.; Gravel, M. Behavior of amphibians on the road in response to car traffic. Herpetologica 2005, 4, 380–388. [Google Scholar] [CrossRef]
- Orlowski, G. Spatial distribution and seasonal pattern in road mortality of the common toad Bufo bufo in an agricultural landscape of south-western Poland. Amphib. Rept. 2007, 1, 25–31. [Google Scholar] [CrossRef]
- Trombulak, S.C.; Frissell, C.A. Review of Ecological Effects of Roads on Terrestrial and Aquatic Communities. Conserv. Biol. 2000, 1, 18–30. [Google Scholar] [CrossRef]
- Jaeger, J.A.G.; Fahrig, L. Effects of Road Fencing on Population Persistence. Conserv. Biol. 2004, 6, 1651–1657. [Google Scholar] [CrossRef]
- Jaeger, J.A.G.; Bowman, J.; Brennan, J.; Fahrig, L.; Bert, D.; Bouchard, J.; Charbonneau, N.; Frank, K.; Gruber, B.; von Toschanowitz, K.T. Predicting when animal populations are at risk from roads: An interactive model of road avoidance behavior. Ecol. Model. 2005, 2–4, 329–348. [Google Scholar] [CrossRef]
- Grilo, C.; Bissonette, J.A.; Santos-Reis, M. Spatial-temporal patterns in Mediterranean carnivore road casualties: Consequences for mitigation. Biol. Conserv. 2009, 142, 301–313. [Google Scholar] [CrossRef]
- Ascensão, F.; Clevenger, A.; Santos-Reis, M.; Urbano, P.; Jackson, N. Wildlife–vehicle collision mitigation: Is partial fencing the answer? An agent-based model approach. Ecol. Model. 2013, 257, 36–43. [Google Scholar] [CrossRef]
- Bager, A.; Fontoura, V. Evaluation of the effectiveness of a wildlife roadkill mitigation system in wetland habitat. Ecol. Eng. 2013, 53, 31–38. [Google Scholar] [CrossRef]
- Malo, J.E.; Suáez, F.; Díez, A. Can we mitigate animal-vehicle accidents using predictive models? J. Appl. Ecol. 2004, 4, 701–710. [Google Scholar] [CrossRef]
- Gomes, L.; Grilo, C.; Silva, C.; Mira, A. Identification methods and deterministic factors of owl roadkill hotspot locations in Mediterranean landscapes. Ecol. Res. 2009, 24, 355–370. [Google Scholar] [CrossRef]
- Ramp, D.; Caldwell, J.; Edwards, K.A.; Warton, D.; Croft, D.B. Modelling of wildlife fatality hotspots along the Snowy Mountain Highway in New South Wales, Australia. Biol. Conserv. 2005, 126, 474–490. [Google Scholar] [CrossRef]
- Teixeira, F.Z.; Coelho, I.P.; Esperandio, I.B.; Oliveira, N.R.; Peter, F.P.; Dornelles, S.S.; Delazeri, N.R.; Tavares, M.; Martins, M.B.; Kindel, A. Are Road-Kill Hotspots Coincident Among Different Vertebrate Groups? Oecol. Aust. 2013, 17, 36–47. [Google Scholar] [CrossRef]
- Shilling, F.M.; Waetjen, D.P. Wildlife-vehicle collision hotspots at US highway extents: Scale and data source effects. Nat. Conserv. 2015, 11, 41–60. [Google Scholar] [CrossRef]
- Poboljšaj, K.; Kotarac, M.; Lešnik, A.; Šalamun, A.; Grobelnik, V.; Jakopič, M. Dvoživke in ceste. Končno poročilo. In Amphibians and Roads. Final Report; Center za kartografijo favne in flore: Miklavž na Dravskem polju, Slovenia, 2000; 73p. [Google Scholar]
- Project Dvoživke in promet v regiji Alpe-Jadran (Amphibians and Traffic in Alps-Adriatic Region). 2003. Available online: http://www.ckff.si/DvoCeste/Aktivnosti.htm (accessed on 1 March 2018).
- Interreg III A Slovenia-Austria Project Varstvo dvoživk in Netopirjev v Regiji Alpe-Jadran (Amphibian and Bat Conservation in Alps-Adriatic region). 2005–2007. Available online: http://www.ckff.si/projekt.php?pid=4 (accessed on 1 March 2018).
- Poboljšaj, K.; Lešnik, A.; Grobelnik, V.; Šalamun, A.; Kotarac, M. Predlog ukrepov za zaščito dvoživk na cestah v upravljanju DRSI. Končno poročilo. In Proposal for Amphibian Protection Measures on Roads Managed by Slovenian Infrastructure Agency; Final report; Center za kartografijo favne in flore: Miklavž na Dravskem polju, Slovenia, 2018; 95p, Available online: http://www.ckff.si/javno/projekti/12874.pdf (accessed on 1 March 2018).
- Poboljšaj, K. Dvoživke in ceste: Dvoživke potrebujejo našo zaščito in pomoč. Trdoživ 2013, 1, 32–33. [Google Scholar]
- Herpetološko Društvo—Societas Herpetologica Slovenica, Žabohod: Posvet o Naravovarstveni Problematiki Dvoživk in Cest. 2014. Available online: https://sites.google.com/site/zabohodposvet/home (accessed on 1 March 2018).
- Društvo Žverca, Pomagajmo Žabicam. 2018. Available online: https://www.pomagajmo-zabicam.si/ (accessed on 1 March 2018).
- Stanković, D.; Lužnik, M.; Poboljšaj, K. Conservation and declines of amphibians in Slovenia. In Amphibian Biology, Status of Conservation and Decline of Amphibians: Eastern Hemisphere. Part 4, Southern Europe and Turkey; Heathwole, A., Wilkinson, J.W., Eds.; Pelagic Publishing: Exeter, UK, 2015; Volume 11, pp. 42–53. ISBN 978-1-907807-53-4. [Google Scholar]
- Anthony, B.; Arntzen, J.W.; Baha El Din, S.; Böhme, W.; Cogălniceanu, D.; Crnobrnja-Isailovic, J.; Crochet, P.-A.; Corti, C.; Griffiths, R.; Kaneko, Y.; et al. Amphibians of the Palaearctic Realm. In Threatened Amphibians of the World; Stuart, S.N., Hoffmann, M., Chanson, J.S., Cox, N.A., Berridge, R.J., Ramani, P., Young, B.E., Eds.; Lynx Edicions with IUCN—The World Conservation Union, Conservation International and Nature Serve: Barcelona, Spain, 2008; pp. 106–111. [Google Scholar]
- Perko, D.; Orožen Adamič, M.; Belec, B.; Fridl, J.; Gabrovec, M.; Hrvatin, M.; Kert, B.; Kladnik, D.; Lovrenčak, F.; Mihelič, L.; et al. Slovenija. Pokrajina in Ljudje, 2nd ed.; Mladinska knjiga: Ljubljana, Slovenia, 1999; 736p. [Google Scholar]
- Verovnik, R.; Rebeušek, F.; Jež, M. Atlas Dnevnih Metuljev (Lepidoptera: Rhopalocera) Slovenije, Atlas of Butterflies (Lepidoptera: Rhopalocera) of Slovenia. Atlas faunae et Florae Sloveniae 3; Center Za Kartografijo Favne in Flore: Miklavž na Dravskem Polju, Slovenia, 2012; 456p, ISBN 978-961-93280-0-2. [Google Scholar]
- Presetnik, P.; Koselj, K.; Zagmajster, M.; Zupančič, N.; Jazbec, K.; Žibrat, U.; Petrinjak, A.; Hudoklin, A. Atlas Netopirjev (Chiroptera) Slovenije, Atlas of bats (Chiroptera) of Slovenia. Atlas Faunae et florae Sloveniae 2; Center za Kartografijo Favne in Flore: Miklavž na Dravskem polju, Slovenia, 2009; 152p, ISBN 978-961-9051290. [Google Scholar]
- Environmental Indicators in Slovenia: [TP01] Land Cover and Land Use (Kušar). 2015. Available online: http://kazalci.arso.gov.si/?data=indicator&ind_id=671&lang_id=94 (accessed on 1 July 2018).
- Slovenian Infrastructure Agency. Roads. Available online: http://www.di.gov.si/en/ (accessed on 1 July 2018).
- Shaffer, H.B.; Juterbock, J.E. Night driving. In Measuring and Monitoring Biological Diversity. Standard Methods for Amphibians; Heyer, W.R., Donnelly, M.A., McDiarmid, R.W., Hayek, L.A., Foster, M.S., Eds.; Smithsonian Institution Press: Washington, DC, USA; London, UK, 1994; pp. 163–166. ISBN 10-1560982845. [Google Scholar]
- SIA-Slovenian Infrastructure Agency. State Roads Vector Data (*shp); Slovenian Infrastructure Agency: Ljubljana, Slovenia, 2017. [Google Scholar]
- The Spatial Records of Actual Use of Agricultural and Forest Land (Ministry of Agriculture, Forestry and Food) in 2017. Available online: http://rkg.gov.si/GERK/ (accessed on 1 March 2018).
- Sillero, N. What does ecological modelling model? A proposed classification of ecological niche models based on their underlying methods. Ecol. Model. 2011, 222, 1343–1346. [Google Scholar] [CrossRef]
- Barbosa, M.A.; Sillero, N.; Martínez-Freiría, F.; Real, R. Ecological niche models in Mediterranean herpetology: Past, present and future. In Ecological Modelling; Zhang, W.J., Ed.; Chapter 8; Nova Science Publishers: Hauppauge, NY, USA, 2012; pp. 173–204. ISBN 978-1-61324-567-5. [Google Scholar]
- Phillips, S.J.; Dudík, M.; Schapire, R.E. A maximum entropy approach to species distribution modeling. In Proceedings of the Twenty-First International Conference on Machine Learning, Banff, AB, Canada, 4–8 July 2004; ACM: New York, NY, USA, 2004; pp. 655–662. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Elith, J.; Graham, C.H.; Lehmann, A.; Leathwick, J.; Ferrier, S. Sample selection bias and presence-only distribution models: Implications for background and pseudo-absence data. Ecol. Appl. 2009, 19, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Sillero, N.; Carretero, M.A. Modelling the past and future distribution of contracting species. The Iberian lizard Podarcis carbonelli (Squamata: Lacertidae) as a case study. J. Comp. Zool. 2013, 252, 289–298. [Google Scholar] [CrossRef]
- Maxent 3.4.1 Software. Available online: http://biodiversityinformatics.amnh.org/open_source/maxent/ (accessed on 15 January 2019).
- Liu, C.; Berry, P.M.; Dawson, T.P.; Pearson, R.G. Selecting thresholds of occurrence in the prediction of species distributions. Ecography 2005, 28, 385–393. [Google Scholar] [CrossRef] [Green Version]
- Van Der Wal, J.; Shoo, L.P.; Graham, C.; Williams, S.E. Selecting pseudo-absence data for presence-only distribution modeling: How far should you stray from what you know? Ecol. Model. 2009, 220, 589–594. [Google Scholar] [CrossRef]
- Lobo, J.M.; Jiménez-Valverde, A.; Real, R. AUC: A misleading measure of the performance of predictive distribution models. Glob. Ecol. Biogeogr. 2008, 17, 145–151. [Google Scholar] [CrossRef]
- Raes, N.; ter Steege, H. A null-model for significance testing of presence-only species distribution models. Ecography 2007, 30, 727–736. [Google Scholar] [CrossRef]
- Martínez-Freiría, F.; Brito, J.C. Quantification of road mortality for amphibians and reptiles in Hoces del Alto Ebro y Rudrón Natural Park in 2005. Basic Appl. Herpetol. 2012, 26, 33–42. [Google Scholar] [CrossRef]
- Eberhardt, E.; Mitchell, S.; Fahrig, L. Road kill hotspots do not effectively indicate mitigation locations when past road kill has depressed populations. J. Wildl. Manag. 2013, 77, 1353–1359. [Google Scholar] [CrossRef]
- Iosif, R.; Popescu, V.D. Modeling road mortality hotspots of Eastern Hermann’s tortoise in Romania. Amphib. Rept. 2013, 3, 1–10. [Google Scholar] [CrossRef]
- Taylor, B.D.; Goldingay, R.L. Wildlife road-kills on three major roads in north-eastern New South Wales. Wildl. Res. 2004, 1, 83–91. [Google Scholar] [CrossRef]
- Garrote, G.; Fernández-López, J.; López, G.; Ruiz, G.; Simón, M.A. Prediction of iberian lynx road–mortality in southern Spain: A new approach using the MaxEnt algorithm. Am. Biodivers. Conserv. 2018, 41, 217–225. [Google Scholar] [CrossRef]
- Naderi, M.; Farashi, A.; Erdi, M.A. Persian leopard’s (Panthera pardus saxicolor) unnatural mortality factors analysis in Iran. PLoS ONE 2018, 13, e0195387. [Google Scholar] [CrossRef] [PubMed]
- Seiler, A. Predicting locations of moose-vehicle collisions in Sweden. J. Appl. Ecol. 2005, 2, 371–382. [Google Scholar] [CrossRef]
- Kramer-Schadt, S.; Revilla, E.; Wiegand, T.; Breitenmoser, U. Fragmented landscapes, road mortality and patch connectivity: Modelling influences on the dispersal of Eurasian lynx. J. Appl. Ecol. 2004, 4, 711–723. [Google Scholar] [CrossRef]
- Nielsen, S.E.; Herrero, S.; Boyce, M.S.; Mace, R.D.; Benn, B.; Gibeau, M.L.; Jevons, S. Modelling the spatial distribution of human-caused grizzly bear mortalities in the Central Rockies ecosystem of Canada. Biol. Conserv. 2004, 1, 101–113. [Google Scholar] [CrossRef]
- D’Amico, M.; Román, J.; de los Reyes, L.; Revilla, E. Vertebrate road-kill patterns in Mediterranean habitats: Who, when and where. Biol. Conserv. 2015, 191, 234–242. [Google Scholar] [CrossRef]
- Ramp, D.; Wilson, V.K.; Croft, D.B. Assessing the impacts of roads in peri-urban reserves: Road-based fatalities and road usage by wildlife in the Royal National Park, New South Wales, Australia. Biol. Conserv. 2006, 129, 348–359. [Google Scholar] [CrossRef]
- Schmidt, B.R.; Zumbach, S. Amphibian Road Mortality and How to Prevent It: A Review. In Urban Herpetology; Mitchell, J.C., Jung Brown, R.E., Bartolomew, B., Eds.; Society for the Study of Amphibians and Reptiles: St. Louis, MI, USA; pp. 157–167.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Training | Training Mean AUC | Training SD AUC | Test | Test Mean AUC | Test SD AUC |
|---|---|---|---|---|---|---|
| Hotspots | 159 | 0.90 | 0.01 | 68 | 0.89 | 0.02 |
| Amphibian sites | 2594 | 0.79 | 0.01 | 1111 | 0.79 | 0.01 |
| Roadkills | 1008 | 0.84 | 0.01 | 432 | 0.84 | 0.01 |
| Variables | Agriculture | Forest | Meadows | Urban | Water | Wetlands |
| Hotspots | 29.96 | 21.18 | 30.23 | 4.63 | 9.71 | 4.29 |
| Amphibian sites | 26.18 | 50.99 | 9.25 | 9.61 | 1.69 | 2.29 |
| Roadkills | 9.25 | 28.40 | 49.41 | 9.79 | 2.47 | 0.69 |
| Variables | Roadkills | Hotspots |
|---|---|---|
| Traffic | −0.07 | −0.02 |
| Agriculture | 0.04 | 0.02 |
| Forest | 0.02 | 0.01 |
| Meadows | −0.05 | −0.01 |
| Urban | −0.03 | −0.01 |
| Water | 0.01 | 0 |
| Wetlands | −0.06 | −0.02 |
| Roadkills | Estimate | Std. Error | t value | p value |
|---|---|---|---|---|
| Intercept | 3.282 | 0.130 | 25.172 | <0.001 *** |
| Traffic | −0.001 | 0.000 | −11.678 | <0.001 *** |
| Agriculture | 0.304 | 0.369 | 0.822 | 0.411 |
| Forest | 175.600 | 56.150 | 3.126 | <0.001 *** |
| Meadows | −67.760 | 60.860 | −1.113 | 0.266 |
| Urban | −123.100 | 227.600 | −0.541 | 0.7789 |
| Water | 48.150 | 7.421 | 6.489 | <0.001 *** |
| Wetlands | −3.833 | 1.083 | −3.540 | <0.001 *** |
| Hotspots | Estimate | Std. Error | t value | p value |
| Intercept | 5.246 | 0.407 | 12.879 | <0.001 *** |
| Traffic | −0.002 | 0.000 | −7.735 | <0.001 *** |
| Agriculture | 0.467 | 1.072 | 0.436 | 0.663 |
| Forest | 258.000 | 146.800 | 1.757 | 0.079 |
| Meadows | −206.600 | 159.700 | −1.293 | 0.196 |
| Urban | 1201.000 | 800.000 | 1.501 | 0.133 |
| Water | 54.750 | 21.750 | 2.518 | 0.012 * |
| Wetlands | −2.467 | 2.989 | −0.825 | 0.409 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sillero, N.; Poboljšaj, K.; Lešnik, A.; Šalamun, A. Influence of Landscape Factors on Amphibian Roadkills at the National Level. Diversity 2019, 11, 13. https://doi.org/10.3390/d11010013
Sillero N, Poboljšaj K, Lešnik A, Šalamun A. Influence of Landscape Factors on Amphibian Roadkills at the National Level. Diversity. 2019; 11(1):13. https://doi.org/10.3390/d11010013
Chicago/Turabian StyleSillero, Neftalí, Katja Poboljšaj, Aleksandra Lešnik, and Ali Šalamun. 2019. "Influence of Landscape Factors on Amphibian Roadkills at the National Level" Diversity 11, no. 1: 13. https://doi.org/10.3390/d11010013
APA StyleSillero, N., Poboljšaj, K., Lešnik, A., & Šalamun, A. (2019). Influence of Landscape Factors on Amphibian Roadkills at the National Level. Diversity, 11(1), 13. https://doi.org/10.3390/d11010013