Bacterial Diversity and Community Structure in Typical Plant Rhizosphere



Abstract
:1. Introduction
2. Rhizosphere Bacterial Community
3. Factors Controlling the Distribution and Abundance of Bacterial Community in the Rhizosphere
4. How Plant Growth Stages Affect Bacterial Community Dynamics
5. Plant–Root Interactions in the Rhizosphere
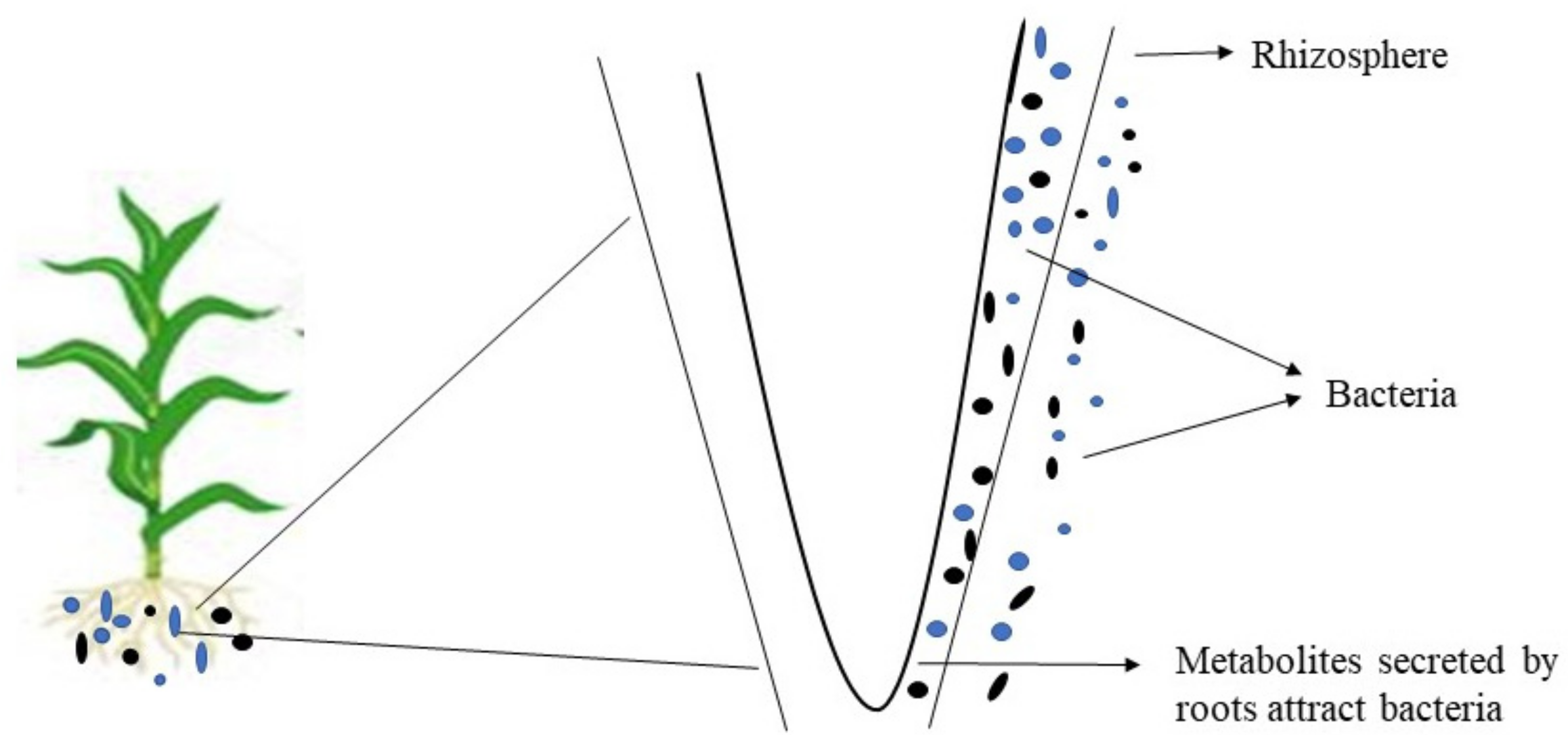
6. Influence of Plant Exudates on Rhizosphere Microbial Dynamics
7. Metabolites
8. Metabolic Fingerprinting
9. The Use of Omics Techniques to Analyze the Rhizosphere
10. Conclusions and Future Outlooks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Babalola, O.O.; Kirby, B.M.; Roes-Hill, L.; Cook, A.E.; Cary, S.C.; Burton, S.G.; Cowan, D.A. Phylogenetic analysis of actinobacterial populations associated with Antarctic Dry Valley mineral soils. Environ. Microbiol. 2009, 11, 566–576. [Google Scholar] [CrossRef]
- Philippot, L.; Raaijmakers, J.; Lemanceau, P.; Van Der Putten, W. Going back to the roots: The microbial ecology of the rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, R.; Gao, J.; Wang, X.; Fan, F.; Ma, X.; Yin, H.; Zhang, C.; Feng, K.; Deng, Y. Thirty-one years of rice-rice-green manure rotations shape the rhizosphere microbial community and enrich beneficial bacteria. Soil Biol. Biochem. 2017, 104, 208–217. [Google Scholar] [CrossRef]
- Fanin, N.; Bertrand, I. Aboveground litter quality is a better predictor than belowground microbial communities when estimating carbon mineralization along a land-use gradient. Soil Biol. Biochem. 2016, 94, 48–60. [Google Scholar] [CrossRef]
- Babalola, O.O. Beneficial bacteria of agricultural importance. Biotechnol. Lett. 2010, 32, 1559–1570. [Google Scholar] [CrossRef]
- Preece, C.; Penuelas, J. Rhizodeposition under drought and consequences for soil communities and ecosystem resilience. Plant Soil 2016, 409, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Lange, M.; Eisenhauer, N.; Sierra, C.; Bessler, H.; Engels, C.; Griffiths, R.; Mellado-Vázquez, P.; Malik, A.; Roy, J.; Scheu, S. Plant diversity increases soil microbial activity and soil carbon storage. Nat. Commun. 2015, 6, 6707. [Google Scholar] [CrossRef]
- Zhang, R.; Vivanco, J.; Shen, Q. The unseen rhizosphere root–soil–microbe interactions for crop production. Curr. Opin. Microbiol. 2017, 37, 8–14. [Google Scholar] [CrossRef]
- Babalola, O.O.; Sanni, A.I.; Odhiambo, G.D.; Torto, B. Plant growth-promoting rhizobacteria do not pose any deleterious effect on cowpea and detectable amounts of ethylene are produced. World J. Microbiol. Biotechnol. 2007, 23, 747–752. [Google Scholar] [CrossRef]
- Enebe, M.; Babalola, O. The influence of plant growth-promoting rhizobacteria in plant tolerance to abiotic stress: A survival strategy. Appl. Microbiol. Biotechnol. 2018, 102, 7821–7835. [Google Scholar] [CrossRef]
- Li, H.; Su, J.; Yang, X.; Zhu, Y. Distinct rhizosphere effect on active and total bacterial communities in paddy soils. Sci. Total Environ. 2019, 649, 422–430. [Google Scholar] [CrossRef]
- Caballero-Mellado, J.; Onofre-Lemus, J.; Estrada-De Los Santos, P.; Martínez-Aguilar, L. The tomato rhizosphere, an environment rich in nitrogen-fixing Burkholderia species with capabilities of interest for agriculture and bioremediation. Appl. Environ. Microbiol. 2007, 73, 5308–5319. [Google Scholar] [CrossRef]
- Petatan-Sagahon, I.; Anducho-Reyes, M.; Silva-Rojas, H.; Arana-Cuenca, A.; Tellez-Jurado, A.; Cárdenas-Álvarez, I.; Mercado-Flores, Y. Isolation of bacteria with antifungal activity against the phytopathogenic fungi Stenocarpella maydis and Stenocarpella macrospora. Int. J. Mol. Sci. 2011, 12, 5522–5537. [Google Scholar] [CrossRef]
- Saubidet, M.; Fatta, N.; Barneix, A. The effect of inoculation with Azospirillum brasilense on growth and nitrogen utilization by wheat plants. Plant Soil 2002, 245, 215–222. [Google Scholar] [CrossRef]
- Yanni, Y.; El-Fattah, F. Towards integrated biofertilization management with free living and associative dinitrogen fixers for enhancing rice performance in the Nile delta. Symbiosis 1999, 27, 319–331. [Google Scholar]
- Muratova, A.; Turkovskaya, O.; Antonyuk, L.; Makarov, O.; Pozdnyakova, L.; Ignatov, V. Oil-oxidizing potential of associative rhizobacteria of the genus Azospirillum. Microbiology 2005, 74, 210–215. [Google Scholar] [CrossRef]
- Åström, B.; Gustafsson, A.; Gerhardson, B. Characteristics of a plant deleterious rhizosphere Pseudomonad and its inhibitory metabolite (s). J. Appl. Bacteriol. 1993, 74, 20–28. [Google Scholar] [CrossRef]
- Murphy, J.; Zehnder, G.; Schuster, D.; Sikora, E.; Polston, J.; Kloepper, J. Plant growth-promoting rhizobacterial mediated protection in tomato against Tomato mottle virus. Plant Dis. 2000, 84, 779–784. [Google Scholar] [CrossRef]
- Carvalho, D.; Oliveira, D.; Corrêa, R.; Campos, V.; Guimarães, R.; Coimbra, J. Rhizobacteria able to produce phytotoxic metabolites. Braz. J. Microbiol. 2007, 38, 759–765. [Google Scholar] [CrossRef] [Green Version]
- Manivanh, L.; Pierret, A.; Rattanavong, S.; Kounnavongsa, O.; Buisson, Y.; Elliott, I.; Maeght, J.; Xayyathip, K.; Silisouk, J.; Vongsouvath, M. Burkholderia pseudomallei in a lowland rice paddy: Seasonal changes and influence of soil depth and physico-chemical properties. Sci. Rep. 2017, 7, 3031. [Google Scholar] [CrossRef]
- Babalola, O.; Akindolire, A. Identification of native rhizobacteria peculiar to selected food crops in Mmabatho municipality of South Africa. Biol. Agric. Hortic. 2011, 27, 294–309. [Google Scholar] [CrossRef]
- Hou, M.P.; Babalola, O.O. Evaluation of plant growth promoting potential of four rhizobacterial species for indigenous system. J. Cent. South Univ. 2013, 20, 164–171. [Google Scholar] [CrossRef]
- Olanrewaju, O.; Glick, B.; Babalola, O. Mechanisms of action of plant growth promoting bacteria. World J. Microbiol. Biotechnol. 2017, 33, 197. [Google Scholar] [CrossRef]
- Adegboye, M.; Babalola, O. Phylogenetic characterization of culturable antibiotic producing Streptomyces from rhizospheric soils. Mol. Biol. S 2013, 1, 001. [Google Scholar]
- Adegboye, M.F.; Babalola, O.O.; Ngoma, L.; Okoh, A.I. Analysis of Streptomyces spp. native to Mahikeng soils in South Africa. J. Pure Appl. Microbiol. 2012, 6, 1001–1010. [Google Scholar]
- Babalola, O.O.; Glick, B.R. The use of microbial inoculants in African agriculture: Current practice and future prospects. J. Food Agric. Environ. 2012, 10, 540–549. [Google Scholar]
- Babalola, O.O. Molecular techniques: An overview of methods for the detection of bacteria. Afr. J. Biotechnol. 2003, 2, 710–713. [Google Scholar] [Green Version]
- Singh, B.K.; Munro, S.; Potts, J.M.; Millard, P. Influence of grass species and soil type on rhizosphere microbial community structure in grassland soils. Appl. Soil Ecol. 2007, 36, 147–155. [Google Scholar]
- Goswami, D.; Thakker, J.; Dhandhukia, P. Portraying mechanics of plant growth promoting rhizobacteria (PGPR): A review. Cogent Food Agric. 2016, 2, 1127500. [Google Scholar] [CrossRef]
- Bumunang, E.W.; Babalola, O.O.; Barros, E. Bacterial community profiling in the rhizosphere of field grown GM and non-GM maize. J. Pure Appl. Microbiol 2013, 7, 1–21. [Google Scholar]
- Beauregard, P.; Chai, Y.; Vlamakis, H.; Losick, R.; Kolter, R. Bacillus subtilis biofilm induction by plant polysaccharides. Proc. Natl. Acad. Sci. USA 2013, 110, E1621–E1630. [Google Scholar] [CrossRef]
- Lahav, I.; Steinberger, Y. Soil bacterial functional diversity in a potato field. Eur. J. Soil Biol. 2001, 37, 59–67. [Google Scholar] [CrossRef]
- Grayston, S.; Griffith, G.; Mawdsley, J.; Campbell, C.; Bardgett, R.D. Accounting for variability in soil microbial communities of temperate upland grassland ecosystems. Soil Biol. Biochem. 2001, 33, 533–551. [Google Scholar] [CrossRef]
- Lejon, D.P.; Sebastia, J.; Lamy, I.; Chaussod, R.; Ranjard, L. Relationships between soil organic status and microbial community density and genetic structure in two agricultural soils submitted to various types of organic management. Microb. Ecol. 2007, 53, 650–663. [Google Scholar] [CrossRef]
- Yadav, B.; Akhtar, M.; Panwar, J. Rhizospheric plant-microbe interactions: Key factors to soil fertility and plant nutrition. In Plant Microbes Symbiosis: Applied Facets; Springer: New Delhi, India, 2015; pp. 127–145. [Google Scholar]
- Schlemper, T.; Leite, M.; Lucheta, A.; Shimels, M.; Bouwmeester, H.; van Veen, J.; Kuramae, E. Rhizobacterial community structure differences among sorghum cultivars in different growth stages and soils. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef] [Green Version]
- Brimecombe, M.J.; Leij, F.A.; Lynch, J.M. Nematode community structure as a sensitive indicator of microbial perturbations induced by a genetically modified Pseudomonas fluorescens strain. Biol. Fertil. Soils 2001, 34, 270–275. [Google Scholar] [CrossRef]
- Zhou, D.; Huang, X.; Chaparro, J.; Badri, D.; Manter, D.; Vivanco, J.; Guo, J. Root and bacterial secretions regulate the interaction between plants and PGPR leading to distinct plant growth promotion effects. Plant Soil 2016, 401, 259–272. [Google Scholar] [CrossRef]
- Marschner, P.; Joergensen, R.; Piepho, H.-P.; Buerkert, A. Legume rotation effects on early growth and rhizosphere microbiology of sorghum in West African soils. Plant Soil 2004, 264, 325–334. [Google Scholar] [CrossRef] [Green Version]
- Reinhold-Hurek, B.; Bünger, W.; Burbano, C.; Sabale, M.; Hurek, T. Roots shaping their microbiome: Global hotspots for microbial activity. Annu. Rev. Phytopathol. 2015, 53, 403–424. [Google Scholar] [CrossRef]
- Marschner, P.; Yang, C.-H.; Lieberei, R.; Crowley, D. Soil and plant specific effects on bacterial community composition in the rhizosphere. Soil Biol. Biochem. 2001, 33, 1437–1445. [Google Scholar] [CrossRef]
- Weisskopf, L.; Abou-Mansour, E.; Fromin, N.; Tomasi, N.; Santelia, D.; Edelkott, I.; Neumann, G.; Aragno, M.; Tabacchi, R.; Martinoia, E. White lupin has developed a complex strategy to limit microbial degradation of secreted citrate required for phosphate acquisition. Plant Cell Environ. 2006, 29, 919–927. [Google Scholar] [CrossRef] [Green Version]
- Rasmann, S.; Turlings, T. Root signals that mediate mutualistic interactions in the rhizosphere. Curr. Opin. Plant Biol. 2016, 32, 62–68. [Google Scholar] [CrossRef] [Green Version]
- Kai, M.; Effmert, U.; Piechulla, B. Bacterial-plant-interactions: Approaches to unravel the biological function of bacterial volatiles in the rhizosphere. Front. Microbiol. 2016, 7, 108. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; Van Themaat, E.; Schulze-Lefert, P. Structure and functions of the bacterial microbiota of plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef]
- Mendes, L.W.; Kuramae, E.E.; Navarrete, A.A.; Van Veen, J.A.; Tsai, S.M. Taxonomical and functional microbial community selection in soybean rhizosphere. ISME J. 2014, 8, 1577–1587. [Google Scholar] [CrossRef]
- Raghavendra, M.; Nayaka, S.; Nuthan, B. Role of rhizosphere microflora in potassium solubilization. In Potassium Solubilizing Microorganisms for Sustainable Agriculture; Springer: New Delhi, India, 2016; pp. 43–59. [Google Scholar]
- Bloemberg, G.; Lugtenberg, B. Molecular basis of plant growth promotion and biocontrol by Rhizobacteria. Curr. Opin. Plant Biol. 2001, 4, 343–350. [Google Scholar] [CrossRef]
- Dimkpa, C.; Svatoš, A.; Dabrowska, P.; Schmidt, A.; Boland, W.; Kothe, E. Involvement of siderophores in the reduction of metal-induced inhibition of auxin synthesis in Streptomyces spp. Chemosphere 2008, 74, 19–25. [Google Scholar] [CrossRef]
- Koumoutsi, A.; Chen, X.; Henne, A.; Liesegang, H.; Hitzeroth, G.; Franke, P.; Vater, J.; Borriss, R. Structural and functional characterization of gene clusters directing nonribosomal synthesis of bioactive cyclic lipopeptides in Bacillus amyloliquefaciens strain FZB42. J. Bacteriol. 2004, 186, 1084–1096. [Google Scholar] [CrossRef]
- Höfte, M.; Bakker, P. Competition for iron and induced systemic resistance by siderophores of plant growth promoting rhizobacteria. In Microbial Siderophores; Springer: Berlin/Heidelberg, Germany, 2007; pp. 121–133. [Google Scholar]
- Chen, X.; Koumoutsi, A.; Scholz, R.; Eisenreich, A.; Schneider, K.; Heinemeyer, I.; Morgenstern, B.; Voss, B.; Hess, W.; Reva, O. Comparative analysis of the complete genome sequence of the plant growth–promoting bacterium Bacillus amyloliquefaciens FZB42. Nat. Biotechnol. 2007, 25, 1007. [Google Scholar] [CrossRef]
- Kalbe, C.; Marten, P.; Berg, G. Strains of the genus Serratia as beneficial rhizobacteria of oilseed rape with antifungal properties. Microbiol. Res. 1996, 151, 433–439. [Google Scholar] [CrossRef]
- Raaijmakers, J.; Vlami, M.; De Souza, J. Antibiotic production by bacterial biocontrol agents. Antonie Van Leeuwenhoek 2002, 81, 537. [Google Scholar] [CrossRef]
- Arora, N.; Kang, S.; Maheshwari, D. Isolation of siderophore-producing strains of Rhizobium meliloti and their biocontrol potential against Macrophomina phaseolina that causes charcoal rot of groundnut. Curr. Sci. Bangalore 2001, 81, 673–677. [Google Scholar]
- Chin-A-Woeng, T.; van den Broek, D.; Lugtenberg, B.; Bloemberg, G. The Pseudomonas chlororaphis PCL1391 sigma regulator psrA represses the production of the antifungal metabolite phenazine-1-carboxamide. Mol. Plant Microbe Interact. 2005, 18, 244–253. [Google Scholar] [CrossRef]
- Boruah, H.; Kumar, B. Plant disease suppression and growth promotion by a Fluorescent Pseudomonas strain. Folia Microbiol. 2002, 47, 137–143. [Google Scholar] [CrossRef]
- Nagarajkumar, M.; Bhaskaran, R.; Velazhahan, R. Involvement of secondary metabolites and extracellular lytic enzymes produced by Pseudomonas fluorescens in inhibition of Rhizoctonia solani, the rice sheath blight pathogen. Microbiol. Res. 2004, 159, 73–81. [Google Scholar] [CrossRef]
- Loper, J.; Gross, H. Genomic analysis of antifungal metabolite production by Pseudomonas fluorescens Pf-5. In New Perspectives and Approaches in Plant Growth-Promoting Rhizobacteria Research; Springer: Dordrecht, The Netherlands, 2007; pp. 265–278. [Google Scholar]
- Park, M.; Kim, Y.; Lee, S.; Kim, I. Identification of an ISR-related metabolite produced by rhizobacterium Klebsiella oxytoca C1036 active against soft-rot disease pathogen in tobacco. Pest Manag. Sci. Former. Pestic. Sci. 2009, 65, 1114–1117. [Google Scholar] [CrossRef]
- Riedlinger, J.; Schrey, S.; Tarkka, M.; Hampp, R.; Kapur, M.; Fiedler, H. Auxofuran, a novel metabolite that stimulates the growth of fly agaric, is produced by the mycorrhiza helper bacterium Streptomyces strain AcH 505. Appl. Environ. Microbiol. 2006, 72, 3550–3557. [Google Scholar] [CrossRef]
- Gealy, D.; Gurusiddaiah, S.; Ogg, A. Isolation and characterization of metabolites from Pseudomonas syringae-strain 3366 and their phytotoxicity against certain weed and crop species. Weed Sci. 1996, 44, 383–392. [Google Scholar] [CrossRef]
- Gealy, D.; Gurusiddaiah, S.; Ogg, A.; Kennedy, A. Metabolites from Pseudomonas fluorescens strain D7 inhibit downy brome (Bromus tectorum) seedling growth. Weed Technol. 1996, 10, 282–287. [Google Scholar] [CrossRef]
- Jousset, A.; Lara, E.; Wall, L.; Valverde, C. Secondary metabolites help biocontrol strain Pseudomonas fluorescens CHA0 to escape protozoan grazing. Appl. Environ. Microbiol. 2006, 72, 7083–7090. [Google Scholar] [CrossRef]
- Kumar, R.; Ayyadurai, N.; Pandiaraja, P.; Reddy, A.; Venkateswarlu, Y.; Prakash, O.; Sakthivel, N. Characterization of antifungal metabolite produced by a new strain Pseudomonas aeruginosa PUPa3 that exhibits broad-spectrum antifungal activity and biofertilizing traits. J. Appl. Microbiol. 2005, 98, 145–154. [Google Scholar] [CrossRef]
- Verma, A.; Kumar, S.; Kumar, G.; Saini, J.; Agrawal, R.; Satlewal, A.; Ansari, M. Rhizosphere metabolite profiling: An opportunity to understand plant-microbe interactions for crop improvement. In Crop Improvement through Microbial Biotechnology; Elsevier: Amsterdam, The Netherlands, 2018; pp. 343–361. [Google Scholar]
- Beneduzi, A.; Ambrosini, A.; Passaglia, L. Plant growth-promoting rhizobacteria (PGPR): Their potential as antagonists and biocontrol agents. Genet. Mol. Biol. 2012, 35, 1044–1051. [Google Scholar] [CrossRef]
- Bais, H.; Park, S.; Weir, T.; Callaway, R.; Vivanco, J. How plants communicate using the underground information superhighway. Trends Plant Sci. 2004, 9, 26–32. [Google Scholar] [CrossRef]
- Santoyo, G.; Orozco-Mosqueda, M.; Govindappa, M. Mechanisms of biocontrol and plant growth-promoting activity in soil bacterial species of Bacillus and Pseudomonas: A review. Biocontrol Sci. Technol. 2012, 22, 855–872. [Google Scholar] [CrossRef]
- Thakur, M.; Bhattacharya, S.; Khosla, P.; Puri, S. Improving production of plant secondary metabolites through biotic and abiotic elicitation. J. Appl. Res. Med. Aromat. Plants 2019, 12, 1–12. [Google Scholar] [CrossRef]
- Cirlini, M.; Dall’Asta, C.; Silvanini, A.; Beghè, D.; Fabbri, A.; Galaverna, G.; Ganino, T. Volatile fingerprinting of chestnut flours from traditional Emilia Romagna (Italy) cultivars. Food Chem. 2012, 134, 662–668. [Google Scholar] [CrossRef]
- Choi, H.; Choi, Y.; Verberne, M.; Lefeber, A.; Erkelens, C.; Verpoorte, R. Metabolic fingerprinting of wild type and transgenic tobacco plants by 1H NMR and multivariate analysis technique. Phytochemistry 2004, 65, 857–864. [Google Scholar] [CrossRef]
- Sachdev, D.; Chaudhari, H.; Kasture, V.; Dhavale, D.; Chopade, B. Isolation and characterization of indole acetic acid (IAA) producing Klebsiella pneumoniae strains from rhizosphere of wheat (Triticum aestivum) and their effect on plant growth. Indian J. Exp. Biol. 2009, 47, 993–1000. [Google Scholar]
- Ndakidemi, P.; Dakora, F. Legume seed flavonoids and nitrogenous metabolites as signals and protectants in early seedling development. Funct. Plant Biol. 2003, 30, 729–745. [Google Scholar] [CrossRef]
- Armero, J.; Requejo, R.; Jorrín, J.; López-Valbuena, R.; Tena, M. Release of phytoalexins and related isoflavonoids from intact chickpea seedlings elicited with reduced glutathione at root level. Plant Physiol. Biochem. 2001, 39, 785–795. [Google Scholar] [CrossRef]
- Friedman, M. Potato glycoalkaloids and metabolites: Roles in the plant and in the diet. J. Agric. Food Chem. 2006, 54, 8655–8681. [Google Scholar] [CrossRef]
- Efange, S.; Brun, R.; Wittlin, S.; Connolly, J.; Hoye, T.; McAkam, T.; Makolo, F.; Mbah, J.; Nelson, D.; Nyongbela, K. Okundoperoxide, a bicyclic cyclofarnesylsesquiterpene endoperoxide from Scleria striatinux with antiplasmodial activity. J. Nat. Prod. 2009, 72, 280–283. [Google Scholar] [CrossRef]
- Kozukue, N.; Han, J.; Lee, K.; Friedman, M. Dehydrotomatine and α-tomatine content in tomato fruits and vegetative plant tissues. J. Agric. Food Chem. 2004, 52, 2079–2083. [Google Scholar] [CrossRef]
- Garcia-Perez, I.; Vallejo, M.; Garcia, A.; Legido-Quigley, C.; Barbas, C. Metabolic fingerprinting with capillary electrophoresis. J. Chromatogr. A 2008, 1204, 130–139. [Google Scholar] [CrossRef]
- Jorge, T.; Rodrigues, J.; Caldana, C.; Schmidt, R.; van Dongen, J.; Thomas-Oates, J.; António, C. Mass spectrometry-based plant metabolomics: Metabolite responses to abiotic stress. Mass Spectrom. Rev. 2016, 35, 620–649. [Google Scholar] [CrossRef]
- White III, R.; Rivas-Ubach, A.; Borkum, M.; Köberl, M.; Bilbao, A.; Colby, S.; Hoyt, D.; Bingol, K.; Kim, Y.; Wendler, J. The state of rhizospheric science in the era of multi-omics: A practical guide to omics technologies. Rhizosphere 2017, 3, 212–221. [Google Scholar] [CrossRef]
- Newman, M.; Lorenz, N.; Hoilett, N.; Lee, N.; Dick, R.; Liles, M.; Ramsier, C.; Kloepper, J. Changes in rhizosphere bacterial gene expression following glyphosate treatment. Sci. Total Environ. 2016, 553, 32–41. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Plant Rhizosphere | Rhizobacteria | Effect | References |
|---|---|---|---|
| Positive effect | |||
| Tomato (Lycopersicon esculentum var. esculentum) | Burkholderia cepacia; B. unamae; B. tropica; B. xenovorans | N2 fixation, plant growth promotion, and bioremediation | [12] |
| Corn (Zea mays) | Bacillus subtilis; Pseudomonas fluorescens; Pantoea agglomerans | Antifungal activity | [13] |
| Wheat (Triticum aestivum) | Azospirillum brasilense | Promotes the uptake of NO3−, K+, and H2PO4− | [14] |
| Rice (Oryza sativa) | Azotobacter vinelandii; Azotobacter chroococcum | Plant growth promotion | [15] |
| Wheat | Azospirillum lipoferum | Promotes the development of the root system of wheat even under the contamination of crude oil | [16] |
| Negative effect | |||
| Wheat | Pseudomonas fluorescens angstrom313 | Plant growth reduction | [17] |
| Tomato | Bacillus subtilis | Causes sour skin, a bacterial soft-rotting disease of onion | [18] |
| Castor oil plant (Ricinus communis) | Bacillus cereus | Causes rootlet rot | [19] |
| Rice paddies | Burkholderia pseudomallei | Causative agents of melioidosis | [20] |
| Rhizobacteria | Metabolites | Function of the Metabolites | References |
|---|---|---|---|
| Pseudomonas spp. | Phenazines, pyrrolnitrin, pyoluteorin, viscosinamide | Viscosinamide found to prevent the infection of sugar beet by Pythium ultimum | [48] |
| Streptomyces spp. | Siderophores | Alleviate metal-contamination stress on plants | [49] |
| Bacillus amyloliquefaciens strain FZB 42 | lipopeptides, surfactins, bacillomycin D, fengycins | Antifungal activity | [50] |
| P. aeruginosa | Pyoverdine, pyochelin, salicylic acid | Induces resistance to plant diseases caused by Botrytis cinerea on bean and tomato, Colletotrichum lindemuthianum | [51] |
| Bacillus amyloliquefaciens | Bacillomycin D, fengycins, surfactins | Enhance plant growth and suppress plant pathogenic organisms | [52] |
| Serratia spp. | Siderophores, pyrrolnitrin, prodigiosin | Antifungal activity against different phytopathognic fungi Verticillium dahlia, Rhizoctonia solani, Sclerotinia sclerotiorum | [53] |
| P. fluorescens | Pyoluteorin, pyrrolnitrin, phenazine-1-carboxylic acid, 2,4-diacetylphloroglucinol | Suppress soil borne plant pathogens, including fungi and nematodes | [54] |
| Rhizobium meliloti | Siderophores | Antifungal activity against Macrophomina phaseolina | [55] |
| P. chlorophis PCL1391 | Phenazine-1-carboxamide | Antifungal | [56] |
| P. fluorescens | Siderophore pyoverdin | Antibacterial and antifungal activity. Enhanced plant growth | [57] |
| P. fluorescens PfMDUS | Chitinase, beta-1,3-glucanase, siderophores, salicylic acid, hydrogen cyanide | Inhibit mycelial growth of R. solani | [58] |
| P. fluorescens Pf-5 | Antibiotics, siderophores | Antibiotics toxic to soil-borne fungi and oomycetes that infect plant roots/siderophores involved in iron acquisition | [59] |
| Klebsiella oxytoca C1036 | Butyl 2-pyrolidone-5-carboxylate | Active against soft-rot disease pathogen in tobacco | [60] |
| Streptomyces AcH 505 | Auxofuran | Stimulates the growth of fly agaric, suppresses growth of ectomycorrhizal fungi | [61] |
| P. syringae strain 366 | Phenazine-1-carboxylic acid, 2-amino phenoxazone, 2-amino phenol | Inhibit downy brome root growth | [62] |
| P. fluorescens D7 | Uncharacterized phytotoxins | Inhibit downy brome root growth | [63] |
| P. fluorescens CHAO | Antibiotics, hydrogen cyanide, and an exoprotease | Protists’ growth inhibition, encystation, paralysis, and cell lysis | [64] |
| P. aeruginosa PUPa3 | Phenazine-1-carboxamide | Broad-spectrum antifungal activity and biofertilizing traits | [65] |
| Plant | Metabolites | Function of the Metabolites | References |
|---|---|---|---|
| Sweet chestnut (Castanea sativa Mill.) | ϒ-butyrolactone, ϒ-terpinene, furfural, benzaldehyde, 4-methyl-2-pentanone | Peculiar aroma, the most typical organoleptic characteristic of chestnut-based products | [71] |
| Tobacco (Nicotiana tabacum) | Salicylic acid, alkaloids, flavonoids | Plays a role in plant growth and development, photosynthesis, ion uptake, and transport. Resistant to pathogens by inducing production of pathogenesis-related proteins. UV filtration and symbiotic nitrogen fixation | [72] |
| Wheat (Triticum spp.) | Tryptophan | For the biosynthesis of indole-3-acetic acid by associative bacteria | [73] |
| Legume seeds | Flavonoids, alkaloids, terpenoids, peptides, amino acids | Serve as eco-sensing signals for suitable rhizobacteria and arbuscular mycorrhizal fungal towards symbiotic mutualisms. Defense molecules against pathogens and insect pests. | [74] |
| Chickpea (Cicer arietinum) | Biochanin A, medicarpin, formononetin, maeckiain | Not stated | [75] |
| Potato (Solanum tuberosum) | Glycoalkaloids, calystegine, lectins | They protect plants against phytopathogens | [76] |
| Spice (Scleria stiatinux) | Okundoperoxide | It contains antiviral and antifungal properties, also used as herbal tea for fevers | [77] |
| Tomato (Solanum lycopersicum) | α-tomatine, dehydrotomatine | Defend plants against attack by microorganisms and herbivores due to their insecticidal activity and have allelopathic effects on many weeds. | [78] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alawiye, T.T.; Babalola, O.O. Bacterial Diversity and Community Structure in Typical Plant Rhizosphere. Diversity 2019, 11, 179. https://doi.org/10.3390/d11100179
Alawiye TT, Babalola OO. Bacterial Diversity and Community Structure in Typical Plant Rhizosphere. Diversity. 2019; 11(10):179. https://doi.org/10.3390/d11100179
Chicago/Turabian StyleAlawiye, Temitayo Tosin, and Olubukola Oluranti Babalola. 2019. "Bacterial Diversity and Community Structure in Typical Plant Rhizosphere" Diversity 11, no. 10: 179. https://doi.org/10.3390/d11100179
APA StyleAlawiye, T. T., & Babalola, O. O. (2019). Bacterial Diversity and Community Structure in Typical Plant Rhizosphere. Diversity, 11(10), 179. https://doi.org/10.3390/d11100179