The Western Amazonian Richness Gradient for Squamate Reptiles: Are There Really Fewer Snakes and Lizards in Southwestern Amazonian Lowlands?



Abstract
:1. Introduction
2. Materials and Methods
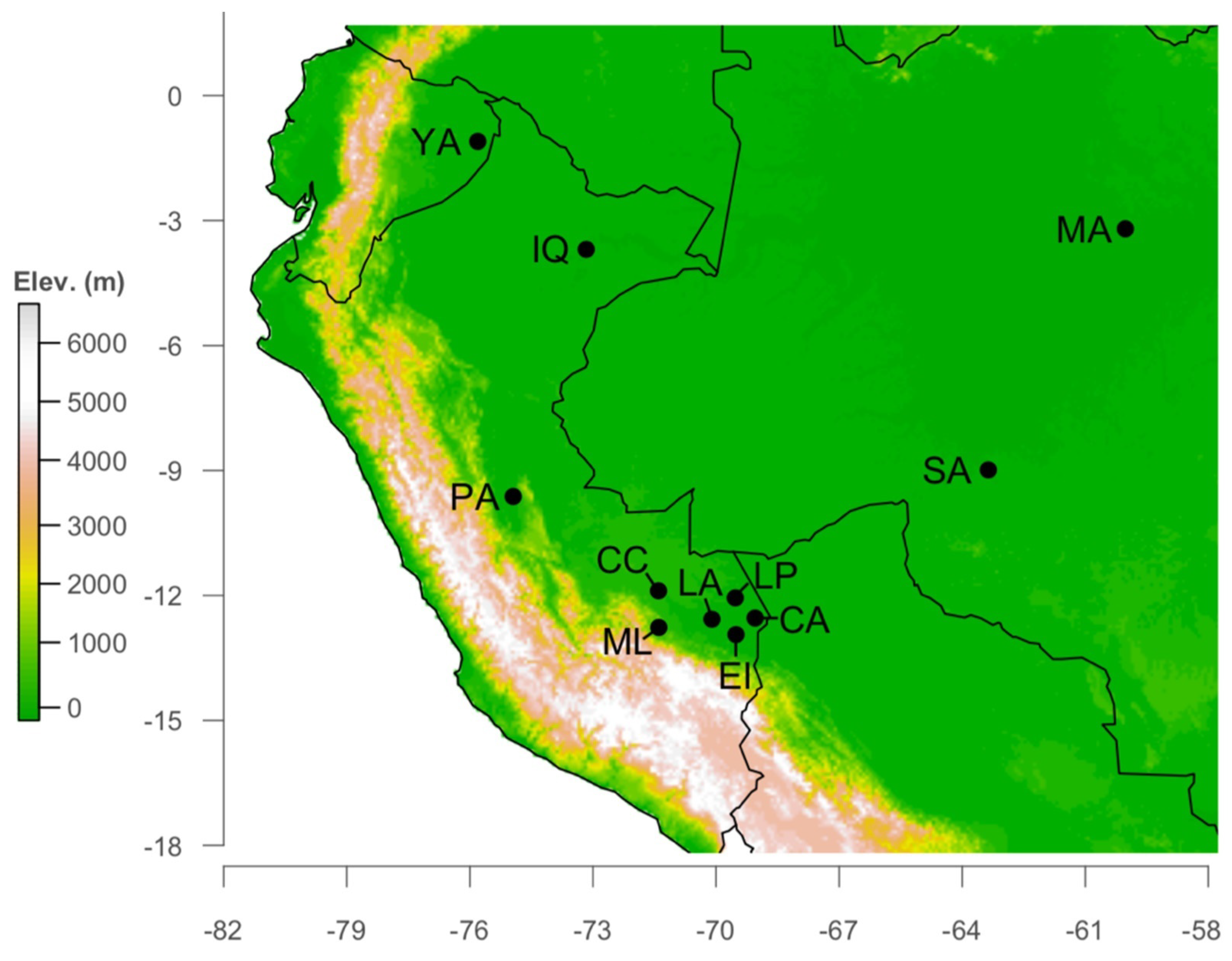
2.1. Study Site
2.2. Data Collection
2.3. Data Tabulation and Analysis
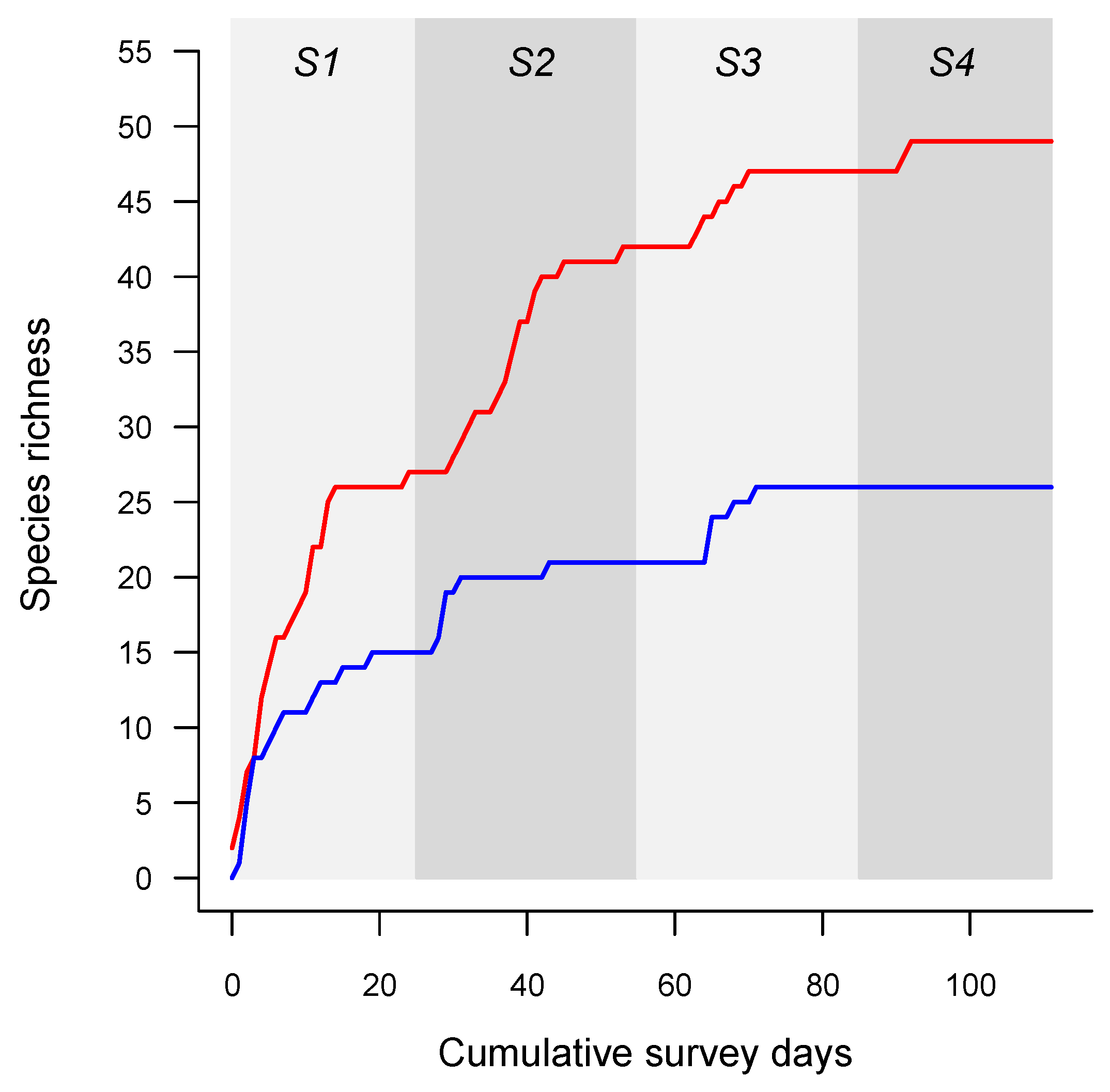
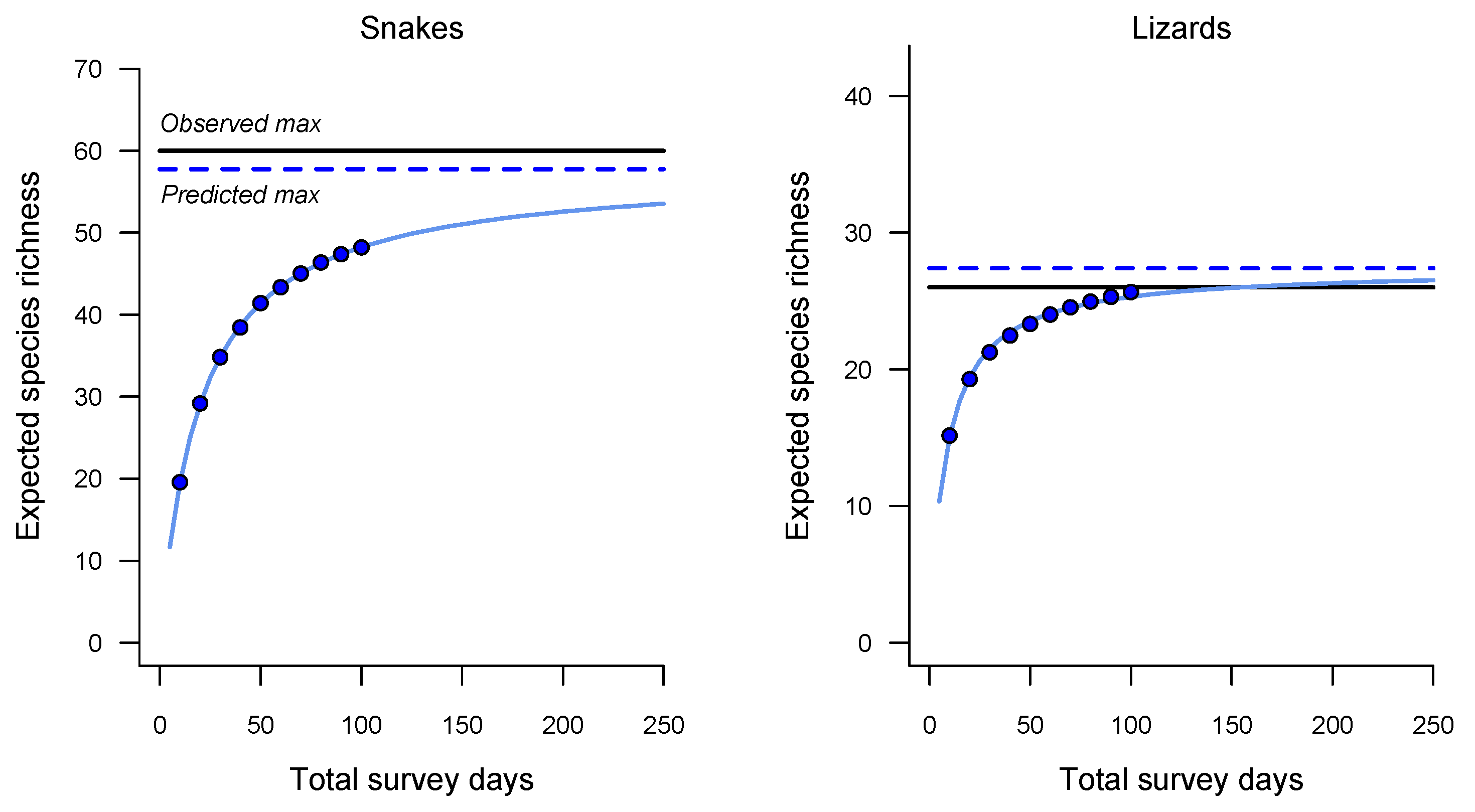
3. Results
4. Discussion
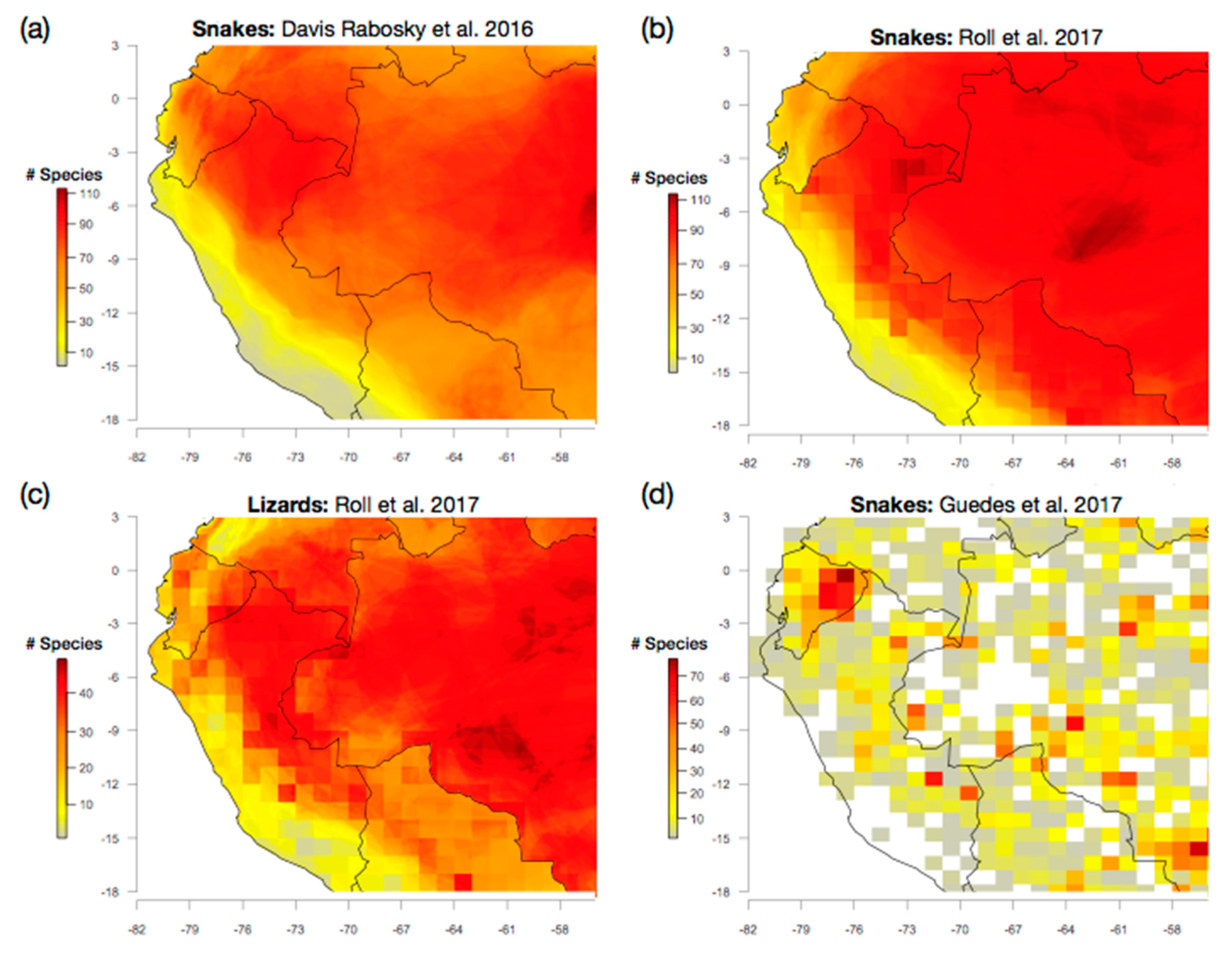
4.1. A Western Amazonian Richness Gradient: Does It Exist?
4.2. Taxonomic Issues
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clade | Species | LABS | LABS-R | LABS-X | Material Examined |
|---|---|---|---|---|---|
| Amphisbaenidae (lizard) | Amphisbaena alba | X 1 | |||
| Amphisbaena fuliginosa | X | X | X | ||
| Anguidae (lizard) | Diploglossus fasciatus | X 2 | |||
| Gekkota (lizard) | Gonatodes hasemani | X | X | X | RAB 2799, MUSM 38982 |
| Gonatodes humeralis | X | X | X | RAB 3434, MUSM 38984 | |
| Pseudogonatodes guianensis | X | X | X | RAB 2508, MUSM 39869 | |
| Thecadactylus solimoensis | X | X | X | RAB 3277, MUSM 39869 | |
| Scincidae (lizard) | Copeoglossum nigropunctatum | X | X | ||
| Exila nigropalmata | X | X | X 3 | RAB 3514, MUSM 39156 | |
| Varzea altamazonica | X | X | |||
| Teioidea (lizard) | Alopoglossus angulatus | X | X | X | UMMZ 246710, MUSM 36813 |
| Ameiva ameiva | X | X | X | RAB 2924, MUSM 39781 | |
| Bachia dorbignyi | X | X | X | RAB 3497, MUSM 39788 | |
| Bachia trisanale | X | X | X3 | RAB 3469, MUSM 38919 | |
| Cercosaura argulus | X | X | X | RAB 2701, MUSM 38930 | |
| Cercosaura bassleri | X | X | X | UMMZ 245038, MUSM 38937 | |
| Cercosaura eigenmanni | X | X | X | UMMZ 246735, MUSM 36933 | |
| Dracaena guianensis | X 1 | ||||
| Iphisa elegans | X | X | X 3 | RAB 2931, MUSM 39020 | |
| Kentropyx altamazonica | X 1,2 | ||||
| Kentropyx pelviceps | X | X | X | UMMZ 246751, MUSM 37269 | |
| Potamites ecpleopus | X 1,2 | ||||
| Ptychoglossus brevifrontalis | X | X | X | UMMZ 246762, MUSM 39120 | |
| Tupinambis teguixin | X | X | X | RAB 3430 | |
| Iguania (lizard) | Anolis fuscoauratus | X | X | X | RAB 2868, MUSM 36859 |
| Anolis ortonii | X | X | X | UMMZ 248371 | |
| Anolis punctatus | X | X | X | RAB 2537, MUSM 39782 | |
| Anolis tandai | X | X | X | RAB 2660, MUSM 38909 | |
| Enyalioides palpebralis | X | X | X | MUSM 38981 | |
| Plica plica | X | X | X | RAB 2704, MUSM 39860 | |
| Plica umbra | X | X | X | RAB 2682, MUSM 39099 | |
| Polychrus liogaster | X 1 | ||||
| Stenocercus fimbriatus | X 1,2 | ||||
| Stenocercus roseiventris | X 1,2 | ||||
| Uracentron azureum | X | X | X | ||
| Uracentron flaviceps | X | X | X | MUSM 39154 | |
| Typhlopidae (snake) | Amerotyphlops reticulatus | X | X | X | UMMZ 248453, MUSM 36160 |
| Leptotyphlopidae (snake) | Epictia diaplocia | X | X | X | RAB 2932, MUSM 39046 |
| Aniliidae (snake) | Anilius scytale | X | X | X 3 | RAB 2499, MUSM 36848 |
| Boidae (snake) | Boa constrictor | X | X | X | |
| Corallus batesii | X | X | X | ||
| Corallus hortulanus | X | X | X | UMMZ 248448, MUSM 36965 | |
| Epicrates cenchria | X | X | X | RAB 2903, MUSM 37120 | |
| Eunectes murinus | X | X | X | RAB 3417 | |
| Colubrinae (snake) | Chironius carinatus / exoletus | X | X | X 3 | UMMZ 246087, MUSM 38943 |
| Chironius fuscus | X | X | X | UMMZ 245047, MUSM 36957 | |
| Chironius multiventris | X | X | X | UMMZ 245049, MUSM 36962 | |
| Chironius scurrulus | X 1,7 | ||||
| Chironius sp. | X | X | X 3,7 | UMMZ 248360 | |
| Dendrophidion dendrophis | X | X | X | MUSM 36979 | |
| Drymarchon corais | X | X | X 3 | RAB 2739, MUSM 36145 | |
| Drymobius rhombifer | X | X | X 3 | UMMZ 245052, MUSM 39824 | |
| Drymoluber dichrous | X | X | X | UMMZ 246799, MUSM 38977 | |
| Leptophis ahaetulla | X | X | X | UMMZ 246823, MUSM 37345 | |
| Oxybelis aeneus | X 1 | ||||
| Oxybelis fulgidus | X 1 | ||||
| Phrynonax poecilonotus | X | X | X 3 | RAB 3532, MUSM 39115 | |
| Rhinobothryum lentiginosum | X | X | X | ||
| Spilotes pullatus | X | X | X | ||
| Spilotes sulphureus | X 1 | ||||
| Tantilla melanocephala | X | X | X 3 | UMMZ 246845, MUSM 39145 | |
| Dipsadinae (snake) | Apostolepis nigroterminata | X | X | X 3 | RAB 3393, MUSM 39783 |
| Atractus elaps | X | X | X 3 | RAB 3414, MUSM 39784 | |
| Atractus major | X 1 | ||||
| Atractus snethlageae | X | X | X 4 | RAB 2666, MUSM 35703 | |
| Atractus sp. | X | X | X 3 | RAB 2924, MUSM 38913 | |
| Clelia clelia | X | X | X | ||
| Dipsas catesbyi | X | X | X | RAB 2802, MUSM 38970 | |
| Dipsas indica | X | X | X 3 | RAB 3561, MUSM 38973 | |
| Drepanoides anomalus | X | X | X | RAB 3408, MUSM 38974 | |
| Erythrolampus dorsocorallinus6 | X | X | X 3 | MUSM 36049 | |
| Erythrolampus oligolepis6 | X | X | X 3 | RAB 3411, MUSM 39049 | |
| Erythrolampus reginae6 | X | X | X | RAB 3436, MUSM 39837 | |
| Erythrolampus sp. | X | X | X | RAB 2711 | |
| Erythrolampus taeniogaster6 | X | X | X | ||
| Helicops angulatus | X | X | X | UMMZ 245053, MUSM 38989 | |
| Helicops leopardinus | X | X | X | ||
| Helicops polylepis | X | X | |||
| Hydrops triangularis | X | X | X 3 | RAB 3500, MUSM 38992 | |
| Imantodes cenchoa | X | X | X | UMMZ 246814, MUSM 35857 | |
| Imantodes lentiferus | X | X | X | RAB 2900, MUSM 39834 | |
| Leptodeira annulata | X | X | X | UMMZ 246822, MUSM 39043 | |
| Oxyrhopus formosus | X | X | X | UMMZ 248365, MUSM 39077 | |
| Oxyrhopus melanogenys | X | X | X | UMMZ 246832, MUSM 39855 | |
| Oxyrhopus petolarius | X | X | X | UMMZ 245072, MUSM 39081 | |
| Philodryas argentea | X | ||||
| Philodryas viridissima | X | X | |||
| Pseudoboa coronata | X | ||||
| Pseudoeryx plicatilis | X | X | X | MUSM 24359 | |
| Siphlophis cervinus | X | X | X 3 | MUSM 39141 | |
| Siphlophis compressus | X | X | X | MUSM 39142 | |
| Taeniophallus brevirostris | X | X | X 3 | RAB 2462, MUSM 39143 | |
| Taeniophallus occipitalis | X | X | X | RAB 3391, MUSM 37708 | |
| Xenodon rhabdocephalus | X | X | X 3 | UMMZ 246850, MUSM 39158 | |
| Xenodon severus | X | X | X | RAB 2712 | |
| Xenopholis scalaris | X | X | X | UMMZ 245077, MUSM 39889 | |
| Elapidae (snake) | Micrurus annellatus | X | X | X | UMMZ 248451, MUSM 39056 |
| Micrurus hemprichii | X | X | X 3,5 | ||
| Micrurus lemniscatus | X | X | X | UMMZ 248456, MUSM 39057 | |
| Micrurus obscurus | X | X | X | UMMZ 248458, MUSM 39846 | |
| Micrurus surinamensis | X | X | X 3 | UMMZ 246861, MUSM 37353 | |
| Viperidae (snake) | Bothrops atrox | X | X | X | RAB 2814, MUSM 35721 |
| Bothrops bilineatus | X | X | X | UMMZ 246865, MUSM 36919 | |
| Bothrops brazili | X | X | X | MUSM 36922 | |
| Lachesis muta | X | X | X | UMMZ 248454, MUSM 36149 |
References
- Rosenzweig, M.L. Species Diversity in Space and Time; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Couvreur, T.L.P. Odd man out: Why are there fewer plant species in African rain forests? Plant Syst. Evol. 2015, 301, 1299–1313. [Google Scholar] [CrossRef]
- Pianka, E.R. Zoogeography andspeciation of Australian desert lizards. Copeia 1972, 127–145. [Google Scholar] [CrossRef]
- Pianka, E.R. Ecology and Natural History of Desert Lizards; Princeton University Press: Princeton, NJ, USA, 1986. [Google Scholar]
- Morton, S.R.; James, C.D. The diversity and abundance of lizards in arid Australia—A new hypothesis. Am. Nat. 1988, 132, 237–256. [Google Scholar] [CrossRef]
- Rabosky, D.L.; Cowan, M.A.; Talaba, A.L.; Lovette, I.J. Species interactions mediate phylogenetic community structure in a hyperdiverse lizard assemblage from arid Australia. Am. Nat. 2011, 178, 579–595. [Google Scholar] [CrossRef] [PubMed]
- Bass, M.S.; Finer, M.; Jenkins, C.N.; Kreft, H.; Cisneros-Heredia, D.F.; McCracken, S.F.; Pitman, N.C.A.; English, P.H.; Swing, K.; Villa, G.; et al. Global Conservation Significance of Ecuador’s Yasuni National Park. PLoS ONE 2010, 5, e8767. [Google Scholar] [CrossRef]
- Jenkins, C.N.; Pimm, S.L.; Joppa, L.N. Global patterns of terrestrial vertebrate diversity and conservation. Proc. Natl. Acad. Sci. USA 2013, 110, E2602–E2610. [Google Scholar] [CrossRef] [PubMed]
- Pitman, N.C.A.; Terborgh, J.W.; Silman, M.R.; Nunez, P.; Neill, D.A.; Ceron, C.E.; Palacios, W.A.; Aulestia, M. A comparison of tree species diversity in two upper Amazonian forests. Ecology 2002, 83, 3210–3224. [Google Scholar] [CrossRef]
- Magnusson, W.E. Snakes and Other Lizards; Open Science Publishers: New York, NY, USA, 2019. [Google Scholar]
- Da Silva, N.J.; Sites, J.W. Patterns of diversity of neotropical squamate reptile species with emphasis on the brazilian amazon and the conservation potential of indigenous reserves. Conserv. Biol. 1995, 9, 873–901. [Google Scholar] [CrossRef]
- Guedes, T.B.; Sawaya, R.J.; Zizka, A.; Laffan, S.; Faurby, S.; Pyron, R.A.; Bernils, R.S.; Jansen, M.; Passos, P.; Prudente, A.L.C.; et al. Patterns, biases and prospects in the distribution and diversity of Neotropical snakes. Glob. Ecol. Biogeogr. 2018, 27, 14–21. [Google Scholar] [CrossRef]
- Rabosky, A.R.D.; Cox, C.L.; Rabosky, D.L.; Title, P.O.; Holmes, I.A.; Feldman, A.; McGuire, J.A. Coral snakes predict the evolution of mimicry across New World snakes. Nat. Commun. 2016, 7, 11484. [Google Scholar] [CrossRef]
- Roll, U.; Feldman, A.; Novosolov, M.; Allison, A.; Bauer, A.M.; Bernard, R.; Bohm, M.; Castro-Herrera, F.; Chirio, L.; Collen, B.; et al. The global distribution of tetrapods reveals a need for targeted reptile conservation. Nat. Ecol. Evol. 2017, 1, 1677–1682. [Google Scholar] [CrossRef] [PubMed]
- Duellman, W. Cusco Amazonico; Comstock Publishing: Ithaca, NY, USA, 2005. [Google Scholar]
- Whitworth, A.; Downie, R.; von May, R.; Villacampa, J.; MacLeod, R. How much potential biodiversity and conservation value can a regenerating rainforest provide? A ‘best-case’ approach from the Peruvian Amazon. Trop. Conserv. Sci. 2016, 9, 224–245. [Google Scholar] [CrossRef]
- De Fraga, R.; Stow, A.J.; Magnusson, W.E.; Lima, A.P. The Costs of Evaluating Species Densities and Composition of Snakes to Assess Development Impacts in Amazonia. PLoS ONE 2014, 9, e105453. [Google Scholar] [CrossRef] [PubMed]
- Greene, H.W. Snakes: The Evolution of Mystery in Nature; University of California Press: Berkeley, CA, USA, 1997. [Google Scholar]
- Catenazzi, A.; Lehr, E.; von May, R. The amphibians and reptiles of Manu National Park and its buffer zone, Amazon basin and eastern slopes of the Andes, Peru. Biota Neotrop. 2013, 13, 269–283. [Google Scholar] [CrossRef] [Green Version]
- Von May, R.; Donnelly, M.A. Do trails affect relative abundance estimates of rainforest frogs and lizards? Austral Ecol. 2009, 34, 613–620. [Google Scholar] [CrossRef]
- Von May, R.; Jacobs, J.M.; Santa-Cruz, R.; Valdivia, J.; Huaman, J.M.; Donnelly, M.A. Amphibian community structure as a function of forest type in Amazonian Peru. J. Trop. Ecol. 2010, 26, 509–519. [Google Scholar] [CrossRef]
- Pitman, N.C.A. An Overview of the Los Amigos Watershed, Madre de Dios, Southeastern Peru; (Unpublished Report); Amazon Conservation Association: Washington, DC, USA, 2010. [Google Scholar]
- Silman, M.R.; Terborgh, J.W.; Kiltie, R.A. Population regulation of a dominant-rain forest tree by a major seed-predator. Ecology 2003, 84, 431–438. [Google Scholar] [CrossRef]
- Griscom, B.W.; Ashton, P.M.S. A self-perpetuating bamboo disturbance cycle in a neotropical forest. J. Trop. Ecol. 2006, 22, 587–597. [Google Scholar] [CrossRef]
- Olivier, J. Bambusiform grasses (Poaceae) from the Los Amigos River, Madre de Dios, Peru. Rev. Peru. Biol. 2008, 15, 121–126. [Google Scholar]
- Thompson, G.G.; Thompson, S.A. Comparative temperature in funnel and pit traps. Aust. J. Zool. 2009, 57, 311–316. [Google Scholar] [CrossRef]
- Ascenso, A.C.; Costa, J.C.L.; Prudente, A.L.C. Taxonomic revision of the Erythrolamprus reginae species group, with description of a new species from Guiana Shield (Serpentes: Xenodontinae). Zootaxa 2019, 4586, 65–97. [Google Scholar] [CrossRef]
- Colwell, R.K.; Coddington, J.A. Estimating terrestrial biodiversity through extrapolation. Philos. Trans. R. Soc. B 1994, 345, 101–118. [Google Scholar] [Green Version]
- De Oliveira, L.; Araujo, I.; Prudente, A.L.C.; de Fraga, R.; de Almeida, A.; Ascenso, A.C. New distributional records of the toad-headed pitviper Bothrocophias hyoprora (Amaral, 1935) in Brazil. Amphib. Reptile Conserv. 2018, 12, 1–4. [Google Scholar]
- Colwell, R.K.; Winkler, D.W. A null model for null models in biogeography. In Ecological Communities: Conceptual Issues and the Evidence; Strong, D.R., Simberloff, D., Abele, L.G., Thistle, A.B., Eds.; Princeton University Press: Princeton, NJ, USA, 1984; pp. 344–359. [Google Scholar]
- Dixon, J.R.; Soini, P. The Reptiles of the Upper Amazon Basin, Iquitos Region, Peru I–VII; Milwaukee Public Museum: Milwaukee, WI, USA, 1986. [Google Scholar]
- Doan, T.M.; Arriaga, W. Microgeographic variation in species composition of the herpetofaunal communities of Tambopata Region, Peru. Biotropica 2002, 34, 101–117. [Google Scholar] [CrossRef]
- Schluter, A.; Icochea, J.; Perez, J.M. Amphibians and reptiles of the lower Rio Llullapichis, Amazonian Peru: Updated species list with ecological and biogeographic notes. Salamandra 2004, 40, 141–146. [Google Scholar]
- Francisco, B.C.S.; Pinto, R.R.; Fernandes, D.S. Taxonomy of Epictia munoai (Orejas-Miranda, 1961) (Squamata: Serpentes: Leptotyphlopidae). Zootaxa 2012, 3512, 42–52. [Google Scholar] [CrossRef]
- Kreft, H.; Koster, N.; Kuper, W.; Nieder, J.; Barthlott, W. Diversity and biogeography of vascular epiphytes in western Amazonia, Yasuní, Ecuador. J. Biogeogr. 2004, 31, 1463–1476. [Google Scholar] [CrossRef]
- Aceby, A.; Kromer, T. Diversidad y distribución vertical de epífitas en los alrededores del campamento Río Eslabón y de la laguna Chalalán, Parque Nacional Madidi, Dpto. La Paz, Bolivia. Rev. Soc. Boliv. Botánica 2001, 3, 104–123. [Google Scholar]
- Burnham, R.J.; Romero-Saltos, H.G. Diversity and distribution of lianas in Yasuní, Ecuador. In Ecology of Lianas; Schnitzer, S.A., Bongers, F., Burnham, R.J., Putz, F.E., Eds.; Wiley: West Sussex, UK, 2015; pp. 50–64. [Google Scholar]
- Karr, J.R.; Robinson, S.K.; Blake, J.G.; Bierregaard, R.O.J. Birds of four neotropical forests. In Four Neotropical Rainforests; Gentry, A.H., Ed.; Yale University Press: New Haven, CT, USA, 1990. [Google Scholar]
- Sullivan, B.L.; Wood, C.L.; Ilif, M.J.; Bonney, R.E.; Fink, D.; Kelling, S. eBird: A citizen-based bird observation network in the biological sciences. Biol. Conserv. 2009, 142, 2288–2292. [Google Scholar] [CrossRef]
- Janson, C.H.; Emmons, L.H. Ecological structure of the nonflying mammal community at Cocha Cashu Biological Station, Manu National Park, Peru. In Four Neotropical Rainforests; Gentry, A.H., Ed.; Yale University Press: New Haven, CT, USA, 1990; pp. 314–338. [Google Scholar]
- Peres, C.A. Primate community structure at twenty western Amazonian flooded and unflooded forests. J. Trop. Ecol. 1997, 13, 381–405. [Google Scholar] [CrossRef]
- Pitman, R.L. Checklist de los Mamíferos no Voladores de Cocha Cashu y Pakitza. 2008. Available online: https://cochacashu.sandiegozooglobal.org/species-lists/ (accessed on 5 September 2019).
- Hoorn, C.; Wesselingh, F.P.; ter Steege, H.; Bermudez, M.A.; Mora, A.; Sevink, J.; Sanmartin, I.; Sanchez-Meseguer, A.; Anderson, C.L.; Figueiredo, J.P.; et al. Amazonia through time: Andean uplift, climate change, landscape evolution, and biodiversity. Science 2010, 330, 927–931. [Google Scholar] [CrossRef] [PubMed]
- Rull, V. Neotropical biodiversity: Timing and potential drivers. Trends Ecol. Evol. 2011, 26, 508–513. [Google Scholar] [CrossRef] [PubMed]
- De Souza, J.R.D.; Venancio, N.M.; de freitas, M.A.; de Souza, M.B.; Verissimo, D. First record of Bothrops taeniatus Wagler, 1824 (Reptilia, Viperidae) for the state of Acre, Brazil. Check List 2013, 9, 430–431. [Google Scholar] [CrossRef]
- Avila-Pires, T.C.S.; Vitt, L.J.; Sartorius, S.S.; Zani, P.A. Squamata (Reptilia) from four sites in southern Amazonia, with a biogeographic analysis of Amazonian lizards. Bol. Mus. Para. Emílio Goeldi. Cienc. Nat. Belém 2009, 4, 99–118. [Google Scholar]
- Da Silva, N. The snakes from Samuel hydroelectric power plant and vicinity, Rondonia, Brazil. Herpetol. Nat. Hist. 1993, 1, 37–86. [Google Scholar]
- Hamdan, B.; Pereira, A.G.; Loss-Oliveira, L.; Rodder, D.; Schrago, C.G. Evolutionary analysis of Chironius snakes unveils cryptic diversity and provides clues to diversification in the Neotropics. Mol. Phylogenet. Evol. 2017, 116, 108–119. [Google Scholar] [CrossRef]
- Nunes, P.M.S.; Fouquet, A.; Curcio, F.F.; Kok, P.J.R.; Rodrigues, M.T. Cryptic species in Iphisa elegans Gray, 1851 (Squamata: Gymnophthalmidae) revealed by hemipenial morphology and molecular data. Zool. J. Linn. Soc. 2012, 166, 361–376. [Google Scholar] [CrossRef]
- Prudente, A.L.C.; Passos, P. New Cryptic Species of Atractus (Serpentes: Dipsadidae) from Brazilian Amazonia. Copeia 2010, 010, 397–404. [Google Scholar] [CrossRef]
- Sturaro, M.J.; Avila-Pires, T.C.S.; Rodrigues, M.T. Molecular phylogenetic diversity in the widespread lizard Cercosaura ocellata (Reptilia: Gymnophthalmidae) in South America. Syst. Biodivers. 2017, 15, 532–540. [Google Scholar] [CrossRef]
- Torres-Carvajal, O.; Echevarria, L.Y.; Lobos, S.E.; Venegas, P.J.; Kok, P.J.R. Phylogeny, diversity and biogeography of Neotropical sipo snakes (Serpentes: Colubrinae: Chironius). Mol. Phylogenet. Evol. 2019, 130, 315–329. [Google Scholar] [CrossRef]
- Meneses-Pelayo, E.; Passos, P. New Polychromatic Species of Atractus (Serpentes: Dipsadidae) from the Eastern Portion of the Colombian Andes. Copeia 2019, 107, 250–261. [Google Scholar] [CrossRef]
- Passos, P.; Scanferla, A.; Melo-Sampaio, P.R.; Brito, J.; Almendariz, A. A giant on the ground: Another large-bodied Atractus (Serpentes: Dipsadinae) from Ecuadorian Andes, with comments on the dietary specializations of the goo-eaters snakes. An. Acad. Bras. Cienc. 2019, 91. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.; Oliveira, M.E. Natural history of snakes in forests of the Manaus region, central Amazonia, Brazil. Herpetol. Nat. Hist. 1999, 6, 78–150. [Google Scholar]





| Locality | Scale | Latitude | Precipitation | Snakes | Lizards | Reference |
|---|---|---|---|---|---|---|
| Manu Learning Center (MLC) 1 | local | −12.8 | 3000 | 40 | 21 | [16] |
| Los Amigos: LABS | local | −12.6 | 2600 | 60 | 27 | This paper |
| Los Amigos: LABS-R | regional | −12.6 | 2600 | 62 | 28 | This paper |
| Los Amigos: LABS-X | regional | −12.6 | 2600 | 68 | 36 | This paper |
| Cusco Amazonico (CA) | local | −12.5 | 2400 | 52 | 28 | [15,32] |
| Tambopata (EI) | local | −12.8 | 2500 | 50 | 26 | [32] |
| Cocha Cashu | local | −11.9 | 2500 | 32 | 25 | [16] |
| Panguana | local | −9.6 | 2500 | 44 | 27 | [33] |
| Yasuni 2 | regional | −0.6 | 3100 | 78 | 33 | [7] |
| Tiputini | local | −0.6 | 3100 | 63 | 31 | [7] |
| Iquitos 3 | regional | −3.7 | 2900 | 89 | 39 | [11] |
| Taxon | Richness (North) | Richness (South) | Diversity Inflation (%) | Scale | Reference |
|---|---|---|---|---|---|
| Trees | 239 | 174 | 40 | local | [9] |
| Trees | 1017 | 693 | 50 | regional 1 | [9] |
| Trees | 1356 | 1004 | 40 | regional 2 | [9] |
| Vascular epiphytes 3 | 313 | 152 | 110 | regional | [35,36] |
| Lianas | 203 | 115 | 80 | local | [37]; Burnham, pers comm |
| Birds | 550–600 | 550–600 | 0 | regional | [7,38,39] |
| Snakes | 90 | 68 | 30 | regional | This study |
| Lizards | 39 | 36 | 10 | regional | This study |
| Frogs | 141 | 100 | 40 | regional | [7,32] |
| Primates | 12–14 | 12–14 | 0 | regional | [7,40,41,42] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rabosky, D.L.; von May, R.; Grundler, M.C.; Davis Rabosky, A.R. The Western Amazonian Richness Gradient for Squamate Reptiles: Are There Really Fewer Snakes and Lizards in Southwestern Amazonian Lowlands? Diversity 2019, 11, 199. https://doi.org/10.3390/d11100199
Rabosky DL, von May R, Grundler MC, Davis Rabosky AR. The Western Amazonian Richness Gradient for Squamate Reptiles: Are There Really Fewer Snakes and Lizards in Southwestern Amazonian Lowlands? Diversity. 2019; 11(10):199. https://doi.org/10.3390/d11100199
Chicago/Turabian StyleRabosky, Daniel L., Rudolf von May, Michael C. Grundler, and Alison R. Davis Rabosky. 2019. "The Western Amazonian Richness Gradient for Squamate Reptiles: Are There Really Fewer Snakes and Lizards in Southwestern Amazonian Lowlands?" Diversity 11, no. 10: 199. https://doi.org/10.3390/d11100199
APA StyleRabosky, D. L., von May, R., Grundler, M. C., & Davis Rabosky, A. R. (2019). The Western Amazonian Richness Gradient for Squamate Reptiles: Are There Really Fewer Snakes and Lizards in Southwestern Amazonian Lowlands? Diversity, 11(10), 199. https://doi.org/10.3390/d11100199