Tree Diversity Reduces Fungal Endophyte Richness and Diversity in a Large-Scale Temperate Forest Experiment

 ,
,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Site Details
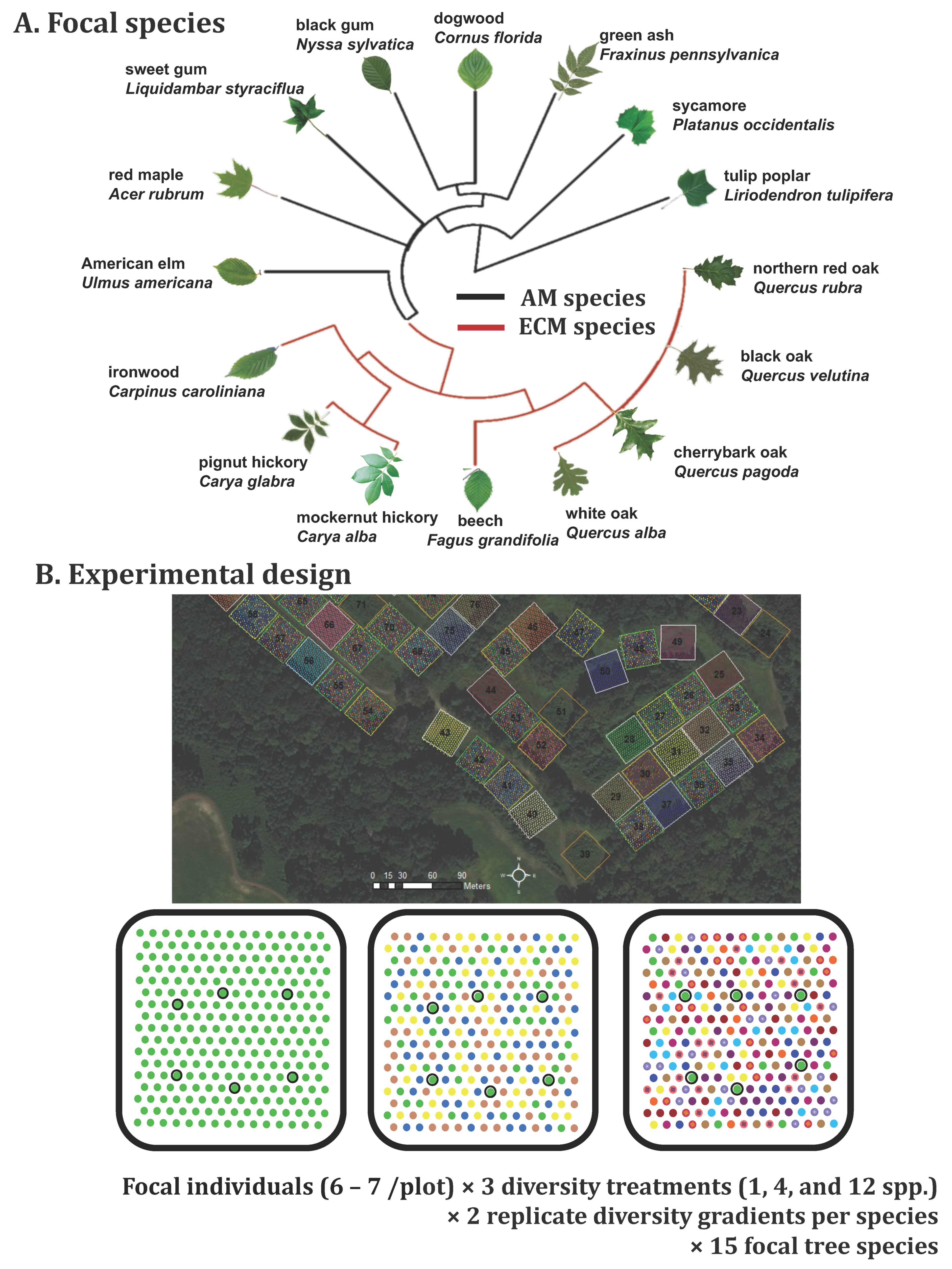
2.2. Experimental Design
2.3. Neighborhood and Plot Phylogenetic Diversity
2.4. Sampling Design
2.5. Sampling Protocol
2.6. DNA Sequencing and Data Processing
2.7. Testing the “Diversity Begets Diversity” Hypothesis (H1) and the “Tree Diversity-Endophyte Community” Hypothesis (H2)
2.8. Testing the “Tree Diversity-Endophyte Function” Hypothesis (H3)
3. Results
3.1. The “Diversity Begets Diversity” Hypothesis (H1): Increases in Plot Richness as Well as Neighborhood Richness and Phylogenetic Diversity Decreased Fungal Richness and Diversity
3.1.1. Fungal Richness
3.1.2. Shannon’s Fungal Diversity
3.1.3. Simpson’s Fungal Effective Species Number
3.1.4. Tree Neighborhood Richness and Neighborhood and Plot Phylogenetic Diversity
3.2. The “Tree Diversity–Endophyte Community” Hypothesis (H2): Host Tree Identity and Diversity Explained Little Variation in Fungal Community Structure
3.3. The “Tree Diversity‒Endophyte Function” Hypothesis (H3): Increases in Plot Richness Decreased Fungal Guild Richness and Diversity
4. Discussion
4.1. Counter to Traditional BEF Theory, Tree Richness and Phylogenetic Diversity Decreased Endophyte Diversity
4.2. Host Tree Richness Mediated the Function of Fungal Endophytes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ellis, E.C. Anthropogenic transformation of the terrestrial biosphere. Philos. Trans. R. Soc. A 2011, 369, 1010–1035. [Google Scholar] [CrossRef] [PubMed]
- Estes, J.A.; Terborgh, J.; Brashares, J.S.; Power, M.E.; Berger, J.; Bond, W.J.; Carpenter, S.R.; Essington, T.E.; Holt, R.D.; Jackson, J.B.; et al. Trophic downgrading of planet Earth. Science 2011, 333, 301–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achard, F.; Eva, H.D.; Stibig, H.J.; Mayaux, P.; Gallego, J.; Richards, T.; Malingreau, J.P. Determination of deforestation rates of the world’s humid tropical forests. Science 2002, 297, 999–1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, S.J. Tropical forests in a changing environment. Trends Ecol. Evol. 2005, 20, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Gerstner, K.; Dormann, C.F.; Stein, A.; Manceur, A.M.; Seppelt, R. Effects of land use on plant diversity—A global meta-analysis. J. Appl. Ecol. 2014, 51, 1690–1700. [Google Scholar] [CrossRef]
- Decaens, T.; Martins, M.B.; Feijoo, A.; Oszwald, J.; Doledec, S.; Mathieu, J.; Arnaud de Sarte, X.; Bonilla, D.; Brown, G.G.; Cuellar Criollo, Y.A.; et al. Biodiversity loss along a gradient of deforestation in Amazonian agricultural landscapes. Conserv. Biol. 2018, 32, 1380–1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilman, D.; Isbell, F.; Cowles, J.M. Biodiversity and ecosystem functioning. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 471–493. [Google Scholar] [CrossRef]
- Mori, A.S.; Lertzman, K.P.; Gustafsson, L. Biodiversity and ecosystem services in forest ecosystems: A research agenda for applied forest research. J. Appl. Ecol. 2017, 54, 12–27. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Matulich, K.L.; Hooper, D.U.; Byrnes, J.E.; Duffy, E.; Gamfeldt, L.; Balvanera, P.; O’Connor, M.I.; Gonzalez, A. The functional role of producer diversity in ecosystems. Am. J. Bot. 2011, 98, 572–592. [Google Scholar] [CrossRef] [Green Version]
- Hooper, D.U.; Chapin, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Rodriguez, R.J.; White, J.F.; Arnold, A.E.; Redman, R.S. Fungal endophytes: Diversity and functional roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef] [PubMed]
- Griffin, E.A.; Traw, M.B.; Morin, P.J.; Pruitt, J.N.; Wright, S.J.; Carson, W.P. Soil fertility mediates sapling responses to foliar bacteria in a tropical forest: Experimental evidence and ecological implications. Ecology 2016, 97, 2998–3008. [Google Scholar] [CrossRef] [PubMed]
- Griffin, E.A.; Wright, S.J.; Morin, P.J.; Carson, W.P. Pervasive interactions between foliar microbes and soil nutrients mediate leaf production and herbivore damage in a tropical forest. New Phytol. 2017, 216, 99–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffin, E.A.; Harrison, J.G.; Kembel, S.W.; Carrell, A.A.; Wright, S.J.; Carson, W.P. Plant host identity and soil macronutrients explain little variation in sapling endophyte community composition: Is disturbance an alternative explanation? J. Ecol. 2019, 107, 1876–1889. [Google Scholar] [CrossRef]
- Laforest-Lapointe, I.; Paquette, A.; Messier, C.; Kembel, S.W. Leaf bacterial diversity mediates plant diversity and ecosystem function relationships. Nature 2017, 546, 145–147. [Google Scholar] [CrossRef]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirtilla, A.M.; Compant, S.; Campisano, A.; Doring, M.; Sessitsch, A. The hidden world within plants: Ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [Green Version]
- Arnold, A.E.; Herre, A.H. Canopy cover and leaf age affect colonization by tropical fungal endophytes: Ecological pattern and process in Theobroma cacao (Malvaceae). Mycologia 2003, 95, 388–398. [Google Scholar] [CrossRef]
- Lindow, S.E.; Brandl, M.T. Microbiology of the phyllosphere. Appl. Environ. Microbiol. 2003, 69, 1875–1883. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, G.S.; Reynolds, D.R. Nocturnal fungi: Airborne spores in the canopy and understory of a tropical rain forest. Biotropica 2005, 37, 462–464. [Google Scholar] [CrossRef]
- Griffin, E.A.; Carson, W.P. Tropical tree endophytes: Cryptic drivers of forest diversity, species composition, and ecosystem function. In Endophytes of Forest Trees: Biology and Applications, 2nd ed.; Pirttila, A.M., Frank, A.C., Eds.; Springer: Dordrecht, The Netherlands, 2018; pp. 63–103. [Google Scholar]
- Christian, N.; Herre, E.A.; Clay, K. Foliar endophytic fungi alter patterns of nitrogen uptake and distribution in Theobroma cacao. New Phytol. 2019, 222, 1573–1583. [Google Scholar] [CrossRef]
- Arnold, A.E.; Mejia, L.C.; Kyllo, D.; Rojas, E.I.; Maynard, Z.; Robbins, N.; Herre, E.A. Fungal endophytes limit pathogen damage in a tropical tree. Proc. Natl. Acad. Sci. USA 2003, 100, 15649–15654. [Google Scholar] [CrossRef] [Green Version]
- Christian, N.; Herre, E.A.; Mejia, L.C.; Clay, K. Exposure to the leaf litter microbiome of healthy adults protects seedlings from pathogen damage. Proc. Natl. Acad. Sci. USA 2017, 284, 20170641. [Google Scholar] [CrossRef] [PubMed]
- Redford, A.J.; Bowers, R.M.; Knight, R.; Linhart, Y.; Fierer, N. The ecology of the phyllosphere: Geographic and phylogenetic variability in the distribution of bacteria on tree leaves. Environ. Microbiol. 2010, 12, 2885–2893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kembel, S.W.; O’Connor, T.K.; Arnold, H.K.; Hubbell, S.P.; Wright, S.J.; Green, J.L. Relationships between phyllosphere bacterial communities and plant functional traits in a neotropical forest. Proc. Natl. Acad. Sci. USA 2014, 111, 13715–13720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laforest-Lapointe, I.; Messier, C.; Kembel, S.W. Temperate tree phyllosphere bacterial community composition. Microbiome 2016, 4, 27. [Google Scholar] [CrossRef] [Green Version]
- Dyer, L.A.; Singer, M.S.; Lill, J.T.; Stireman, J.O.; Gentry, G.L.; Marquis, R.J.; Ricklefs, R.E.; Greeney, H.F.; Wagner, D.L.; Morais, H.C.; et al. Host specificity of Lepidoptera in tropical and temperate forests. Nature 2007, 448, 696–699. [Google Scholar] [CrossRef]
- Forister, M.L.; Novotny, V.; Panorska, A.K.; Baje, L.; Basset, Y.; Butterill, P.T.; Cizek, L.; Coley, P.D.; Dem, F.; Diniz, I.R.; et al. The global distribution of diet breadth in insect herbivores. Proc. Natl. Acad. Sci. USA 2015, 112, 442–447. [Google Scholar] [CrossRef] [Green Version]
- Hausmann, N.T.; Hawkes, C.V. Plant neighborhood control of arbuscular mycorrhizal community composition. New Phytol. 2009, 183, 1188–1200. [Google Scholar] [CrossRef]
- Liu, Y.; Shi, G.; Mao, L.; Cheng, G.; Jiang, S.; Ma, X.; An, L.; Du, G.; Collins Johnson, N.; Feng, H. Direct and indirect influences of 8 yr of nitrogen and phosphorus fertilization on Glomeromycota in an alpine meadow ecosystem. New Phytol. 2012, 194, 523–535. [Google Scholar] [CrossRef]
- Peay, K.G.; Bidartondo, M.I.; Arnold, E.A. Not every fungus is everywhere: Scaling to the biogeography of fungal-plant interactions across roots, shoots, and ecosystems. New Phytol. 2010, 185, 878–882. [Google Scholar] [CrossRef]
- Adams, R.I.; Miletto, M.; Taylor, J.W.; Bruns, T.D. Dispersal in microbes: Fungi in indoor air are dominated by outdoor air and show dispersal limitation at short distances. ISME J. 2013, 7, 1262–1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffin, E.A.; Carson, W.P. The ecology and natural history of foliar bacteria with a focus on tropical forests and agroecosystems. Bot. Rev. 2015, 81, 105–149. [Google Scholar] [CrossRef]
- Srivastava, D.S.; Cadotte, M.W.; MacDonald, A.A.M.; Marushia, R.G.; Mirotchnick, N. Phylogenetic diversity and the functioning of ecosystems. Ecol. Lett. 2012, 15, 637–648. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, G.S.; Webb, C.O. Phylogenetic signal in plant pathogen-host range. Proc. Natl. Acad. Sci. USA 2007, 104, 4979–4983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Grossman, J.J.; Vanhellemont, M.; Barsoum, N.; Bauhus, J.; Bruelheide, H.; Castagneyrol, B.; Cavender-Bares, J.; Eisenhauer, N.; Ferlian, O.; Gravel, D.; et al. Synthesis and future research directions linking tree diversity to growth, survival, and damage in a global network of tree diversity experiments. Environ. Exp. Bot. 2018, 152, 68–89. [Google Scholar] [CrossRef]
- Paquette, A.; Hector, A.; Castagneyrol, B.; Vanhellemont, M.; Koricheva, J.; Scherer-Lorenzen, M.; Verheyen, K. A million and more trees for science. Nat. Ecol. Evol. 2018, 2, 763–766. [Google Scholar] [CrossRef]
- Erikson, D.L.; Jones, F.A.; Swenson, N.G.; Pei, N.; Bourg, N.A.; Chen, W.; Davies, S.J.; Ge, X.J.; Hao, Z.; Howe, R.W.; et al. Comparative evolutionary diversity and phylogenetic structure across multiple forest dynamics plots: A meta-phylogeny approach. Front. Genet. 2014, 5, 358. [Google Scholar] [CrossRef] [Green Version]
- Devaney, J.L.; Pullen, J.; Cook-Patton, S.C.; Burghardt, K.T.; Parker, J.D. Tree diversity exacerbates deer damage but facilitates establishment of late successional species in a forest experiment. Ecology. in review.
- Dickie, I.A.; Koide, R.T.; Fayish, A.C. Vesicular-arbuscular mycorrhizal infection of Quercus rubra seedlings. New Phytol. 2001, 151, 257–264. [Google Scholar] [CrossRef]
- Dickie, I.A.; Koide, R.T.; Steiner, K.C. Influences of established trees on mycorrhizas, nutrition, and growth of Quercus rubra seedlings. Ecol. Monog. 2002, 72, 505–521. [Google Scholar] [CrossRef]
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faith, P.D. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Swenson, N.G. Functional and Phylogenetic Ecology in R; Springer: New York, NY, USA, 2014. [Google Scholar]
- Zanne, A.E.; Tank, D.C.; Cornwell, W.K.; Eastman, J.M.; Smith, S.A.; FitzJohn, R.G.; McGlinn, D.J.; O’Meara, B.C.; Moles, A.T.; Reich, P.B.; et al. Three keys to the radiation of angiosperms into freezing environments. Nature 2014, 506, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Webb, C.O.; Ackerly, D.D.; Kembel, S.W. Phylocom: Software for the analysis of phylogenetic community structure and trait evolution. Bioinformatics 2008, 24, 2098–2100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seabloom, E.W.; Condon, B.; Kinkel, L.; Komatsu, K.J.; Lumibao, C.Y.; May, G.; McCulley, R.L.; Borer, E.T. Effects of nutrient supply, herbivory, and host community on fungal endophyte diversity. Ecology 2019, 100, e02758. [Google Scholar] [CrossRef] [PubMed]
- Borcard, D.; Legendre, P. All-scale spatial analysis of ecological data by means of principal coordinates of neighbour matrices. Ecol. Model. 2002, 153, 51–68. [Google Scholar] [CrossRef]
- Adespatial: Multivariate Multiscale Spatial Analysis. Available online: https://rdrr.io/cran/adespatial (accessed on 4 June 2019).
- Arnold, A.E.; Lutzoni, F. Diversity and host range of foliar fungal endophytes: Are tropical leaves biodiversity hotspots? Ecology 2007, 88, 541–549. [Google Scholar] [CrossRef]
- Kaewkla, O.; Franco, C.M. Rational approaches to improving the isolation of endophytic actinobacteria from Australian native trees. Microb. Ecol. 2013, 65, 384–393. [Google Scholar] [CrossRef]
- Shaffer, J.P.; Sarmiento, C.; Zalamea, P.C.; Gallery, R.E.; Davis, A.S.; Baltrus, D.A.; Arnold, A.E. Diversity, specificity, and phylogenetic relationships of endohyphal bacteria in fungi that inhabit tropical seeds and leaves. Front. Ecol Evol. 2016, 4, 116. [Google Scholar] [CrossRef] [Green Version]
- Shaffer, J.P.; U’Ren, J.M.; Gallery, R.E.; Baltrus, D.A.; Arnold, A.E. An endohyphal bacterium (Chitinophaga, Bacteriodetes) alters carbon source use by Fusarium keratoplasticum (F. solanum species complex, Nectriaceae). Front. Microbiol. 2017, 8, 350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vancov, T.; Keen, B. Amplification of soil fungal community DNA using the ITS86F and ITS4 primers. FEMS Microbiol. Ecol. 2009, 296, 91–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Beeck, M.O.; Lievens, B.; Busschaert, P.; Declerck, S.; Vangronsveld, J.; Colpaert, J.V. Comparison and validation of some ITS primer pairs useful for fungal metabarcoding studies. PLoS ONE 2014, 9, e97629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C.; Flyvbjerg, H. Error filtering, pair assembly, and error correction for next-generation sequencing reads. Bioinformatics 2015, 31, 3476–3482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. UNOISE2: Improved error-correction for Illumina 16S and ITS amplicon sequencing. BioRxiv 2016, 081257. [Google Scholar]
- Callahan, B.J.; McMurdie, P.J.; Holmes, S.P. Exact sequence variants should replace operational taxonomic units in marker-gene data analysis. ISME J. 2017, 11, 2639–2643. [Google Scholar] [CrossRef] [Green Version]
- Deshpande, V.; Wang, Q.; Greenfield, P.; Charleston, M.; Porras-Alfaro, A.; Kuske, C.R.; Cole, J.R.; Midgley, D.J.; Tran-Dinh, N. Fungal identification using a Bayesian classifier and the Warcup training set of internal transcribed spacer sequences. Mycologia 2016, 108, 1–5. [Google Scholar] [CrossRef]
- Abarenkov, K.; Henrik Nilsson, R.; Larsson, K.H.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Høiland, K.; Kjøller, R.; Larsson, E.; Pennanen, T.; et al. The UNITE database for molecular identification of fungi–recent updates and future perspectives. New Phytol. 2010, 186, 281–285. [Google Scholar] [CrossRef]
- Edgar, R.C. SUNTAX: A simple non-Bayesian taxonomy classifier for 16S and ITS sequenced. BioRxiv 2016, 074161. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.; Wagner, H. Package ‘vegan’. Community Ecol. Package 2013, 10, 631–637. [Google Scholar]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Aust. Ecol. 2001, 26, 32–46. [Google Scholar]
- Jost, L. Entropy and diversity. Oikos 2006, 113, 363–375. [Google Scholar] [CrossRef]
- Jost, L. Partitioning diversity and independent alpha and beta diversity components. Ecology 2007, 88, 2427–2439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacqmin-Gadda, H.; Sibilot, S.; Proust, C.; Molina, J.M.; Thiebeaut, R. Robustness of the linear mixed model to misspecified error distribution. Comput. Stat. Data Anal. 2007, 51, 5142–5154. [Google Scholar] [CrossRef] [Green Version]
- Tedersoo, L.; Smith, M.E. Lineages of ectomycorrhizal fungi revisited: Foraging strategies and novel lineages revealed by sequences from belowground. Fungal Biol. Rev. 2013, 27, 83–99. [Google Scholar] [CrossRef] [Green Version]
- Carrell, A.A.; Frank, A.C. Pinus flexilis and Picea engelmannii share a simple and consistent needle endophyte microbiota with a potential role in nitrogen fixation. Front. Microbiol. 2014, 5, 333. [Google Scholar] [CrossRef] [Green Version]
- De Meyer, S.E.; De Beuf, K.; Vekeman, B.; Willems, A. A large diversity of non-rhizobial endophytes found in legume root nodules in Flanders (Belgium). Soil Biol. Biochem. 2015, 83, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Agler, M.T.; Ruhe, J.; Kroll, S.; Morhenn, C.; Kim, S.-T.; Weigel, D.; Kemen, E.M. Microbial hub taxa link host and abiotic factors to plant microbiome variation. PLoS Biol. 2016, 14, e1002352. [Google Scholar] [CrossRef] [Green Version]
- Unterseher, M.; Siddique, A.B.; Brachmann, A.; Persoh, D. Diversity and composition of the leaf mycobiome of beech (Fagus sylvatica) are affected by local habitat conditios and leaf biochemistry. PLoS ONE 2016, 11, e0152878. [Google Scholar] [CrossRef] [PubMed]
- Allan, E.; Weisser, W.W.; Weigelt, A.; Roscher, C.; Fisher, M.; Hillebrand, H. More diverse plant communities have higher functioning over time due to turnover in complementary dominant species. Proc. Natl. Acad. Sci. USA 2011, 108, 17034–17039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reich, P.B.; Tilman, D.; Isbell, F.; Mueller, K.; Hobbie, S.E.; Flynn, D.F.B.; Eisenhauer, N. Impacts of biodiversity loss escalate through time as redundancy fades. Science 2012, 336, 589–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravenek, J.M.; Bessler, H.; Engels, C.; Scherer-Lorenzen, M.; Gessler, A.; Gockele, A.; De Luca, E.; Temperton, V.M.; Ebeling, A.; Roscher, C.; et al. Long-term study of root biomass in a biodiversity experiment reveals shifts in diversity effects over time. Oikos 2014, 12, 1528–1536. [Google Scholar] [CrossRef]
- Weisser, W.W.; Roscher, S.T.; Meyer, S.T.; Ebeling, A.; Luo, G.; Allan, E.; Bebler, H.; Barnard, R.L.; Buchmann, N.; Buscot, F.; et al. Biodiversity effects on ecosystem functioning in a 15-year grassland experiment: Patterns, mechanisms, and open questions. Basic Appl. Ecol. 2017, 23, 1–73. [Google Scholar] [CrossRef]
- Siemann, E.; Tilman, D.; Haarstad, J.; Ritchie, M. Experimental tests of the dependence of arthropod diversity on plant diversity. Am. Nat. 1998, 152, 738–750. [Google Scholar] [CrossRef]
- Parker, J.D.; Duffy, J.E.; Orth, R.J. Plant species diversity and composition: Experimental effects on marine epifaunal assemblages. Mar. Ecol. Prog. Ser. 2001, 224, 55–67. [Google Scholar] [CrossRef]
- Reusch, T.B.; Ehlers, A.; Hämmerli, A.; Worm, B. Ecosystem recovery after climatic extremes enhanced by genotypic diversity. Proc. Natl. Acad. Sci. USA 2005, 102, 2826–2831. [Google Scholar] [CrossRef] [Green Version]
- Scherber, C.; Eisenhauer, N.; Weisser, W.W.; Schmid, B.; Voigt, W.; Fischer, M.; Schulze, E.D.; Roscher, C.; Weigelt, A.; Allan, E.; et al. Bottom-up effects of plant diversity on multitrophic interactions in a biodiversity experiment. Nature 2010, 468, 553–556. [Google Scholar] [CrossRef]
- Hutchinson, G.E. Homage to Santa Rosalia or why are there so many kinds of animals? Am. Nat. 1959, 93, 145–159. [Google Scholar] [CrossRef] [Green Version]
- Strong, D.R.; Lawton, J.H.; Southwood, S.R. Insects on Plants. Community Patterns and Mechanisms; Blackwell Scientific Publications: Hoboken, NJ, USA, 1984. [Google Scholar]
- Higgins, K.L.; Arnold, A.E.; Coley, P.D.; Kursar, T.A. Communities of fungal endophytes in tropical forest grasses: Highly diverse host-and habitat generalists characterized by strong spatial structure. Fungal Ecol. 2014, 8, 1–11. [Google Scholar] [CrossRef]
- Seabloom, E.W.; Borer, E.T.; Gross, K.; Kendig, A.E.; Lacroix, C.; Mitchell, C.E.; Mordecai, E.A.; Power, A.G. The community ecology of pathogens: Coinfection, coexistence, and community composition. Ecol. Lett. 2015, 18, 401–415. [Google Scholar] [CrossRef] [PubMed]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Augspurger, C.K. Seedling survival of tropical tree species: Interactions of dispersal distance, light-gaps, and pathogens. Ecology 1984, 65, 1705–1712. [Google Scholar] [CrossRef]
- Augspurger, C.K.; Kelly, C.K. Pathogen mortality of tropical tree seedlings: Experimental studies of the effects of dispersal distance, seedling density, and light conditions. Oecologia 1984, 61, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Packer, A.; Clay, K. Soil pathogens and spatial patterns of seedling mortality in a temperate tree. Nature 2000, 404, 278–281. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.J.; Beaulieu, W.T.; Bever, J.D.; Clay, K. Conspecific negative density dependence and forest diversity. Science 2012, 336, 904–907. [Google Scholar] [CrossRef] [Green Version]
- Bernays, E.A. Evolution of feeding behavior in insect herbivores. Bioscience 1998, 48, 35–44. [Google Scholar] [CrossRef]
- Bernays, E.; Graham, M. On the evolution of host specificity in phytophagous arthropods. Ecology 1988, 69, 886–892. [Google Scholar] [CrossRef]
- Schoonhoven, L.M.; Van Loon, B.; van Loon, J.J.; Dicke, M. Insect-Plant Biology; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Suryanarayanan, T.S.; Venkatesan, G.; Murali, T.S. Endophytic fungal communities in leaves of tropical forest trees: Diversity and distribution patterns. Curr. Sci. 2003, 85, 489–493. [Google Scholar]
- Suryanarayanan, T.S.; Murali, T.S.; Thirunavukkarasu, N.; Rajulu, M.G.; Venkatesan, G.; Sukumar, R. Endophytic fungal communities in woody perennials of three tropical forest types of the Western Ghats, southern India. Biodivers. Conserv. 2011, 20, 913–928. [Google Scholar] [CrossRef]
- Vincent, J.B.; Weiblen, G.D.; May, G. Host associations and beta diversity of fungal endophyte communities in New Guinea rainforest trees. Mol. Ecol. 2016, 25, 825–841. [Google Scholar] [CrossRef] [PubMed]
- Badri, D.V.; Zolla, G.; Bakker, M.G.; Manter, D.K.; Vivanco, J.M. Potential impact of soil microbiomes on the leaf metabolome and on herbivore feeding behavior. New Phytol. 2013, 198, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, C.J.; Li, B.; McMinn, R.; Brock, M.T.; Maignien, L.; Ewers, B.E.; Kliebenstein, D.; Weinig, C. The effect of rhizosphere microbes outweighs host plant genetics in reducing insect herbivory. Mol. Ecol. 2019, 7, 1801–1811. [Google Scholar] [CrossRef] [PubMed]
- Schoenherr, A.P.; Rizzo, E.; Jackson, N.; Manosalva, P.; Gomez, S.K. Mycorrhiza-induced resistance in potato involves priming of defense responses against cabbage looper (Noctuidae: Lepidoptera). Environ. Entomol. 2019, 48, 370–381. [Google Scholar] [CrossRef]
- Bakker, P.A.; Berendsen, R.L.; Doornbos, R.F.; Wintermans, P.C.; Pieterse, C.M. The rhizosphere revisited: Root microbiomics. Front. Plant Sci. 2013, 4, 165. [Google Scholar] [CrossRef] [Green Version]
- Pieterse, C.M.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.; Bakker, P.A. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [Green Version]
- Pereira, A.; Ferreira, I.; Marcelino, F.; Valentão, P.; Andrade, P.; Seabra, R.; Estevinho, L.; Bento, A.; Pereira, J. Phenolic compounds and antimicrobial activity of olive (Olea europaea L. Cv. Cobrançosa) leaves. Molecules 2007, 12, 1153–1162. [Google Scholar] [CrossRef]
- Meier, C.L.; Bowman, W.D. Phenolic-rich leaf carbon fractions differentially influence microbial respiration and plant growth. Oecologia 2008, 158, 95–107. [Google Scholar] [CrossRef]
- Mercier, J.; Lindow, S.E. Role of leaf surface sugars in colonization of plants by bacterial epiphytes. Appl. Environ. Microbiol. 2000, 66, 369–374. [Google Scholar] [CrossRef] [Green Version]
- Leveau, J.H.; Lindow, S.E. Appetite of an epiphyte: Quantitative monitoring of bacterial sugar consumption in the phyllosphere. Proc. Natl. Acad. Sci. USA 2001, 98, 3446–3453. [Google Scholar] [CrossRef] [Green Version]
- Broz, A.K.; Broeckling, C.D.; De-la-Peña, C.; Lewis, M.R.; Greene, E.; Callaway, R.M.; Sumner, L.W.; Vivanco, J.M. Plant neighbor identity influences plant biochemistry and physiology related to defense. BMC Plant Biol. 2010, 10, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mraja, A.; Unsicker, S.B.; Reichelt, M.; Gershenzon, J.; Roscher, C. Plant community diversity influences allocation to direct chemical defence in Plantago lanceolata. PLoS ONE 2011, 6, e28055. [Google Scholar] [CrossRef] [PubMed]
- McGarigal, K.; Marks, B.J. Fragstats: Spatial Pattern Analysis Program for Quantifying Landscape Structure; US Department of Agriculture Reference Manual; Forest Science Department, Oregon State University: Corvallis, OR, USA, 1994. [Google Scholar]
- Nagendra, H. Opposite trends in response for the Shannon and Simpson indices of landscape diversity. Appl. Geogr. 2002, 22, 175–186. [Google Scholar] [CrossRef]
- Hodge, A.; Campbell, C.D.; Fitter, A.H. An arbuscular mycorrhizal fungus accelerates decomposition and acquires nitrogen directly from organic material. Nature 2001, 413, 297–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craine, J.M.; Elmore, A.J.; Aidar, M.P.; Bustamante, M.; Dawson, T.E.; Hobbie, E.A.; Kahmen, A.; Mack, M.C.; McLauchlan, K.K.; Michelsen, A.; et al. Global patterns of foliar nitrogen isotopes and their relationships with climate, mycorrhizal fungi, foliar nutrient concentrations, and nitrogen availability. New Phytol. 2009, 183, 980–992. [Google Scholar] [CrossRef] [PubMed]
- Read, D.J.; Perez-Moreno, J. Mycorrhizas and nutrient cycling in ecosystems—A journey towards relevance? New Phytol. 2003, 157, 475–492. [Google Scholar] [CrossRef]
- Kembel, S.W.; Mueller, R.C. Plant traits and taxonomy drive host associations in tropical phyllosphere fungal communities. Botany 2014, 92, 303–311. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, E.R.; Younginger, B.S.; LeRoy, C.J. Fungal endophyte-infected leaf litter alters in-stream microbial communities and negatively influences aquatic fungal sporulation. Oikos 2019, 128, 405–415. [Google Scholar] [CrossRef]
- Anderson-Teixeira, K.J.; Davies, S.J.; Bennett, A.C.; Gonzalez-Akre, E.B.; Muller-Landau, H.C.; Wright, S.J.; Abu Salim, K.; Almeyda Zambrano, A.M.; Alonso, A.; Baltzer, J.L.; et al. CTFS-Forest GEO: A worldwide network monitoring forests in an era of global change. Glob. Chang. Biol. 2015, 21, 528–549. [Google Scholar] [CrossRef] [Green Version]
- Kral, K.; Shue, J.; Vrska, T.; Gonzalez-Akre, E.B.; Parker, G.G.; McShea, W.J.; McMahon, S.M. Fine-scale patch mosaic of developmental stages in Northeast American secondary temperate forests: The European perspective. Europ. J. For. Res. 2016, 135, 981–996. [Google Scholar] [CrossRef]
- Latz, E.; Eisenhauer, N.; Rall, B.C.; Allan, E.; Roscher, C.; Scheu, S.; Jousset, A. Plant diversity improves protection against soil-borne pathogens by fostering antagonistic bacterial communities. J. Ecol. 2012, 100, 597–604. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Wright, J.P.; Cadotte, M.W.; Carroll, I.T.; Hector, A.; Srivastava, D.S.; Loreau, M.; Weis, J.J. Impacts of plant diversity on biomass production increase through time because of species complementarity. Proc. Natl. Acad. Sci. USA 2007, 104, 18123–18128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, K.E.; Tilman, D.; Fornara, D.; Hobbie, S.E. Root depth distribution and the diversity-productivity relationship in a long-term grassland experiment. Ecology 2013, 94, 787–793. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Response | Intercept | Diversity = 4 | Diversity = 12 | ECM | Height | MEM 1 | MEM 2 | MEM 3 |
|---|---|---|---|---|---|---|---|---|
| Richness | 119.2 *** (107.1, 131.4) | −6. 9(1.5, −15.3) | −15.8 *** (−7.6, −24.0) | 28.5 ** (8.3, 48.7) | −4.8 (−11.6, 2.0) | 3.2 (−3.6, 10.0) | 2.6 (−2.0, 7.1) | −5.1 ** (−10.2, 0.0) |
| Shannon’s equivalent | 14.4 *** (12.1, 16.8) | −1.9 ** (−3.8, 0.0) | −2.9 *** (−4.8, −1.0) | 4.8 ** (0.7, 8.8) | −0.3 (−1.7, 1.2) | 0.4 (−1.0, 1.9) | 0.8 (−0.2, 1.8) | −1.8 *** (−3.0, −0.7) |
| Simpson’s sp. number | 1.8 *** (0.9, 2.9) | 1.2 ** (0.0, 2.5) | −0.1 (−1.3, 1.1) | −0. 9(−2.4, 0.6) | −0.0 (−0.7, 0.6) | −0.4 (−1.1, 0.3) | 0.0 (−0.5, 0.6) | 0.3 (−0.3, 0.9) |
| Response | Intercept | SES_PD | Height | MEM 1 | MEM 2 | MEM 3 |
|---|---|---|---|---|---|---|
| Neighborhood | ||||||
| Richness | 110.8 *** (99.9, 121.6) | −5.8 *** (−9.4, −2.2) | −5.6 (−9.4, −2.2) | 2.2 (−4.4, 8.8) | 1.9 (−2.6, 6.5) | −4.9 * (−10.0, 0.2) |
| Shannon’s equivalent. | 12.8 *** (10.7, 14.9) | −1 *** (−1.8, −0.2) | −0.5 (−2.0, 1.0) | 0.2 (−1.2, 1.7) | 0.7 (−0.3, 1.7) | −1.7 *** (−2.8, −0.6) |
| Simpson’s sp. number. | 2.3 *** (1.6, 3.0) | 0 (−0.6, 0.5) | 0 (−0.7, 0.7) | −0.3 (−0.9, 0.4) | −0.1 (−0.6, 0.5) | 0.2 (−0.4, 0.8) |
| Plot | ||||||
| Richness | 114.8 *** (95.7, 133.9) | −7.7 ** (−14.8, −0.5) | −7 * (−14.1, 0.2) | 0.1 (−7.5, 7.7) | 4.1 (−2.9, 11) | −5.2 (−13.8, 3.5) |
| Shannon’s equivalent | 11 *** (7.2, 14.8) | −1 (−2.4, 0.4) | −0.5 (−1.0, 09) | −0.2 (−1.7, 1.3) | 0.4 (−0.9, 1.7) | −1 (−2.7, 0.7) |
| Simpson’s sp. number. | 1.3 *** (1.2, 1.4) | 0 (0, 0.1) | 0 (0, 0.1) | 0 (−0.1, 0.1) | 0 (−0.1, 0) | 0 (0, 0.1) |
| Response | Intercept | Diversity = 4 | Diversity = 12 | Height | MEM 1 | MEM 2 | MEM 3 |
|---|---|---|---|---|---|---|---|
| Pathogens | 60.6 *** (54.8, 66.4) | −3.0 (−7.4, 1.5) | −6.6 *** (−11.0, −2.2) | −2.7 (−6.2, 0.7) | 1.6 (−1.7, 4.9) | 0.6 (−1.8, 3.0) | −3.2 ** (−5.8, −0.6) |
| Saprotrophs | 55.3 *** (50.7, 59.9) | −2.7 (−6.8, 1.5) | −5.6 *** (−9.7, −1.5) | −2.3 (−5.3, 0.7) | 0.5 (−2.4, 3.4) | 1.1 (−1.1, 3.2) | −2.4 ** (−4.8, −0.1) |
| Parasites | 12.3 *** (10.7, 13.9) | −0.2 (−1.7, 1.3) | −1.4 * (−2.9, 0.1) | −0.4 (−1.4, 0.7) | −0.6 (−1.6, 0.5) | 0.4 (−0.4, 1.1) | −0.9 ** (−1.8, −0.1) |
| Lichens | 12.2 *** (10.6, 13.9) | −0.4 (−1.9, 1.1) | −1.5 ** (−3.0, −0.01) | −0.5 (−1.5, 0.6) | −0.5 (−1.5, 0.6) | 0.3 (−0.5, 1.1) | −1.2 *** (−2.0, −0.3) |
| Symbionts | 28.7 *** (25.2, 32.2) | −1.3 (−3.8, 1.2) | −2.893 ** (−5.36, −0.426) | −1.9 * (−3.9, 0.1) | −0.04 (−2.0, 1.9) | 0.3 (−1.1, 1.6) | −1.7 ** (−3.2, −0.2) |
| Response | Intercept | Diversity = 4 | Diversity = 12 | Height | MEM 1 | MEM 2 | MEM 3 |
|---|---|---|---|---|---|---|---|
| Guild Diversity (Shannon’s) | 3.57 *** | −0.19 * | −0.24 ** | 0.0 | −0.07 | 0.08 | −0.02 |
| Pathogens | 7.4 *** (6.4, 8.3) | −0.6 (−1.5, 0.4) | −1.0 ** (−1.9, −0.1) | −0.2 (−0.9, 0.4) | 0.2 (−0.4, 0.9) | −1.2 *** (−1.7, −0.7) | −0.6 ** (−1.1, −0.1) |
| Saprotrophs | 7.7 *** (6.8, 8.5) | −0.5 (−1.4, 0.5) | −1.5 *** (−2.4, −0.5) | −0.1 (−0.7, 0.5) | −0.1 (−0.6, 0.5) | −1.2 *** (−1.7, −0.7) | −0.8 *** (−1.3, −0.4) |
| Parasites | 7.7 *** (6.8, 8.5) | −0.5 (−1.4, 0.5) | −1.5 *** (−2.4, −0.5) | −0.1 (−0.7, 0.5) | −0.1 (−0.6, 0.5) | −1.2 *** (−1.7, −0.7) | −0.8 *** (−1.3, −0.4) |
| Lichens | 2.7 *** (2.4, 2.9) | −0.1 (−0.4, 0.1) | −0.1 (−0.4, 0.1) | −0.01 (−0.1, 0.1) | 0.1 (−0.03, 0.2) | −0.1 (−0.2, 0.1) | −0.1 (−0.2, 0.1) |
| Symbionts | 4.7 *** (4.2, 5.1) | −0.1 (−0.7, 0.5) | −0.3 (−0.9, 0.3) | −0.1 (−0.5, 0.2) | −0.3 * (−0.6, 0.03) | −0.4 *** (−0.7, −0.1) | −0.3 ** (−0.6, −0.1) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Griffin, E.A.; Harrison, J.G.; McCormick, M.K.; Burghardt, K.T.; Parker, J.D. Tree Diversity Reduces Fungal Endophyte Richness and Diversity in a Large-Scale Temperate Forest Experiment. Diversity 2019, 11, 234. https://doi.org/10.3390/d11120234
Griffin EA, Harrison JG, McCormick MK, Burghardt KT, Parker JD. Tree Diversity Reduces Fungal Endophyte Richness and Diversity in a Large-Scale Temperate Forest Experiment. Diversity. 2019; 11(12):234. https://doi.org/10.3390/d11120234
Chicago/Turabian StyleGriffin, Eric A., Joshua G. Harrison, Melissa K. McCormick, Karin T. Burghardt, and John D. Parker. 2019. "Tree Diversity Reduces Fungal Endophyte Richness and Diversity in a Large-Scale Temperate Forest Experiment" Diversity 11, no. 12: 234. https://doi.org/10.3390/d11120234
APA StyleGriffin, E. A., Harrison, J. G., McCormick, M. K., Burghardt, K. T., & Parker, J. D. (2019). Tree Diversity Reduces Fungal Endophyte Richness and Diversity in a Large-Scale Temperate Forest Experiment. Diversity, 11(12), 234. https://doi.org/10.3390/d11120234