Crypsis Decreases with Elevation in a Lizard

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
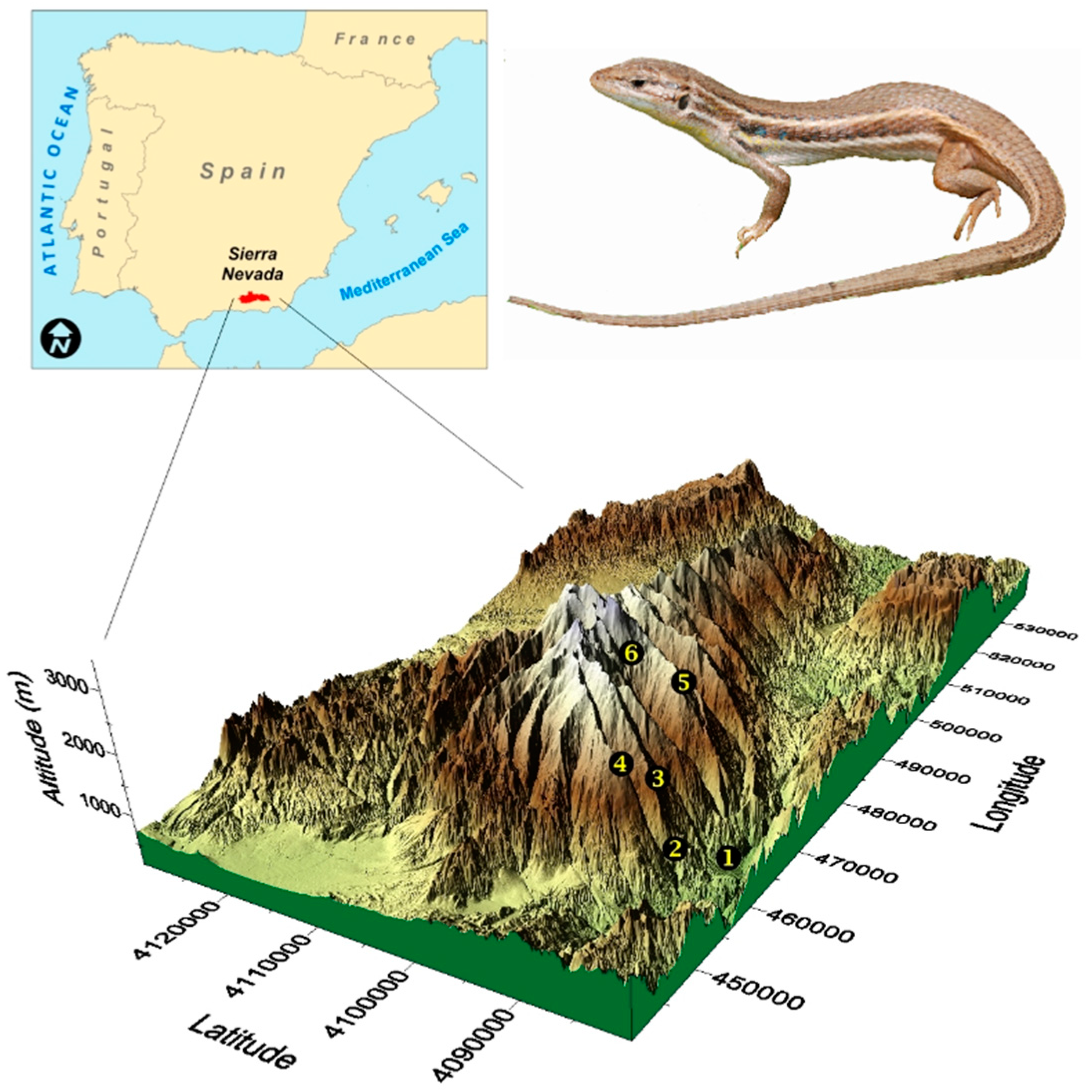
2.1. Study Species
2.2. Study Area
2.3. Sampling
2.4. Colour Measurement
2.5. Statistical Analyses
3. Results
3.1. Elevational Variation of the Environmental Background Colour
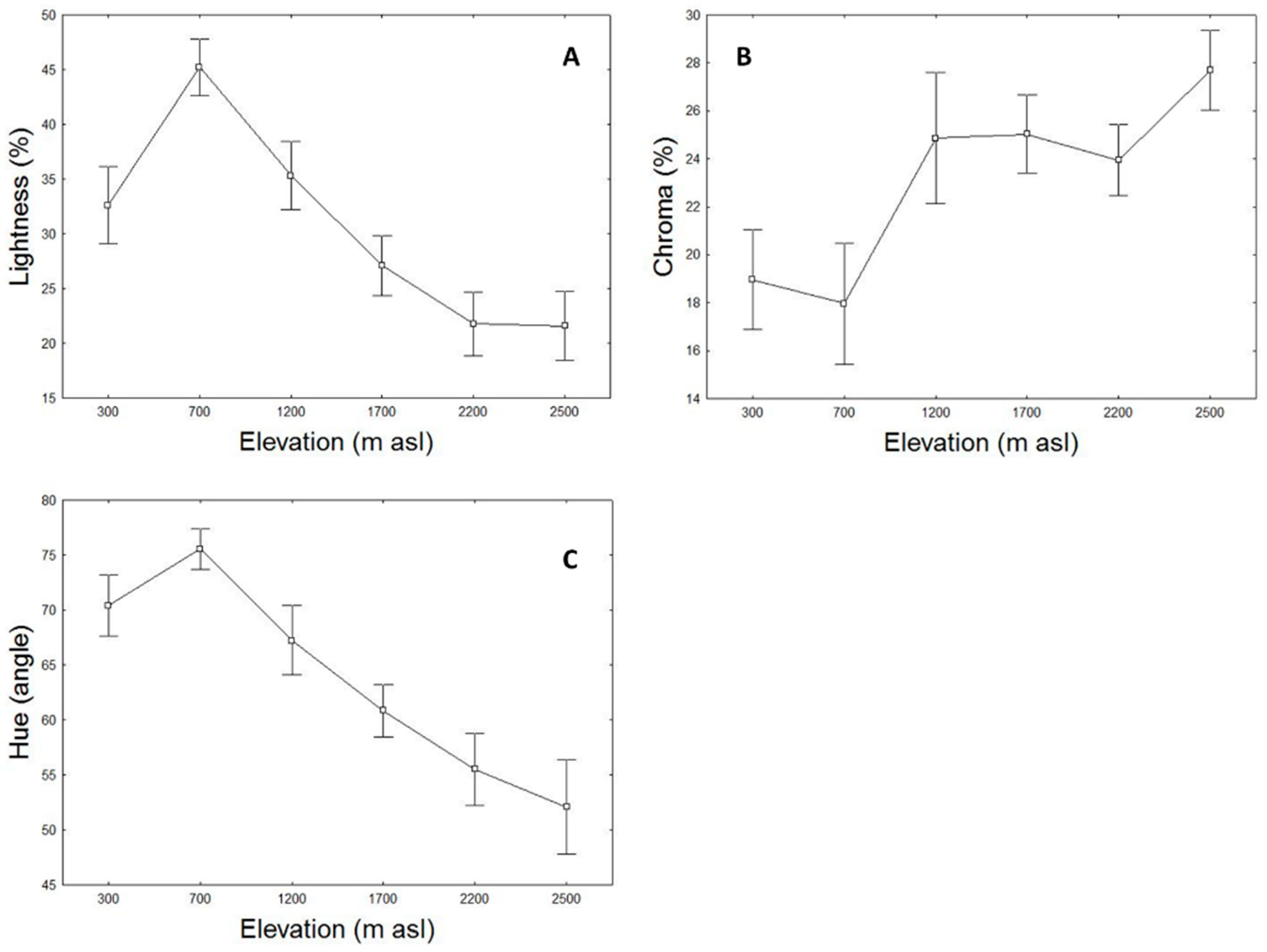
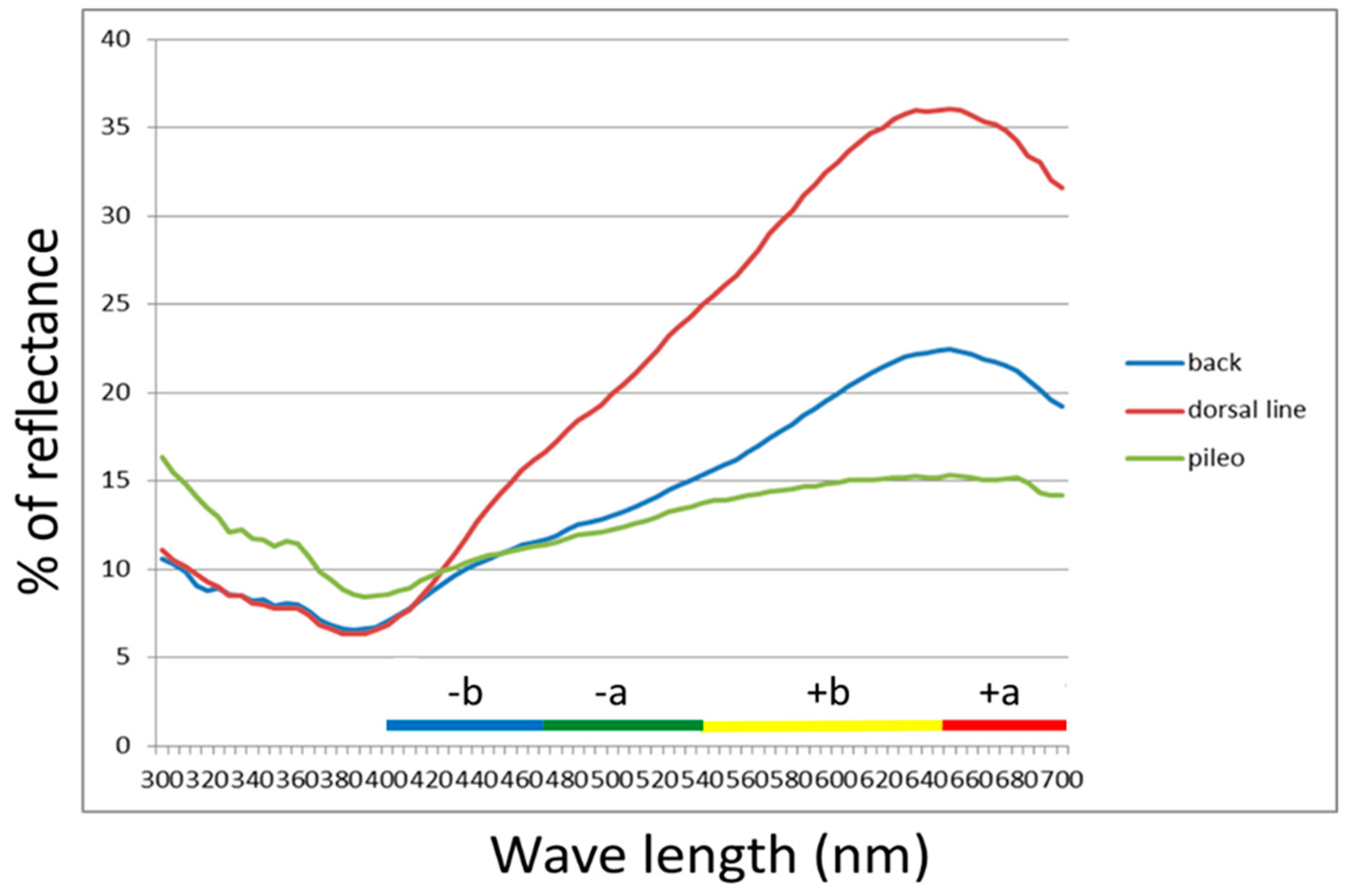
3.2. Elevational Variation of Lizard Dorsal Colour
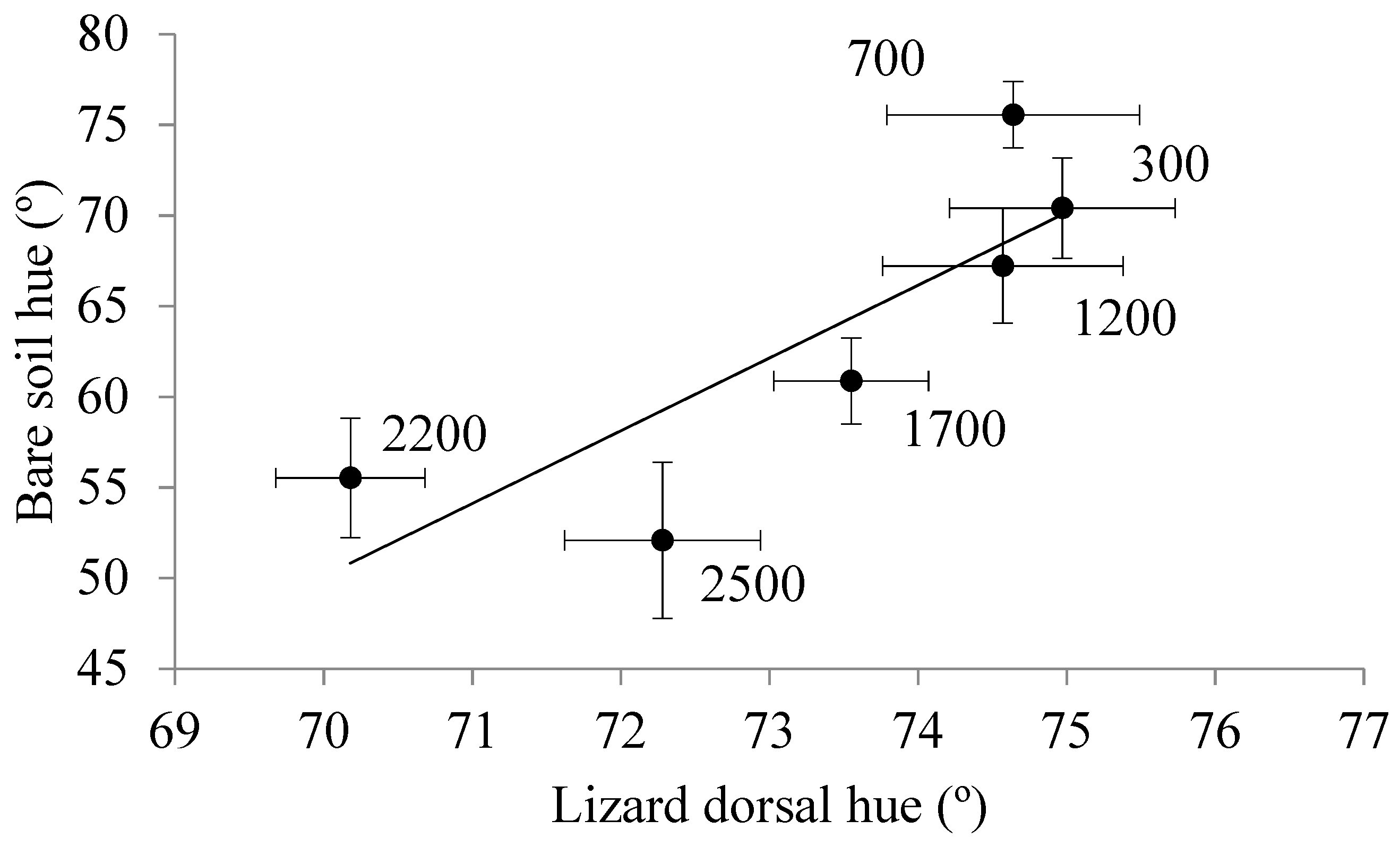
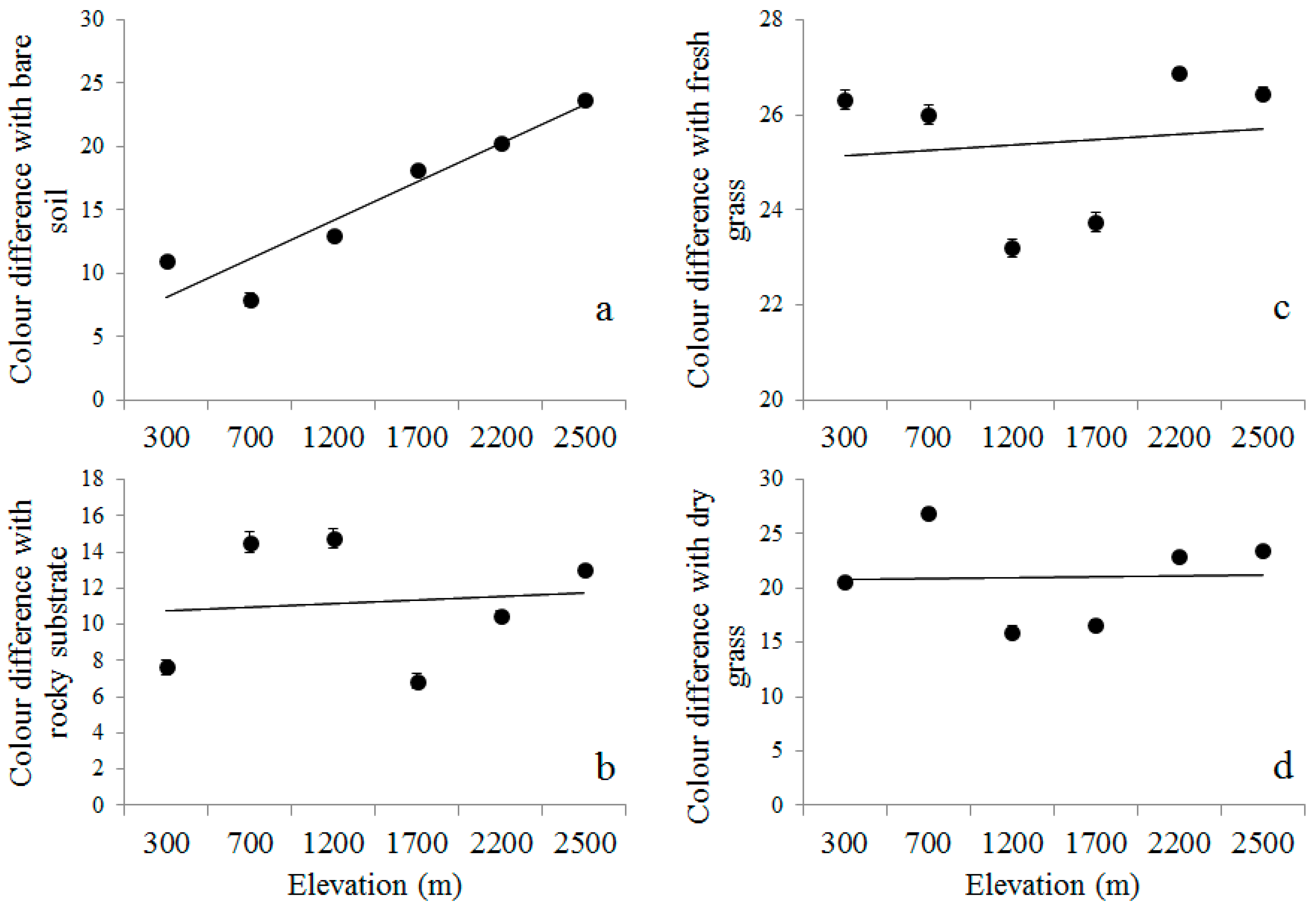
3.3. Elevational Variation in the Degree of Crypsis
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Ethics Statement
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Elevation | March | April | May | June | July | August | Total |
|---|---|---|---|---|---|---|---|
| 300 | 1/2 | 4/6 | 9/8 | 10/20 | 10/12 | - | 34/48 |
| 700 | - | 1/0 | 6/7 | 12/11 | 10/5 | 1/0 | 30/23 |
| 1200 | - | 0/2 | 9/4 | 12/8 | 6/4 | 0/1 | 27/19 |
| 1700 | 0/1 | 0/1 | 13/17 | 11/7 | 4/6 | 3/4 | 31/36 |
| 2200 | - | 0/1 | 10/8 | 16/14 | 11/8 | 2/3 | 39/34 |
| 2500 | - | - | 6/13 | 18/19 | 11/11 | - | 35/43 |
| Total | 1/3 | 5/10 | 53/57 | 79/79 | 52/46 | 6/8 | 196/203 |
Appendix B
| Elevation | SVL Males | SVL Females |
|---|---|---|
| 300 | 55 | 53 |
| 700 | 55 | 54 |
| 1200 | 55 | 56 |
| 1700 | 55 | 59 |
| 2200 | 62 | 65 |
| 2500 | 63 | 67 |
Appendix C

Appendix D
| L* | 300 | 700 | 1200 | 1700 | 2200 | 2500 | |
|---|---|---|---|---|---|---|---|
| C* | |||||||
| 300 | 0.25 | 1.91 | 0.62 | 1.93 *** | 1.53 *** | ||
| 700 | 0.70 | 0.59 | 1.29 | 0.16 * | 0.24 | ||
| 1200 | 0.06 | 0.63 | 0.99 | 1.56 *** | 1.16 * | ||
| 1700 | 0.83 | 0.13 | 0.77 | 0.88 *** | 0.49 ** | ||
| 2200 | 2.73 *** | 2.04 *** | 2.67 *** | 1.90 *** | 1.17 | ||
| 2500 | 2.85 *** | 2.16 *** | 2.79 *** | 2.02 *** | 0.12 | ||
| Hue Angle | |||||||
| 300 | 0.33 | 0.40 | 1.42 | 4.79 *** | 2.69 | ||
| 700 | 0.07 | 1.09 | 4.46 ** | 2.36 | |||
| 1200 | 1.02 | 4.38 * | 2.28 | ||||
| 1700 | 3.36 * | 1.26 | |||||
| 2200 | 2.10 | ||||||
Appendix E
| Bare Soil | 300 | 700 | 1200 | 1700 | 2200 | 2500 | |
| Rock | |||||||
| 300 | 0.01 | 2.77 * | 8.86 ** | 10.54 ** | 13.36 ** | ||
| 700 | 8.01 | 2.78 * | 8.87 ** | 10.55 ** | 13.36 ** | ||
| 1200 | 5.86 ** | 2.15 | 6.09 ** | 7.77 ** | 10.58 ** | ||
| 1700 | 1.99 | 10.00 ** | 7.85 ** | 1.68 | 4.49 ** | ||
| 2200 | 2.02 | 5.99 ** | 3.84 ** | 4.01 ** | 2.81 ** | ||
| 2500 | 5.24 ** | 8.77 | 0.62 | 7.23 ** | 3.22 ** | ||
| Fresh Grass | 300 | 700 | 1200 | 1700 | 2200 | 2500 | |
| Dry grass | |||||||
| 300 | 0.03 | 3.15 ** | 2.85 ** | 0.61 | 0.55 | ||
| 700 | 7.51 ** | 3.18 ** | 2.89 ** | 0.58 | 0.52 | ||
| 1200 | 5.47 ** | 12.99 ** | 0.29 | 3.75 ** | 3.70 ** | ||
| 1700 | 4.75 ** | 12.27 ** | 0.72 | 3.46 ** | 3.41 ** | ||
| 2200 | 1.69 | 5.83 ** | 7.16 ** | 6.44 ** | 0.05 | ||
| 2500 | 3.58 ** | 3.93 ** | 9.06 ** | 8.34 ** | 1.89 | ||
References
- Hill, J.E.; DeVault, T.L.; Belant, J.L.; Algar, A. Cause-specific mortality of the world’s terrestrial vertebrates. Glob. Ecol. Biogeogr. 2019, 28, 680–689. [Google Scholar] [CrossRef] [Green Version]
- Ruxton, G.D.; Allen, W.L.; Sherratt, T.N.; Speed, M.P. Avoiding Attack: The Evolutionary Ecology of Crypsis, Aposematism, and Mimicry; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Caro, T. Antipredator Defenses in Birds and Mammals; Chicago University Press: Chicago, IL, USA, 2005. [Google Scholar]
- Stevens, M.; Merilaita, S. (Eds.) Animal Camouflage: Mechanisms and Function; Cambridge University Press: Cambridge, UK, 2011; ISBN 0521152577. [Google Scholar]
- Cooper, W.E.J.; Blumstein, D.T. (Eds.) Escaping from Predators: An Integrative View of Escape Decisions; Cambridge University Press: Cambridge, UK, 2015. [Google Scholar]
- Merilaita, S.; Stevens, M. Crypsis through Background Matching. In Animal Camouflage: Mechanisms and Function; Stevens, M., Merilaita, S., Eds.; Cambridge University Press: Cambridge, UK, 2011; pp. 17–33. [Google Scholar]
- Cuthill, I.C. Camouflage. J. Zool. 2019, 308, 75–92. [Google Scholar] [CrossRef] [Green Version]
- Stevens, M.; Merilaita, S. Animal camouflage: Current issues and new perspectives. Phil. Trans. R. Soc. B 2009, 364, 423–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merilaita, S.; Scott-Samuel, N.E.; Cuthill, I.C. How camouflage works. Phil. Trans. R. Soc. B 2017, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endler, J.A. On the measurement and classification of colour in studies of animal colour patterns. Biol. J. Linn. Soc. 1990, 41, 315–352. [Google Scholar] [CrossRef]
- Kaufman, D.W. Adaptive coloration in Peromyscus polionotus: Experimental selection by owls. J. Mammal. 1974, 55, 271–283. [Google Scholar] [CrossRef]
- Cooper, J.M.; Allen, J.A. Selection by wild birds on artificial dimorphic prey on varied backgrounds. Biol. J. Linn. Soc. 1994, 51, 433–446. [Google Scholar] [CrossRef]
- Bond, A.B.; Kamil, A.C. Visual predators select for crypticity and polymorphism in virtual prey. Nature 2002, 415, 609–613. [Google Scholar] [CrossRef] [Green Version]
- Stuart-Fox, D.M.; Moussalli, A.; Marshall, N.J.; Owens, I.P.F. Conspicuous males suffer higher predation risk: Visual modelling and experimental evidence from lizards. Anim. Behav. 2003, 66, 541–550. [Google Scholar] [CrossRef]
- Cuthill, I.C.; Stevens, M.; Sheppard, J.; Maddocks, T.; Párraga, C.A.; Troscianko, T.S. Disruptive coloration and background pattern matching. Nature 2005, 434, 72–74. [Google Scholar] [CrossRef]
- Dimitrova, M.; Merilaita, S. Hide and seek: Properties of prey and background patterns affect prey detection by blue tits. Behav. Ecol. 2014, 25, 402–408. [Google Scholar] [CrossRef] [Green Version]
- Merilaita, S.; Dimitrova, M. Accuracy of background matching and prey detection: Predation by blue tits indicates intense selection for highly matching prey colour pattern. Funct. Ecol. 2014, 28, 1208–1215. [Google Scholar] [CrossRef] [Green Version]
- Michalis, C.; Scott-Samuel, N.E.; Gibson, D.P.; Cuthill, I.C. Optimal background matching camouflage. Proc. R. Soc. B 2017, 284, 20170709. [Google Scholar] [CrossRef] [PubMed]
- Walton, O.C.; Stevens, M. Avian vision models and field experiments determine the survival value of peppered moth camouflage. Commun. Biol. 2018, 1, 118. [Google Scholar] [CrossRef]
- Slagsvold, T.; Dale, S.; Kruszewicz, A. Predation favours cryptic coloration in breeding male pied flycatchers. Anim. Behav. 1995, 50, 1109–1121. [Google Scholar] [CrossRef]
- Cook, L.M.; Grant, B.S.; Saccheri, I.J.; Mallet, J. Selective bird predation on the peppered moth: The last experiment of Michael Majerus. Biol. Lett. 2012, 8, 609–612. [Google Scholar] [CrossRef]
- Troscianko, J.; Wilson-Aggarwal, J.; Stevens, M.; Spottiswoode, C.N. Camouflage predicts survival in ground-nesting birds. Sci. Rep. 2016, 6, 19966. [Google Scholar] [CrossRef] [Green Version]
- Norris, K.S.; Lowe, C.H. An analysis of background color-matching in amphibians and reptiles. Ecology 1964, 45, 565–580. [Google Scholar] [CrossRef]
- Harris, A.C.; Weatherall, I.L. Geographic variation for colour in the sandburrowing beetle Chaerodes trachyscelides White (Coleoptera: Tenebrionidae) on New Zealand beaches analysed using CIELAB L values. Biol. J. Linn. Soc. 1991, 44, 93–104. [Google Scholar] [CrossRef]
- Stuart-Fox, D.; Moussalli, A.; Johnston, G.R.; Owens, I.P.F. Evolution of color variation in dragon lizards: Quantitative tests of the role of crypsis and local adaptation. Evolution 2004, 58, 1549–1559. [Google Scholar] [CrossRef]
- Hoekstra, H.E.; Krenz, J.G.; Nachman, M.W. Local adaptation in the rock pocket mouse (Chaetodipus intermedius): Natural selection and phylogenetic history of populations. Heredity 2005, 94, 217–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenblum, E.B. Convergent evolution and divergent selection: Lizards at the White Sands ecotone. Am. Nat. 2006, 167, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Marshall, K.L.A.; Philpot, K.E.; Damas-Moreira, I.; Stevens, M. Intraspecific colour variation among lizards in distinct island environments enhances local camouflage. PLoS ONE 2015, 10, e0135241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cadena, V.; Smith, K.R.; Endler, J.A.; Stuart-Fox, D. Geographic divergence and colour change in response to visual backgrounds and illumination intensity in bearded dragons. J. Exp. Biol. 2017, 220, 1048–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Hanlon, A.; Feeney, K.; Dockery, P.; Gormally, M.J. Quantifying phenotype-environment matching in the protected Kerry spotted slug (Mollusca: Gastropoda) using digital photography: Exposure to UV radiation determines cryptic colour morphs. Front. Zool. 2017, 14, 319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, H.; Li, J.; Wo, Y.; Shao, G.; Zhao, W.; Aguilar-Gómez, D.; Jin, Y. Effects of substrate color on intraspecific body color variation in the toad-headed lizard, Phrynocephalus versicolor. Ecol. Evol. 2019, 9, 10253–10262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endler, J.A. A predator’s view of animal color patterns. Evol. Biol. 1978, 11, 319–364. [Google Scholar]
- Nachman, M.W.; Hoekstra, H.E.; D’Agostino, S.L. The genetic basis of adaptive melanism in pocket mice. Proc. Natl. Acad. Sci. USA 2003, 100, 5268–5273. [Google Scholar] [CrossRef] [Green Version]
- Laurent, S.; Pfeifer, S.P.; Settles, M.L.; Hunter, S.S.; Hardwick, K.M.; Ormond, L.; Sousa, V.C.; Jensen, J.D.; Rosenblum, E.B. The population genomics of rapid adaptation: Disentangling signatures of selection and demography in white sands lizards. Mol. Ecol. 2016, 25, 306–323. [Google Scholar] [CrossRef]
- Körner, C. The use of “altitude” in ecological research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef]
- Keller, I.; Alexander, J.M.; Holderegger, R.; Edwards, P.J. Widespread phenotypic and genetic divergence along altitudinal gradients in animals. J. Evol. Biol. 2013, 26, 2527–2543. [Google Scholar] [CrossRef] [PubMed]
- Endler, J.A. Natural selection on colour patterns in Poecilia reticulata. Evolution 1980, 34, 76–91. [Google Scholar] [CrossRef] [PubMed]
- Camacho, L.; Avilés, L. Decreasing predator density and activity explain declining predation of insect prey along elevational gradients. Am. Nat. 2019, 194, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Cloudsley-Thompson, J.L. Multiple factors in the evolution of animal colouration. Naturwissenschaften 1999, 86, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Clusella-Trullas, S.; van Wyk, J.H.; Spotila, J.R. Thermal melanism in ectotherms. J. Thermal. Biol. 2007, 32, 235–245. [Google Scholar] [CrossRef]
- Ulbing, C.K.S.; Muuse, J.M.; Miner, B.E. Melanism protects alpine zooplankton from DNA damage caused by ultraviolet radiation. Proc. R. Soc. B 2019, 286, 20192075. [Google Scholar] [CrossRef]
- Marshall, K.L.A.; Stevens, M. Wall lizards display conspicuous signals to conspecifics and reduce detection by avian predators. Behav. Ecol. 2014, 25, 1325–1337. [Google Scholar] [CrossRef] [Green Version]
- Huey, R.B.; Slatkin, M. Cost and benefits of lizard thermoregulation. Q. Rev. Biol. 1976, 51, 363–384. [Google Scholar] [CrossRef]
- Orton, R.W.; McBrayer, L.D. Resolving tradeoffs among crypsis, escape behavior, and microhabitat use in sexually dichromatic species. Oecologia 2019, 189, 91–104. [Google Scholar] [CrossRef]
- Reguera, S.; Zamora-Camacho, F.J.; Moreno-Rueda, G. The lizard Psammodromus algirus (Squamata: Lacertidae) is darker at high altitudes. Biol. J. Linn. Soc. 2014, 112, 132–141. [Google Scholar] [CrossRef] [Green Version]
- Salvador, A. Lagartija Colilarga—Psammodromus Algirus (Linnaeus, 1758). In Enciclopedia Virtual de los Vertebrados Españoles; Salvador, A., Marco, A., Eds.; Museo Nacional de Ciencias Naturales: Madrid, Spain, 2015. [Google Scholar]
- Díaz, J.A.; Carrascal, L.M. Regional distribution of a Mediterranean lizard: Influence of habitat cues and prey abundance. J. Biogeogr. 1991, 18, 291–297. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Rueda, G.; Melero, E.; Reguera, S.; Zamora-Camacho, F.J.; Álvarez-Benito, I. Prey availability, prey selection, and trophic niche width in the lizard Psammodromus algirus along an elevational gradient. Curr. Zool. 2018, 64, 603–613. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Camacho, F.J.; Reguera, S.; Moreno-Rueda, G. Thermoregulation in the lizard Psammodromus algirus along a 2200-m elevational gradient in Sierra Nevada (Spain). Int. J. Biometeorol. 2016, 60, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Díaz, J.A. Breeding coloration, mating opportunities, activity, and survival in the lacertid lizard Psammodromus algirus. Can. J. Zool. 1993, 71, 1104–1110. [Google Scholar] [CrossRef]
- Martín, J.; López, P. Amphibians and reptiles as prey of birds in Southwestern Europe. Herpetol. Inf. Serv. 1990, 82, 1–43. [Google Scholar]
- Martín, J.; Avery, R.A. Effects of tail loss on the movement patterns of the lizard, Psammodromus algirus. Funct. Ecol. 1998, 12, 794–802. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Tris, J.; Díaz, J.A.; Tellería, J.L. Loss of body mass under predation risk: Cost of antipredatory behaviour or adaptive fit-for-escape? Anim. Behav. 2004, 67, 511–521. [Google Scholar] [CrossRef]
- López, P.; Martín, J. Effects of microhabitat-dependent predation risk on vigilance during intermittent locomotion in Psammodromus algirus lizards. Ethology 2013, 119, 316–324. [Google Scholar] [CrossRef]
- Díaz, J.A. Choice of compass directions around shrub patches by the heliothermic lizard Psammodromus algirus. Herpetologica 1992, 48, 293–300. [Google Scholar]
- Martín, J.; López, P. Influence of habitat structure on the escape tactics of the lizard Psammodromus algirus. Can. J. Zool. 1995, 73, 129–132. [Google Scholar] [CrossRef]
- Martín, J.; López, P. Fleeing to unsafe refuges: Effects of conspicuousness and refuge safety on the escape decisions of the lizard Psammodromus Algirus. Can. J. Zool. 2000, 78, 265–270. [Google Scholar] [CrossRef] [Green Version]
- Díaz, J.A.; Verdú-Ricoy, J.; Iraeta, P.; Llanos-Garrido, A.; Pérez-Rodríguez, A.; Salvador, A. There is more to the picture than meets the eye: Adaptation for crypsis blurs phylogeographical structure in a lizard. J. Biogeogr. 2017, 44, 397–408. [Google Scholar] [CrossRef] [Green Version]
- Zamora-Camacho, F.J.; Reguera, S.; Moreno-Rueda, G.; Pleguezuelos, J.M. Patterns of seasonal activity in a Mediterranean lizard along a 2200m altitudinal gradient. J. Thermal. Biol. 2013, 38, 64–69. [Google Scholar] [CrossRef]
- Carretero, M.A. Sources of colour pattern variation in Mediterranean Psammodromus algirus. Neth. J. Zool. 2002, 52, 43–60. [Google Scholar] [CrossRef] [Green Version]
- Montgomerie, R. Analazing Colors. In Bird Coloration. Vol. I: Mechanisms and Measurements; Hill, G.E., McGraw, K.J., Eds.; Harvard University Press: Cambridge, UK, 2006; pp. 90–147. [Google Scholar]
- Kemp, D.J.; Herberstein, M.E.; Fleishman, L.J.; Endler, J.A.; Bennett, A.T.D.; Dyer, A.G.; Hart, N.S.; Marshall, J.; Whiting, M.J. An integrative framework for the appraisal of coloration in nature. Am. Nat. 2015, 185, 705–724. [Google Scholar] [CrossRef] [Green Version]
- Stevens, M.; Párraga, C.A.; Cuthill, I.C.; Partridge, J.C.; Troscianko, T.S. Using digital photography to study animal coloration. Biol. J. Linn. Soc. 2007, 90, 211–237. [Google Scholar] [CrossRef] [Green Version]
- Cuthill, I.C.; Allen, W.L.; Arbuckle, K.; Caspers, B.; Chaplin, G.; Hauber, M.E.; Hill, G.E.; Jablonski, N.G.; Jiggins, C.D.; Kelber, A.; et al. The biology of color. Science 2017, 357. [Google Scholar] [CrossRef] [Green Version]
- Endler, J.A. Frequency-dependent predation, crypsis and aposematic coloration. Phil. Trans. R. Soc. B 1988, 319, 505–523. [Google Scholar] [CrossRef]
- Coulson, K.L.; Reynolds, D.W. The spectral reflectance of natural surfaces. J. Appl. Meteorol. 1971, 10, 1285–1295. [Google Scholar] [CrossRef] [Green Version]
- Eck, T.F.; Bhartia, P.K.; Hwang, P.H.; Stowe, L.L. Reflectivity of Earth’s surface and clouds in ultraviolet from satellite observations. J. Geophys. Res. Atmos. 1987, 92, 4287–4296. [Google Scholar] [CrossRef]
- Bergeron, Z.T.; Fuller, R.C. Using human vision to detect variation in avian coloration: How bad is it? Am. Nat. 2018, 191, 269–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karpestam, E.; Merilaita, S.; Forsman, A. Detection experiments with humans implicate visual predation as a driver of colour polymorphism dynamics in pygmy grasshoppers. BMC Ecol. 2013, 13, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Belliure, J.; Carrascal, L.M. Influence of heat transmission mode on heating rates and on the selection of patches for heating in a Mediterranean lizard. Physiol. Biochem. Zool. 2002, 75, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Díaz, J.A. Temporal patterns of basking behaviour in a Mediterranean lacertid lizard. Behaviour 1991, 118, 1–14. [Google Scholar] [CrossRef]
- Díaz, J.A.; Cabezas-Díaz, S. Seasonal variation in the contribution of different behavioural mechanisms to lizard thermoregulation. Funct. Ecol. 2004, 18, 867–875. [Google Scholar] [CrossRef]
- Zamora-Camacho, F.J.; Rubiño-Hispán, M.V.; Reguera, S.; Moreno-Rueda, G. Thermal dependence of sprint performance in the lizard Psammodromus algirus along a 2200-meter elevational gradient: Cold-habitat lizards do not perform better at low temperatures. J. Thermal. Biol. 2015, 52, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Merilaita, S.; Tuomi, J.; Jormalainen, V. Optimization of cryptic coloration in heterogeneous habitats. Biol. J. Linn. Soc. 1999, 67, 151–161. [Google Scholar] [CrossRef]
- Houston, A.I.; Stevens, M.; Cuthill, I.C. Animal camouflage: Compromise or specialize in a 2 patch-type environment? Behav. Ecol. 2007, 18, 769–775. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, J.; Ripa, J. The origin of polymorphic crypsis in a heterogeneous environment. Evolution 2010, 64, 1386–1394. [Google Scholar] [CrossRef]
- Hughes, A.; Liggins, E.; Stevens, M. Imperfect camouflage: How to hide in a variable world? Proc. R. Soc. B 2019, 286, 20190646. [Google Scholar] [CrossRef] [Green Version]
- Kawecki, T.J.; Ebert, D. Conceptual issues in local adaptation. Ecol. Lett. 2004, 7, 1225–1241. [Google Scholar] [CrossRef] [Green Version]
- Fox, S.F.; Perea-Fox, S.; Castro Franco, R. Development of the tail autotomy adaptation in lizards under disparate levels of predation at high and low elevations in Mexico. Southwest. Nat. 1994, 39, 311–322. [Google Scholar]





| Predictors | d.f. |
Lightness F-Value (p-Value) | Chroma F-Value (p-Value) | Hue F-Value (p-Value) |
|---|---|---|---|---|
| Bare soil | ||||
| Elevation | 5, 88 | 8.10 (<0.001) | 3.01 (0.015) | 6.82 (<0.001) |
| Month | 2, 88 | 1.76 (0.18) | 1.74 (0.18) | 0.57 (0.57) |
| Rock | ||||
| Elevation | 5, 48 | 1.88 (0.11) | 1.53 (0.20) | 0.33 (0.89) |
| Month | 2, 48 | 0.73 (0.49) | 1.27 (0.29) | 0.13 (0.88) |
| Fresh grass | ||||
| Elevation | 5, 87 | 1.26 (0.29) | 0.68 (0.64) | 2.46 (0.039) |
| Month | 2, 87 | 1.20 (0.31) | 2.68 (0.07) | 5.54 (0.004) |
| Dry grass | ||||
| Elevation | 5, 104 | 1.01 (0.41) | 0.82 (0.54) | 0.99 (0.42) |
| Month | 2, 104 | 0.50 (0.61) | 3.42 (0.036) | 0.87 (0.42) |
| Predictors | d.f. | Lightness F-Value (p-Value) | Chroma F-Value (p-Value) | Hue F-Value (p-Value) |
|---|---|---|---|---|
| Elevation | 5, 389 | 11.36 (<0.001) | 18.89 (<0.001) | 4.90 (<0.001) |
| Month | 5, 389 | 2.83 (0.016) | 4.76 (<0.001) | 1.43 (0.21) |
| Year | 2, 389 | 1.42 (0.24) | 2.87 (0.06) | 19.21 (<0.001) |
| Sex | 1, 389 | 1.60 (0.21) | 6.88 (0.009) | 3.08 (0.08) |
| Background | Lizard Dorsum | ||
|---|---|---|---|
| Lightness | Chroma | Hue Angle | |
| Bare soil | 0.71 (0.11) | −0.37 (0.47) | 0.94 (0.0048) |
| Rock | 0.71 (0.11) | −0.26 (0.62) | −0.17 (0.87) |
| Fresh grass | −0.60 (0.21) | −0.77 (0.07) | 0.77 (0.07) |
| Dry grass | 0.26 (0.62) | −0.03 (0.96) | 0.43 (0.40) |
| Predictors | d.f. | Colour Difference with Bare Soil F-Value (p-Value) | Colour Difference with Rock F-Value (p-Value) | Colour Difference with Fresh Grass F-Value (p-Value) | Colour Difference with Dry Grass F-value (p-value) |
|---|---|---|---|---|---|
| Elevation | (5, 389) | 272.13 (<0.001) | 71.36 (<0.001) | 32.11 (<0.001) | 78.78 (<0.001) |
| Month | (5, 389) | 1.12 (0.35) | 4.13 (0.001) | 1.18 (0.32) | 4.54 (0.0005) |
| Year | (2, 389) | 2.63 (0.07) | 4.04 (0.018) | 1.13 (0.32) | 2.70 (0.07) |
| Sex | (1, 389) | 0.29 (0.59) | 0.18 (0.67) | 4.05 (0.04) | 0.11 (0.74) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno-Rueda, G.; González-Granda, L.G.; Reguera, S.; Zamora-Camacho, F.J.; Melero, E. Crypsis Decreases with Elevation in a Lizard. Diversity 2019, 11, 236. https://doi.org/10.3390/d11120236
Moreno-Rueda G, González-Granda LG, Reguera S, Zamora-Camacho FJ, Melero E. Crypsis Decreases with Elevation in a Lizard. Diversity. 2019; 11(12):236. https://doi.org/10.3390/d11120236
Chicago/Turabian StyleMoreno-Rueda, Gregorio, Laureano G. González-Granda, Senda Reguera, Francisco J. Zamora-Camacho, and Elena Melero. 2019. "Crypsis Decreases with Elevation in a Lizard" Diversity 11, no. 12: 236. https://doi.org/10.3390/d11120236
APA StyleMoreno-Rueda, G., González-Granda, L. G., Reguera, S., Zamora-Camacho, F. J., & Melero, E. (2019). Crypsis Decreases with Elevation in a Lizard. Diversity, 11(12), 236. https://doi.org/10.3390/d11120236