Boreal Sand Hills are Areas of High Diversity for Boreal Ants (Hymenoptera: Formicidae)



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Research Plots and Vegetation Physiognomies
2.3. Specimen Sampling and Identification
2.4. Statistical Analysis
3. Results
3.1. Sampling Comparison
3.2. Diversity Comparisons
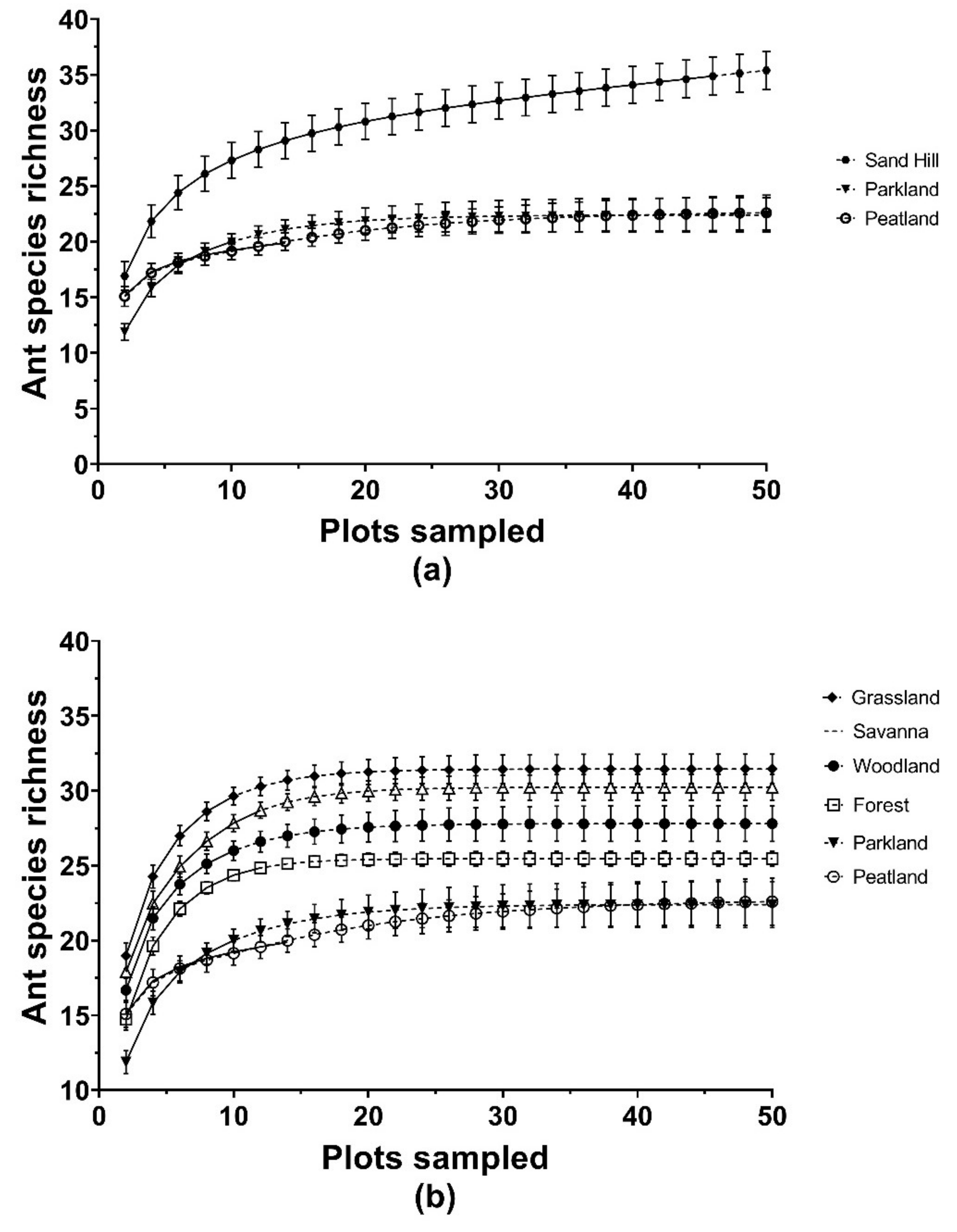
3.3. Species Accumulation and Species Rank Curves
4. Discussion
4.1. Diversity on Sand Hills
4.2. Notable Species Records
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Study Area | Latitude/Longitude | Area Type | Vegetation | Date of Recent Fires | Disturbance |
|---|---|---|---|---|---|
| Waskahegan Natural Area | 53°30′21.96″ N 112°56′8.81″ W | Aspen Parkland | Aspen Parkland (mixed Aspen and Black Spruce) | Has not burned recently | Grazed by cattle |
| Fort McMurray Peatland Forest Area | 56°25′43.59″ N 111° 4′49.89″ W | Peatland Forest | Black Spruce and Larch | May 2016 | Cut lines |
| North Bruderheim Natural Area | 53°52′8.54″ N 112°56′40.10″ W | Sand Hills | Jack Pine Forest and mixed aspen/jack pine forest | May 2009 | All terrain vehicle and petroleum industry |
| Northwest Bruderheim Natural Area | 53°52′8.54″ N 112°56′40.10″ W | Sand Hills | Jack Pine Forest and mixed aspen/jack pine/ forest | May 2009 | All terrain vehicle and petroleum industry |
| Opal Natural Area | 53°59′13.59″ N 113°18′34.96″ W | Sand Hills | Jack Pine Forest and mixed aspen/jack pine/black spruce forest | May 2010 | All terrain vehicle |
| Redwater Natural Area | 53°56′27.66″ N 112°57′17.19″ W | Sand Hills | Jack Pine Forest and mixed aspen/jack pine forest | Has not burned recently | All terrain vehicle and petroleum industry |
| Woodbend Forest | 53°23′31.66″ N 113°45′15.38″ W | Sand Hills | Jack Pine Forest and mixed aspen/jack pine/black spruce forest | Has not burned recently | Petroleum industry |
| Vegetation Type in Sand Hills | Description | Canopy Cover |
|---|---|---|
| Grassland | Areas of open canopy often located on the tops of dunes. Open sand patches are common. Lichen, sedges, and small shrubs such as roses or pin cherry make up common ground cover. | 0–5% |
| Savannah | Openings with a few jack pine and aspen. Often dominated by lichen, with bryophytes in shaded areas. Sedges and grasses are common, with scattered shrubs such as saskatoons, pin cherry and roses. | >5–25% |
| Woodland | Jack pine and aspen with mix of lichen and bryophyte ground cover. Roses and saskatoons are common. Grasses and sedges are uncommon | >25–60% |
| Forest | Jack pine forest interspersed with rare aspen. Ground cover dominated by bryophytes. Sometimes thick shrubs, sometimes none. | >60–100% |
References
- Holldöbler, B.; Wilson, E.O. The Ants; The Belknap Press of Harvard University Press: Cambridge, MA, USA, 1990; pp. 1–746. [Google Scholar]
- Handel, S.H.; Finch, S.B.; Schatz, G.E. Ants disperse a majority of herbs in a mesic forest community in New York State. Bull. Torrey Bot. Club 1981, 108, 430–437. [Google Scholar] [CrossRef]
- Briese, D. The effects of ants on the soil of a semi-arid saltbush habitat. Insectes Sociaux 1982, 29, 375–386. [Google Scholar] [CrossRef]
- Folgarait, P. Ant biodiversity and its relationship to ecosystem functioning: A review. Biodivers. Conserv. 1998, 7, 1221–1244. [Google Scholar] [CrossRef]
- Acorn, J.H. Sand Hill Arthropods in Canadian Grasslands. In Arthropods of Canadian Grasslands: Inhabitants of a Changing Landscape; Floate, K.D., Ed.; Biological Survey of Canada: Ottawa, ON, Canada, 2011; Volume 2, pp. 25–43. [Google Scholar]
- Wolfe, S.; Huntley, D.; Ollerhead, J. Relict Late Wisconsinan Dune Fields of the Northern Great Plains, Canada. Géogr. Phys. Quat. 2014, 58, 323–336. [Google Scholar] [CrossRef]
- Wolfe, S.A.; Huntley, D.J.; David, P.P.; Ollerhead, J.; Sauchyn, D.J.; McDonald, G.M. Late eighteenth century drought induced sand dune activity, Great Sand Hills, Saskatchewan. Can. J. Earth Sci. 2001, 38, 105–117. [Google Scholar] [CrossRef]
- Lewis, F.; Dowding, E.S. The vegetation of Alberta: II. the swamp, moor and bog forest vegetation of central Alberta. J. Ecol. 1928, 16, 19–70. [Google Scholar] [CrossRef]
- Downing, D.J.; Pettapiece, W.W. Natural Regions and Subregions of Alberta. Natural Regions Committee Government of Alberta. 2006. Available online: https://www.albertaparks.ca/media/2942026/nrsrcomplete_may_06.pdf (accessed on 13 November 2018).
- Howe, M.A.; Knight, G.T.; Clee, C. The importance of coastal dunes for terrestrial invertebrates in Wales and the UK, with particular reference to aculeate Hymenoptera (bees, wasps & ants). J. Coast. Conserv. 2010, 14, 91–102. [Google Scholar]
- Larsen, P. Spatial and temporal variations in boreal forest fire frequency in Northern Alberta. J. Biogeogr. 1997, 24, 663–673. [Google Scholar] [CrossRef]
- Galle, L. Structure and succession of ant assemblages in a north European sand dune area. Holarct. Ecol. 1991, 14, 31–37. [Google Scholar] [CrossRef]
- Galle, L.; Körmöczi, L.; Hornung, E.; Kerekes, J. Structure of ant assemblages in a Middle-European successional sand-dune area. Tiscia 1998, 31, 19–28. [Google Scholar]
- Boulton, A.M.; Davies, F.D.; Ward, P.S. Species richness, abundance and composition of ground-dwelling ants in northern California grassland: Role of plants, soil, and grazing. Environ. Entomol. 2005, 34, 96–104. [Google Scholar] [CrossRef]
- Palladini, J.D.; Jones, M.G.; Sanders, N.J.; Jules, E.S. The recovery of ant communities in regenerating temperate conifer forests. For. Ecol. Manag. 2007, 242, 619–624. [Google Scholar] [CrossRef]
- Lemon, P. A spherical densitometer for estimating forest overstory density. For. Sci. 1956, 2, 314–320. [Google Scholar]
- Glasier, J.R.N.G. Community Ecology of Ants (Hymenoptera: Formicidae) un the Central Sand Hills of Alberta, and a Key to the Ants of Alberta. Master’s Thesis, University of Alberta, Edmonton, AB, Canada, 2012. [Google Scholar]
- Weeks, R.D., Jr.; McIntyre, N.E. A comparison of live versus kill pitfall trapping techniques using various killing agents. Entomol. Exp. Appl. 1997, 82, 267–273. [Google Scholar] [CrossRef]
- Bestelmeyer, B.T.; Agosti, D.; Alonso, L.E.; Brandão, C.R.F.; Brown, W.L., Jr.; Delabie, J.H.C.; Silvestre, R. Field techniques for the study of ground-dwelling ants: An overview, description, and evaluation. In Ants Standard Methods for Measuring and Monitoring Biodiversity; Agosti, D., Majer, J.D., Alonso, L.E., Schultz, T., Eds.; Smithsonian Institution Press: Washington, DC, USA, 2000; pp. 122–144. [Google Scholar]
- Creighton, W.S. The ants of North America. Bull. Mus. Comp. Zool. 1950, 104, 1–585. [Google Scholar]
- Wheeler, G.C.; Wheeler, J. The Ants of North Dakota; University of North Dakota Press: Grand Forks, ND, USA, 1963; pp. 1–325. [Google Scholar]
- Francoeur, A. Revision taxonomique des espesce nearctiques du groupe Fusca, genre Formica (Formicidae, Hymenoptera). Mem. De La Soc. Entomogique Du Que. 1973, 3, 1–316. [Google Scholar]
- Mackay, W.; Mackay, E. The Ants of New Mexico (Hymenoptera: Formicidae); The Edwin Mellen Press: Lewiston, NY, USA, 2002; pp. 1–398. [Google Scholar]
- Hasen, L.D.; Klotz, J.H. The Carpenter Ants of the United States and Canada; Cornell University Press: Ithaca, NY, USA, 2005; pp. 1–204. [Google Scholar]
- Glasier, J.R.N.; Acorn, J.H.; Nielsen, S.; Proctor, H. Ants (Hymenoptera: Formicidae) of Alberta: A key to species based primarily on the worker caste. Can. J. Arthropod Identif. 2013, 22, 1–104. [Google Scholar]
- Colwell, R.K.; Estimate, S. Statistical Estimation of Species Richness and Shared Species from Samples, Version 8.2, User’s Guide and Application. 2009. Available online: http://purl.oclc.org/estimate (accessed on 12 August 2018).
- Heron, J. Ants of the south Okanagan grasslands, British Columbia. Arthropods Can. Grassl. 2005, 11, 17–22. [Google Scholar]
- Higgins, R.; Lindgren, S. Species List of Ants Collected in Central and North-Central British Columbia. 2005. Available online: http://web.unbc.ca/~lindgren/ants_species_list.html (accessed on 8 October 2018).
- Glasier, J.R.N.; Acorn, J.H. An Annotated List of Ants (Hymenoptera: Formicidae) from the Grasslands of Alberta and Saskatchewan. In Arthropods of Canadian Grasslands (Volume 4): Biodiversity and Systematics Part 2; Giberson, D.J., Cárcamo, H.A., Eds.; Biological Survey of Canada: Ottawa, ON, Canada, 2014; pp. 299–314. [Google Scholar]
- Lessard, J.; Buddle, C.M. The effects of urbanization on ant assemblages (Hymenoptera: Formicidae) associated with the Molson nature reserve, Quebec. Can. Entomol. 2005, 137, 215–225. [Google Scholar] [CrossRef]
- Lassau, S.A.; Hochuli, D.F. Effects of habitat complexity on ant assemblages. Ecography 2004, 27, 157–164. [Google Scholar] [CrossRef]
- Delabie, J.H.C.; Fisher, B.L.; Majer, J.D.; Wright, I.W. Sampling effort and choice of methods. In Ants Standard Methods for Measuring and Monitoring Biodiversity; Agosti, D., Majer, J.D., Alonso, L.E., Schultz, T., Eds.; Smithsonian Institution Press: Washington, DC, USA, 2000; pp. 122–144. [Google Scholar]
- Mackay, W. A review of the New World ants of the genus Dolichoderus (Hymenoptera: Formicidae). Sociobiology 1993, 22, 1–148. [Google Scholar]
- Francoeur, A.; Buschinger, A. Biosystematiwue de la tribu Leptothoacini (Formicidae, Hymenoptera) 1. Le genera Formicoxenus dans la region Holarctique. Nat. Can. 1985, 112, 343–403. [Google Scholar]
- Social Insects Specialist Group. Harpagoxenus canadensis. In IUCN 2011. IUCN Red List of Threatened Species, Version 2011.1. 1996. Available online: www.iucnredlist.org (accessed on 7 December 2018).
- Stuart, R.J.; Alloway, T.M. The slave-making ant, Harpagoxenus canadensis, M. R. Smith, and its host-species, Leptothorax muscorum (Nylander): Slave raiding and territoriality. Behaviour 1983, 85, 58–90. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D.N. Barcoding, BOLD: The barcode of life data system (www.barcodinglife.org). NPR Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Francoeur, A. Les fourmis de la forêt boréale du Québec. Nat. Can. 2001, 125, 108–114. [Google Scholar]



| Species | Aspen Parkland | Peatland Forest | Sand Hill Grassland | Sand Hill Savannah | Sand Hill Woodland | Sand Hill Forest |
|---|---|---|---|---|---|---|
| Camponotus herculeanus | x | x | x | x | x | x |
| Camponotus nearcticus | x | x | ||||
| Camponotus novaeboracensis | x | x | x | x | x | x |
| Dolichoderus taschenbergi | x | x | x | x | ||
| Formica accreta | x | x | x | x | x | x |
| Formica adamsi | x | x | x | x | ||
| Formica aserva | x | x | x | x | x | x |
| Formica dakotensis | x | x | x | x | x | |
| Formica densiventris | x | x | x | x | x | |
| Formica hewitti | x | x | x | x | x | |
| Formica impexa | x | x | ||||
| Formica lasioides | x | x | x | x | x | |
| Formica neorufibarbis | x | x | x | x | x | x |
| Formica obscuriventris | x | x | x | x | x | |
| Formica oreas | x | x | x | x | ||
| Formica podzolica | x | x | x | x | x | x |
| Formica subintegra | x | |||||
| Formica ulkei | x | x | x | x | x | |
| Formicoxenus hirticornis | x | |||||
| Formicoxenus quebecensis | x | x | x | |||
| Harpagoxenus canadensis | x | |||||
| Lasius americanus | x | |||||
| Lasius aphidicolus | x | x | ||||
| Lasius crypticus | x | x | ||||
| Lasius neoniger | x | x | x | x | ||
| Lasius pallitarsis | x | x | x | x | x | |
| Leptothorax canadensis | x | x | x | x | x | x |
| Myrmica ab001 | x | x | x | x | x | |
| Myrmica alaskensis | x | x | x | x | x | x |
| Myrmica brevispinosa | x | x | x | x | ||
| Myrmica detritinodis | x | x | x | x | x | x |
| Myrmica fracticornis | x | x | x | x | x | x |
| Myrmica incompleta | x | |||||
| Myrmica lampra | x | |||||
| Myrmica lobifrons | x | |||||
| Myrmica nearctica | x | x | x | x | ||
| Myrmica quebecensis | x | |||||
| Polyergus mexicanus | x | x | x | |||
| Tapinoma sessile | x | x | x | x | ||
| Total Genera | 7 | 5 | 8 | 8 | 7 | 8 |
| Total Species | 20 | 20 | 30 | 29 | 26 | 25 |
| Ecosystem | Species (Generic) Richness | MMMeans | Fisher’s Alpha | Shannon Index | Simpson Diversity Index |
|---|---|---|---|---|---|
| Sand Hills | |||||
| Overall | 35 (10) | 34.91 | 3.99 (0.21) | 2.58 (0) | 9.98 |
| Grassland | 30 (8) | 33.94 | 4.07 (0.26) | 2.52 (0) | 9.31 |
| Savannah | 29 (8) | 31.90 | 3.67 (0.23) | 2.30 (0.12) | 6.88 |
| Woodland | 26 (8) | 30.12 | 3.48 (0.23) | 2.27 (0) | 6.70 |
| Forest | 25 (8) | 28.97 | 3.64 (0.27) | 2.15 (0) | 5.94 |
| Aspen Parkland | 20 (7) | 23.87 | 3.07 (0.26) | 1.69 (0.05) | 5.45 |
| Peatland Forest | 20 (5) | 20.71 | 2.42 (0.18) | 1.85 (0) | 4.64 |
| Locality | Ecosystem | Ant Species (Genera) Richness | Author(s) |
|---|---|---|---|
| Central Alberta Sand Hills, Alberta, Canada | Pine Barrens | 35 (10) | This Paper |
| Waskahegan, Alberta, Canada | Aspen Parkland | 20 (8) | This Paper |
| Fort McMurray, Alberta, Canada | Peatland Forest | 20 (6) | This Paper |
| Southern Okanagan Grassland, British Columbia, Canada | Brush Shrub Steppe | 31 (13) | Heron 2001 [27] |
| Prince George, British Columbia, Canada | Boreal Forest | 23 (10) | Higgins and Lindgren 2005 [28] |
| Great Sand Hills, Saskatchewan, Canada | Open Dunes Grassland | 20 (6) | Glasier and Acorn 2014 [29] Personal Collection |
| Molson Reserve, Quebec, Canada | Maple-Beech Forest | 24 (12) | Lessard and Buddle 2005 [30] |
| Hanko Peninsula, Finland | Pine Barrens | 24 (7) | Galle 1991 [12] |
| Kampinos National Park, Poland | Pine Barrens | 22(7) | Galle et al. 1998 [13] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glasier, J.R.N.; Nielsen, S.E.; Acorn, J.; Pinzon, J. Boreal Sand Hills are Areas of High Diversity for Boreal Ants (Hymenoptera: Formicidae). Diversity 2019, 11, 22. https://doi.org/10.3390/d11020022
Glasier JRN, Nielsen SE, Acorn J, Pinzon J. Boreal Sand Hills are Areas of High Diversity for Boreal Ants (Hymenoptera: Formicidae). Diversity. 2019; 11(2):22. https://doi.org/10.3390/d11020022
Chicago/Turabian StyleGlasier, James R.N., Scott E. Nielsen, John Acorn, and Jaime Pinzon. 2019. "Boreal Sand Hills are Areas of High Diversity for Boreal Ants (Hymenoptera: Formicidae)" Diversity 11, no. 2: 22. https://doi.org/10.3390/d11020022
APA StyleGlasier, J. R. N., Nielsen, S. E., Acorn, J., & Pinzon, J. (2019). Boreal Sand Hills are Areas of High Diversity for Boreal Ants (Hymenoptera: Formicidae). Diversity, 11(2), 22. https://doi.org/10.3390/d11020022