Geomorphology and Altitude Effects on the Diversity and Structure of the Vanishing Montane Forest of Southern Ecuador

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
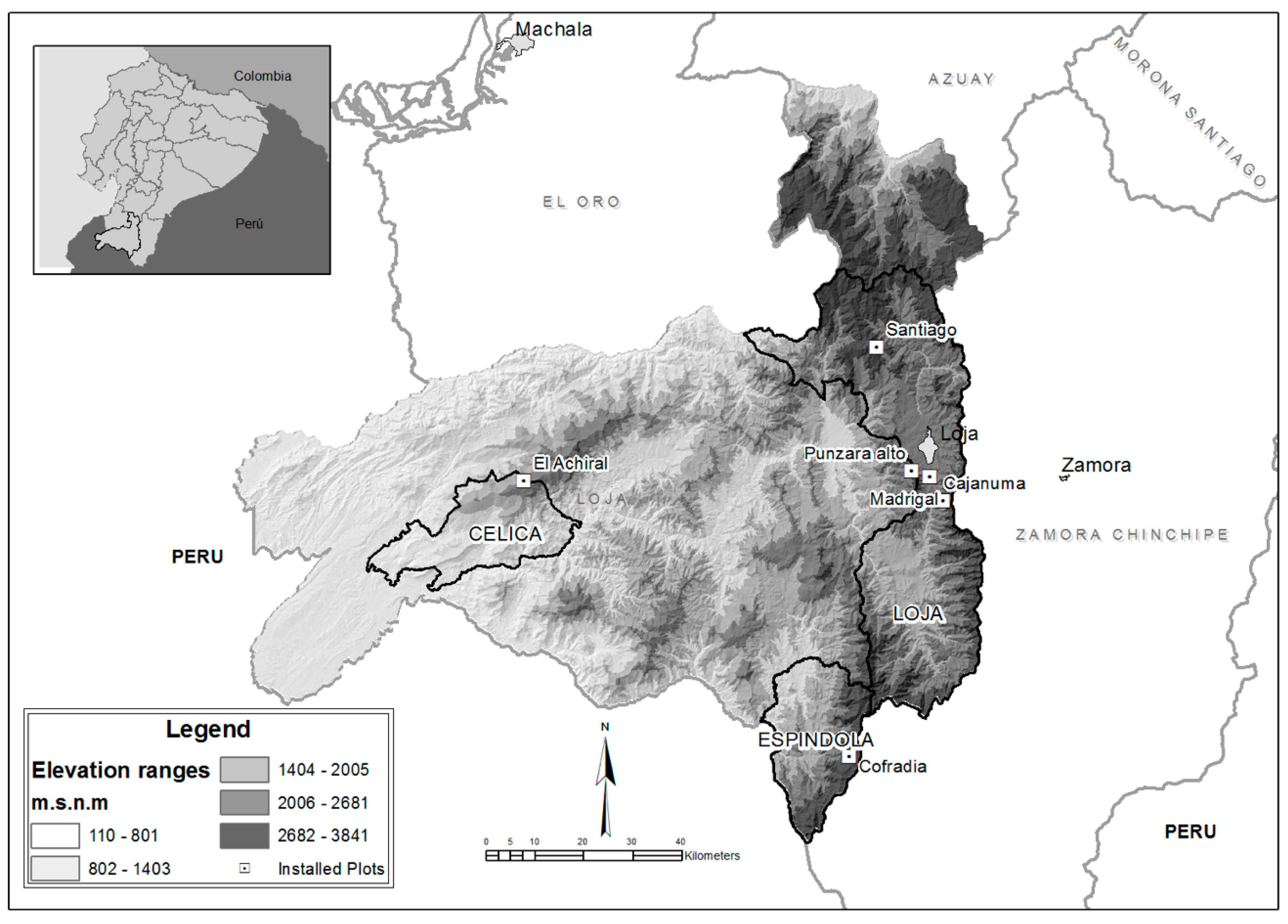
2.1. Study Area
2.2. Sampling Design
2.3. Statistical Analysis
2.3.1. Taxonomic Diversity
2.3.2. Composition
3. Results
3.1. Alpha Diversity
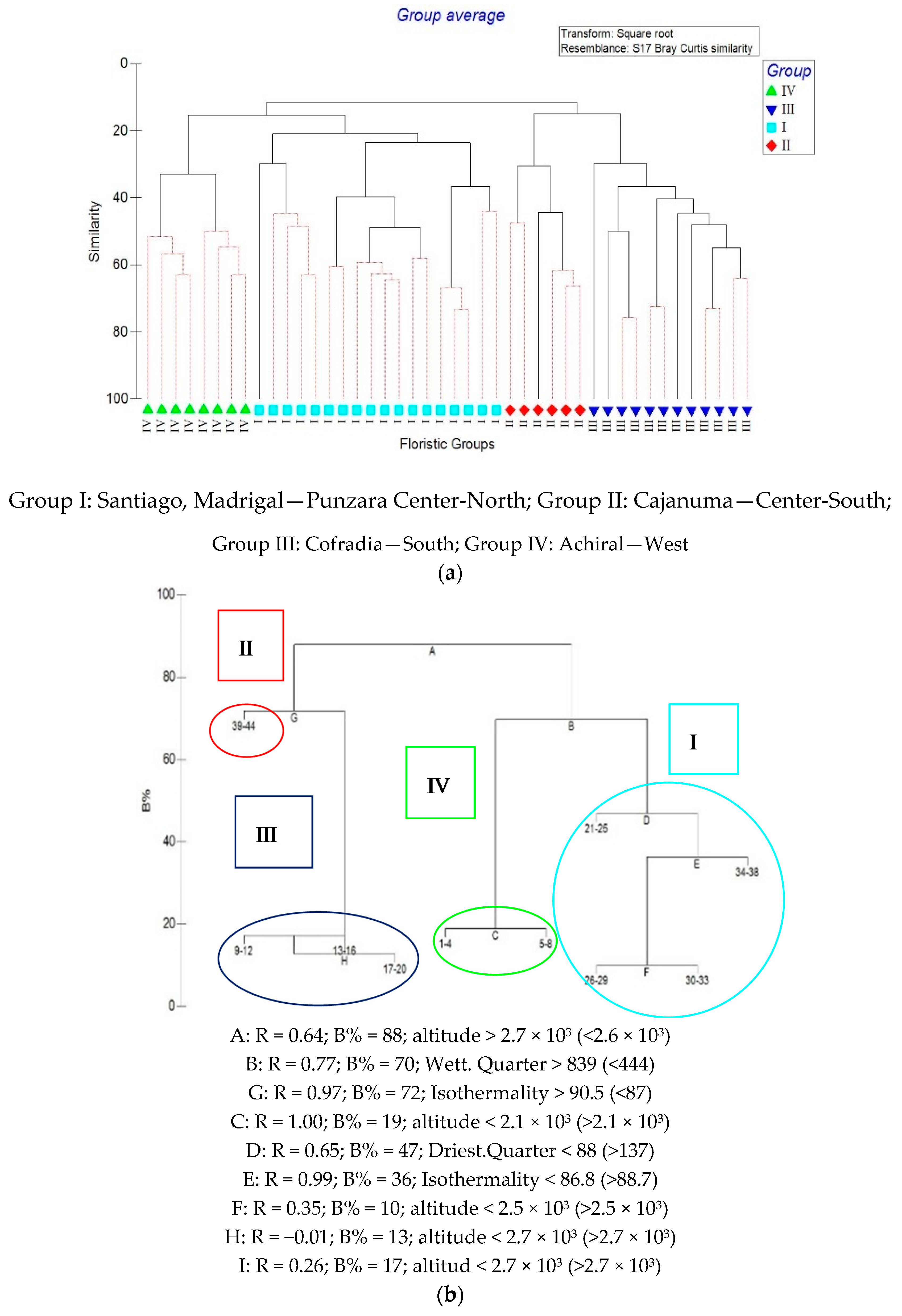
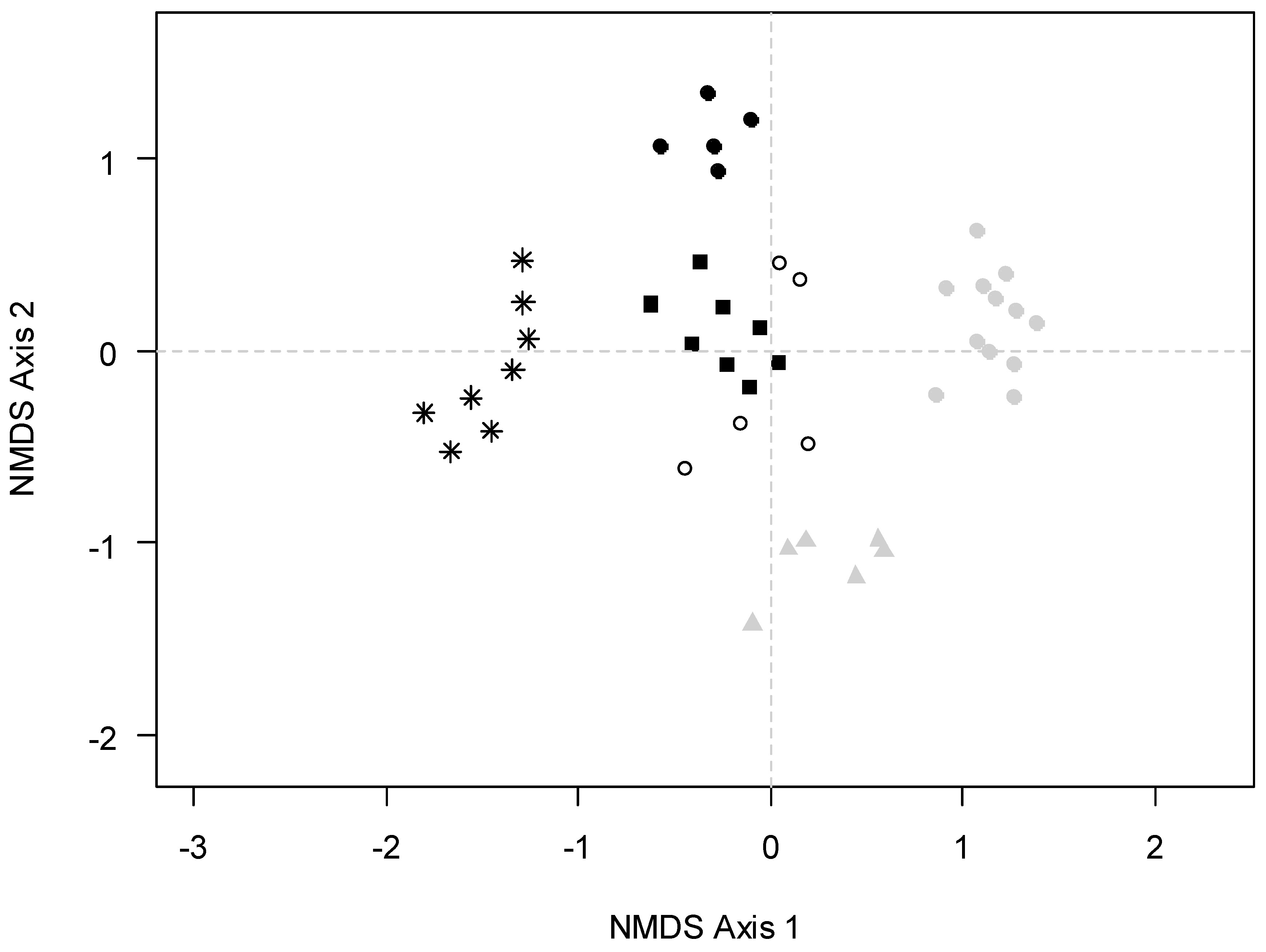
3.2. Species Turnover
3.3. Modeling the Diversity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da-Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.D.; Kappelle, M. Introducción a los bosques nublados del neotrópico: Una síntesis regional. In Bosques nublados del Neotrópico; Kappelle, M., Brown, A.D., Eds.; Instituto Nacional de Biodiversidad (INBio): Santo Domingo de Heredia, Costa Rica, 2001; pp. 27–40. ISBN 9968-702-50-1. [Google Scholar]
- Olson, D.M.; Dinerstein, E. The Global 200: Priority ecoregions for global conservation. Ann. Mo. Bot. Gard. 2002, 89, 199–224. [Google Scholar] [CrossRef]
- Mena-Vásconez, P.M. Las áreas protegidas con bosque montano en el Ecuador. In Biodiversity and Conservation of Neotropical Montane Forests; Churchill, P., Balslev, H., Forero, E., Luteyn, J.L., Eds.; The New York Botanical Garden: New York, NY, USA, 1995; pp. 627–635. ISBN 0893273902. [Google Scholar]
- Valencia, R. Composition and structure of an Andean forest fragment in Eastern Ecuador. In Biodiversity and Conservation of Neotropical Montane Forests; Churchill, P., Balslev, H., Forero, E., Luteyn, J.L., Eds.; The New York Botanical Garden: New York, NY, USA, 1995; pp. 239–249. ISBN 0893273902. [Google Scholar]
- Tapia-Armijos, M.F.; Homeier, J.; Espinosa, C.I.; Leuschner, C.; de la Cruz, M. Deforestation and forest fragmentation in South Ecuador since the 1970s–losing a hotspot of biodiversity. PLoS ONE 2015, 10, e0133701. [Google Scholar] [CrossRef] [PubMed]
- Uday, V.; Bussman, R. Floristic distribution of the montane cloud forest at the Tapichalaca Reserve, Canton Palanda, Zamora province. Lyonia 2004, 7, 92–98. [Google Scholar]
- Madsen, J.E.; Øllgaard, B. Floristic composition, structure, and dynamics of an upper montane rain forest in Southern Ecuador. Nord. J. Bot. 1994, 14, 403–423. [Google Scholar] [CrossRef]
- Bussmann, R.W. The montane forests of Reserva Biologica San Francisco (Zamora-Chinchipe, Ecuador)-vegetation zonation and natural regeneration. Erde 2001, 132, 9–25. [Google Scholar]
- Van der Hammen, T. History of the montane forests of the northern Andes. Plant Syst. Evol. 1989, 162, 109–114. [Google Scholar] [CrossRef]
- Sodiro, L. Ojeada General Sobre la Vegetación Ecuatoriana; Universidad de Quito: Quito, Ecuador, 1874. [Google Scholar]
- Diels, L. Contribuciones al Conocimiento de la Flora y Vegetación de Ecuador; Espinosa, R., Ed.; Anales de la Universidad Central: Quito, Ecuador, 1937. [Google Scholar]
- Acosta-Solís, M. Divisiones Fitogeográficas y Formaciones Geobotánicas del Ecuador; Casa de la Cultura ecuatoriana: Quito, Ecuador, 1968. [Google Scholar]
- Harling, G. The vegetation types of Ecuador: A brief survey. In Tropical Botany; Larsen, K., Holm-Nielsen, L.B., Eds.; Academic Press: London, UK, 1979; pp. 164–174. ISBN 012437350X. [Google Scholar]
- Cañadas-Cruz, L.; Cogez, X.; Lyannaz, J.P.; Ammerman, C.B.; Henry, P.R.; Muñoz, K.A.; Wilson, P.N. El Mapa Bioclimático y Ecológico del Ecuador; Ministerio de Agricultura y Ganadería: Quito, Ecuador, 1983. [Google Scholar]
- Ulloa-Ulloa, C.; Jorgensen, P.M. Árboles y Arbustos de los Andes del Ecuador; Department of Systematic Botany: Aarhus, Denmark, 1993. [Google Scholar]
- Mosandl, R.; Günter, S.; Stimm, B.; Weber, M. Ecuador suffers the highest deforestation rate in South America. In Gradients in a Tropical Mountain Ecosystem of Ecuador; Beck, E., Bendix, J., Kottke, I., Makeschin, F., Mosandl, R., Eds.; Springer: Berlin, Germany, 2008; pp. 37–40. ISBN 9783540735267. [Google Scholar]
- Sierra, R. Propuesta Preliminar de un Sistema de Clasificación de Vegetación para el Ecuador Continental; EcoCiencia: Quito, Ecuador, 1999; ISBN 9789978409435. [Google Scholar]
- Bussmann, R.W. Bosques andinos del sur de Ecuador, clasificación, regeneración y uso. Rev. Peruana Biol. 2005, 12, 203–216. [Google Scholar] [CrossRef]
- Lozano, P. Los tipos de bosque en el sur de Ecuador. In Botánica Austroecuatoriana–Estudios Sobre los Recursos Vegetales en las Provincias de El Oro, Loja y Zamora Chinchipe; Aguirre, Z., Madsen, J.E., Cotton, E., Balslev, H., Eds.; Abya Yala: Quito, Ecuador, 2002; pp. 29–50. ISBN 9789978222515. [Google Scholar]
- Lozano, P.; Bussmann, R.W.; Küppers, M. Diversidad florística del bosque montano en el Occidente del Parque Nacional Podocarpus, Sur del Ecuador y su influencia en la flora pionera en deslizamientos naturales. Rev. Cient. UDO Agríc. 2007, 7, 142–159. [Google Scholar]
- Becking, M. Sistema Microregional de Conservación Podocarpus. Tejiendo (Micro) Corredores de Conservación Hacia la Cogestión de una Reserva de la Biosfera Podocarpus-El Cóndor; Programa Podocarpus: Loja, Ecuador, 2004. [Google Scholar]
- Lozano, P.; Busmann, R.W.; Küppers, M.; Lozano, D. Natural landslides and pioneer communities in the Mountain Ecosystems of Eastern Podocarpus National Park. Caldasia 2008, 30, 1–19. [Google Scholar] [CrossRef]
- Homeier, J.; Breckle, S. Gap-dynamics in a tropical lower montane forest in South Ecuador. In Gradients in a Tropical Mountain Ecosystem of Ecuador; Beck, E., Bendix, J., Kottke, I., Makeschin, F., Mosandl, R., Eds.; Springer: Berlin, Germany, 2008; pp. 311–317. ISBN 9783540735267. [Google Scholar]
- Jørgensen, P.M.; León-Yánez, S. Catálogo de las plantas vasculares del Ecuador. Monogr. Syst. Bot. Mo. Bot. Gard. 1999, 75, 1–1181. [Google Scholar]
- Ulloa, C.; Neill, D.A. Cinco Años de Adiciones a la Flora del Ecuador 1999–2004; Funbotanica; Universidad Técnica Particular de Loja: Loja, Ecuador, 2005. [Google Scholar]
- Magurran, A.E. Why diversity? In Ecological Diversity and Its Measurement; Springer: Dordrecht, The Netherlands, 1988; pp. 1–5. [Google Scholar]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef] [Green Version]
- Sabah, A.A.W.; Charles, S.B.; Salem, M.A.A. Principal component and multiple regression analysis in modeling of ground-level ozone and factors affecting its concentrations. Environ. Model. Softw. 2005, 20, 1263–1271. [Google Scholar] [CrossRef]
- Malinowski, R.M. Factor Analysis in Chemistry; Wiley: New York, NY, USA, 1991; ISBN 978-0-471-53009-1. [Google Scholar]
- Statheropoulos, M.; Vassiliadis, N.; Pappa, A. Principal component and canonical correlation analysis for examining air pollution and meteorological data. Atmos. Environ. 1998, 32, 1087–1095. [Google Scholar] [CrossRef]
- Cayuela, L.; Benayas, J.M.R.; Justel, A.; Salas-Rey, J. Modelling tree diversity in a highly fragmented tropical montane landscape. Glob. Ecol. Biogeogr. 2006, 15, 602–613. [Google Scholar] [CrossRef]
- R Development Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; ISBN 3-900051-07-0. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; Plymouth Routines in Multivariate Ecological Research: Plymouth, MA, USA; Primer-E Ltd.: Auckland, New Zealand, 2006. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Oksanen, M.J. Package ‘vegan’. Community Ecology Package. Version, 2(9). 2013. Available online: https://cran.r-project.org/web/packages/vegan/vegan.pdf (accessed on 12 October 2018).
- Josse, C.; Navarro, G.; Comer, P.; Evans, R.; Faber-Langendoen, D.; Fellows, M.; Kittel, G.; Menard, S.; Pyne, M.; Teague, J. Ecological Systems of Latin America and the Caribbean: A Working Classification of Terrestrial Systems; NatureServe: Arlington, VA, USA, 2003. [Google Scholar]
- Churchill, S.P.; Balslev, H.; Forero, E.; Luteyn, J.L. Biodiversity and Conservation of Neotropical Montane Forests; New York Botanical Garden: New York, NY, USA, 1995. [Google Scholar]
- Bush, M.B.; Hanselman, J.A.; Hooghiemstra, H. Andean montane forests and climate change. In Tropical Rainforest Response to Climatic Change; Bush, M.B., Flenley, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 59–79. ISBN 978-3-642-05383-2. [Google Scholar]
- Antonelli, A.; Sanmartín, I. Why are there so many plant species in the Neotropics? Taxon 2011, 60, 403–414. [Google Scholar] [CrossRef]
- Givnish, T.J. Altitudinal gradients in tropical forest composition, structure, and diversity in the Sierra de Manantlán. J. Ecol. 1998, 86, 999–1020. [Google Scholar]
- Homeier, J.; Breckle, S.W.; Günter, S.; Rollenbeck, R.T.; Leuschner, C. Tree Diversity, Forest Structure and Productivity along Altitudinal and Topographical Gradients in a Species-Rich Ecuadorian Montane Rain Forest. Biotropica 2010, 42, 140–148. [Google Scholar] [CrossRef]
- Rafiqpoor, D.; Kier, G.; Kreft, H. Global centers of vascular plant diversity. Nova Acta Leopoldina 2005, 92, 61–83. [Google Scholar]
- Condit, R.; Hubbell, S.P.; Foster, R.B. Mortality rates of 205 neotropical tree and shrub species and the impact of a severe drought. Ecol. Monogr. 1995, 65, 419–439. [Google Scholar] [CrossRef]
- Kessler, M. The impact of population processes on patterns of species richness: Lessons from elevational gradients. Basic Appl. Ecol. 2009, 10, 295–299. [Google Scholar] [CrossRef] [Green Version]
- Kessler, M.; Kluge, J.; Hemp, A.; Ohlemüller, R. A global comparative analysis of elevational species richness patterns of ferns. Glob. Ecol. Biogeogr. 2011, 20, 868–880. [Google Scholar] [CrossRef]
- Willig, M.R.; Kaufman, D.M.; Stevens, R.D. Latitudinal gradients of biodiversity: Pattern, process, scale, and synthesis. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 273–309. [Google Scholar] [CrossRef]
- Kreft, H.; Jetz, W. Global patterns and determinants of vascular plant diversity. Proc. Natl. Acad. Sci. Biol. 2007, 104, 5925–5930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCain, C.M. Elevational gradients in diversity of small mammals. Ecology 2005, 86, 366–372. [Google Scholar] [CrossRef]
- McCain, C.M. Could temperature and water availability drive elevational species richness patterns? A global case study for bats. Glob. Ecol. Biogeogr. 2007, 16, 1–13. [Google Scholar] [CrossRef]
- McCain, C.M. Global analysis of bird elevational diversity. Glob. Ecol. Biogeogr. 2009, 18, 346–360. [Google Scholar] [CrossRef]
- McCain, C.M. Global analysis of reptile elevational diversity. Glob. Ecol. Biogeogr. 2010, 4, 541–553. [Google Scholar] [CrossRef]
- Rahbek, C. The role of spatial scale and the perception of large-scale species-richness patterns. Ecol. Lett. 2005, 8, 224–239. [Google Scholar] [CrossRef]
- Jørgensen, P.M.; Ulloa-Ulloa, C.; Madsen, J.E.; Valencia, R. A floristic analysis of the high Andes of Ecuador. In Biodiversity and Conservation of Neotropical Montane Forests; Churchill, S.P., Balslev, H., Forero, E., Luteyn, J.L., Eds.; New York Bot. Gard: New York, NY, USA, 1995; pp. 221–238. [Google Scholar]
- Matteucci, S.D.; Colma, A. Metodología para el Estudio de la Vegetación; Secretaría General de la Organización de los Estados Americanos, Programa Regional de Desarrollo Científico y Tecnológico: Washington, DC, USA, 1982. [Google Scholar]
- Espinosa, C.I.; De la Cruz, M.; Luzuriaga, A.L.; Escudero, A. Bosques tropicales secos de la región Pacífico Ecuatorial: Diversidad, estructura, funcionamiento e implicaciones para la conservación. Ecosistemas 2012, 21, 1–2. [Google Scholar]
- Richter, M.; Moreira-Muñoz, A. Heterogeneidad climática y diversidad de la vegetación en el sur de Ecuador: Un método de fitoindicación. Rev. Peruana Biol. 2005, 12, 217–238. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Groups | Families | # Species | Relative Diversity |
|---|---|---|---|
| I | MELASTOMATACEAE | 10 | 9.01 |
| LAURACEAE | 8 | 7.21 | |
| ASTERACEAE | 6 | 5.41 | |
| CUNNONIACEAE | 6 | 5.41 | |
| RUBIACEAE | 6 | 5.41 | |
| II | AQUIFOLIACEAE | 6 | 9.38 |
| MELASTOMATACEAE | 6 | 9.38 | |
| CUNNONIACEAE | 5 | 7.81 | |
| LAURACEAE | 5 | 7.81 | |
| SYMPLOCACEAE | 5 | 7.81 | |
| III | AQUIFOLIACEAE | 6 | 9.38 |
| MELASTOMATACEAE | 6 | 9.38 | |
| CUNNONIACEAE | 5 | 7.81 | |
| LAURACEAE | 5 | 7.81 | |
| SYMPLOCACEAE | 5 | 7.81 | |
| IV | LAURACEAE | 6 | 11.11 |
| EUPHORBIACEAE | 5 | 9.26 | |
| MYRTACEAE | 4 | 7.41 | |
| ASTERACEAE | 3 | 5.56 | |
| MELASTOMATACEAE | 3 | 5.56 |
| Groups | Species | Basal Area (m2) | Relative Dominance |
|---|---|---|---|
| I | Schefflera ferruginea (Kunth) Harms | 4 | 8.1 |
| Critoniopsis pycnantha (Benth.) H. Rob. | 2.5 | 5.1 | |
| Oreopanax eriocephalus Harms | 2.2 | 4.5 | |
| Nectandra reticulata (Ruiz & Pav.) Mez | 2.2 | 4.5 | |
| Clusia alata Triana & Planch. | 1.6 | 3.3 | |
| II | Prumnopitys montana (Humb. & Bonpl.) Laub. | 1.8 | 8.6 |
| Persea ferruginea Kunth | 1.4 | 6.4 | |
| Clusia alata Triana & Planch. | 1.3 | 6.2 | |
| Gordonia fruticosa (Schrad.) H. Keng. | 1.3 | 6 | |
| Weinmannia ovata Cav. | 1.3 | 5.9 | |
| III | Podocarpus oleifolius D. Don ex Lamb. | 10.1 | 17.4 |
| Clusia alata Triana & Planch. | 4.1 | 7.1 | |
| Persea ferruginea Kunth | 3.7 | 6.3 | |
| Citronella sp. | 3.5 | 6 | |
| Podocarpus sprucei Parlatore | 3.4 | 5.9 | |
| IV | Guarea kunthiana A. Juss. | 2.6 | 8.8 |
| Miconia jahnii Pittier | 2.3 | 7.9 | |
| Endlicheria sp. | 2.1 | 7 | |
| Nectandra laurel Ness | 1.8 | 6.1 | |
| Myrcianthes discolor (Kunth) McVaugh. | 1.8 | 6 |
| Groups IV & III | Average Dissimilarity = 95.40 | |||||
|---|---|---|---|---|---|---|
| Group IV | Group III | |||||
| Species | Av.Abund | Av.Abund | Av.Diss | Diss/SD | Contrib% | Cum.% |
| Podocarpus oleifolius D. Don ex Lamb. | 0 | 3.06 | 3.65 | 2.98 | 3.83 | 3.83 |
| Miconia jahnii Pittier | 2.98 | 0.14 | 3.41 | 2.66 | 3.58 | 7.41 |
| Guarea kunthiana A. Juss. | 2.62 | 0 | 3.1 | 2.51 | 3.24 | 10.65 |
| Clusia latipes Triana & Planch. | 0 | 2.19 | 2.61 | 1.71 | 2.74 | 13.39 |
| Aniba muca (Ruiz&Pav) Mez | 2.25 | 0.22 | 2.59 | 1.4 | 2.72 | 16.11 |
| Groups IV & I | Average dissimilarity = 84.56 | |||||
| Group IV | Group I | |||||
| Guarea kunthiana A. Juss. | 2.62 | 0.21 | 2.85 | 1.96 | 3.37 | 3.37 |
| Miconia jahnii Pittier | 2.98 | 1.15 | 2.43 | 1.35 | 2.87 | 6.25 |
| Aniba muca (Ruiz&Pav) Mez | 2.25 | 0.57 | 2.39 | 1.27 | 2.82 | 9.07 |
| Cupania cinerea Poepp. | 1.72 | 0 | 2.13 | 0.88 | 2.52 | 11.6 |
| Nectandra laurel Ness | 1.65 | 0.9 | 1.87 | 1.09 | 2.21 | 13.8 |
| Groups III & I | Average dissimilarity = 86.22 | |||||
| Group III | Group I | |||||
| Podocarpus oleifolius D. Don ex Lamb. | 3.06 | 0.22 | 3.35 | 2.25 | 3.88 | 3.88 |
| Clusia latipes Triana & Planch. | 2.19 | 0.23 | 2.36 | 1.48 | 2.74 | 6.62 |
| Ilex andicola Loes | 2.09 | 0.11 | 2.35 | 1.61 | 2.72 | 9.34 |
| Podocarpus sprucei Parlatore | 1.98 | 0 | 2.28 | 2.15 | 2.64 | 11.99 |
| Citronella sp. | 1.93 | 0 | 2.18 | 1.18 | 2.53 | 14.51 |
| Groups IV & II | Average dissimilarity = 91.53 | |||||
| Group IV | Group II | |||||
| Weinmannia ovata Cav. | 0 | 2.75 | 2.83 | 1.59 | 3.09 | 3.09 |
| Guarea kunthiana A. Juss. | 2.62 | 0 | 2.76 | 2.39 | 3.01 | 6.1 |
| Aniba muca (Ruiz&Pav) Mez | 2.25 | 0 | 2.39 | 1.4 | 2.61 | 8.72 |
| Persea ferruginea Kunth | 0 | 2.2 | 2.31 | 1.22 | 2.53 | 11.24 |
| Gordonia fruticosa | 0 | 2.19 | 2.24 | 2.25 | 2.45 | 13.69 |
| Groups III & II | Average dissimilarity = 85.12 | |||||
| Group III | Group II | |||||
| Weinmannia ovata Cav. | 0 | 2.75 | 2.79 | 1.59 | 3.28 | 3.28 |
| Miconia jahnii Pittier | 0.14 | 2.61 | 2.55 | 1.9 | 2.99 | 6.27 |
| Gordonia fruticosa | 0 | 2.19 | 2.21 | 2.25 | 2.6 | 8.87 |
| Clusia alata Triana & Planch. | 1.64 | 2.07 | 2.18 | 1.31 | 2.57 | 11.43 |
| Ilex andicola Loes | 2.09 | 0 | 2.18 | 1.7 | 2.56 | 13.99 |
| Groups I & II | Average dissimilarity = 85.28 | |||||
| Group I | Group II | |||||
| Weinmannia ovata Cav. | 0 | 2.75 | 2.76 | 1.54 | 3.23 | 3.23 |
| Gordonia fruticosa (Schrad.)H.Keng | 0 | 2.19 | 2.18 | 2.15 | 2.56 | 5.79 |
| Clusia alata Triana & Planch. | 1.11 | 2.07 | 2.1 | 1.15 | 2.46 | 8.25 |
| Weinmannia elliptica Kunth | 0.06 | 2.07 | 2.01 | 1.51 | 2.36 | 10.61 |
| Miconia jahnii Pittier | 1.15 | 2.61 | 1.99 | 1.44 | 2.33 | 12.94 |
| Vectors | ||||
|---|---|---|---|---|
| NMDS1 | NMDS2 | r2 | p-Value | |
| Altitude | 0.882 | −0.469 | 0.7101 | <0.0001 |
| Temperature Season | 0.730 | −0.686 | 0.1713 | 0.0199 |
| Isothermality | 0.467 | 0.884 | 0.2462 | 0.0059 |
| Factors | ||||
| West Cordillera | −1.458 | −0.105 | ||
| East Cordillera | 0.324 | 0.023 | ||
| Physiographic province | 0.3924 | <0.0001 | ||
| Denudative | 0.5856 | 0.128 | ||
| Fluvial Erosional | −0.488 | −0.106 | ||
| Great landscape | 0.2474 | <0.0001 | ||
| Low Hills | −0.2503 | 0.093 | ||
| High Hills | 0.7127 | 0.436 | ||
| High Mountain | 0.2963 | −1.111 | ||
| Low Mountain | −0.0453 | −0.131 | ||
| Dissected Mountain | −1.458 | −0.105 | ||
| Landscape | 0.7057 | <0.0001 | ||
| Andesitic collade | 1.142 | 0.151 | ||
| Intrusive | −0.045 | −0.131 | ||
| Metamorphic | 0.016 | −0.096 | ||
| Andesitic Tuffs | −1.458 | −0.105 | ||
| Whitish Tuffs | −0.250 | 0.093 | ||
| Litology | 0.6351 | <0.0001 |
| MODEL | Estimate | Std. Error | t Value | Pr(>|t|) |
|---|---|---|---|---|
| Intercept | 0.0085217 | 0.00089 | 9.479 | <0.001 *** |
| PC 1 | −0.0012392 | 0.00028 | −4.294 | <0.001 *** |
| PC 2 | 0.0027677 | 0.00089 | 3.095 | <0.01 * |
| Parameters | G1 (Santiago-Punzara-Madrigal) | G2 (Cajanuma) | G3 (Cofradía) | G4 (Achiral) |
|---|---|---|---|---|
| Shannon Diversity Index (/400 m2) | 28.9 ± 8.6a | 30 ± 4.51b | 21.1 ± 4.3a | 22.8 ± 2.3a |
| Species Tree Richness/400 m | 2.6 ± 0.4a | 2.58 ± 0.22a | 2.44 ± 0.34a | 2.88 ± 0.12b |
| Trees/400 m (>10 cm DAP) | 86.1 ± 28.9a | 127.3 ± 40.9b | 112 ± 15.3a | 96.7 ± 17.9a |
| Basal Area (>10 cm DAP) | 2.7 ± 0.9a | 3.6 ± 1.2a | 4.9 ± 1.8b | 3.7 ± 0.6a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cabrera, O.; Benítez, Á.; Cumbicus, N.; Naranjo, C.; Ramón, P.; Tinitana, F.; Escudero, A. Geomorphology and Altitude Effects on the Diversity and Structure of the Vanishing Montane Forest of Southern Ecuador. Diversity 2019, 11, 32. https://doi.org/10.3390/d11030032
Cabrera O, Benítez Á, Cumbicus N, Naranjo C, Ramón P, Tinitana F, Escudero A. Geomorphology and Altitude Effects on the Diversity and Structure of the Vanishing Montane Forest of Southern Ecuador. Diversity. 2019; 11(3):32. https://doi.org/10.3390/d11030032
Chicago/Turabian StyleCabrera, Omar, Ángel Benítez, Nixon Cumbicus, Carlos Naranjo, Pablo Ramón, Fani Tinitana, and Adrián Escudero. 2019. "Geomorphology and Altitude Effects on the Diversity and Structure of the Vanishing Montane Forest of Southern Ecuador" Diversity 11, no. 3: 32. https://doi.org/10.3390/d11030032
APA StyleCabrera, O., Benítez, Á., Cumbicus, N., Naranjo, C., Ramón, P., Tinitana, F., & Escudero, A. (2019). Geomorphology and Altitude Effects on the Diversity and Structure of the Vanishing Montane Forest of Southern Ecuador. Diversity, 11(3), 32. https://doi.org/10.3390/d11030032