Prokaryotic Dynamics in the Meromictic Coastal Lake Faro (Sicily, Italy)


 , ,
, ,  ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
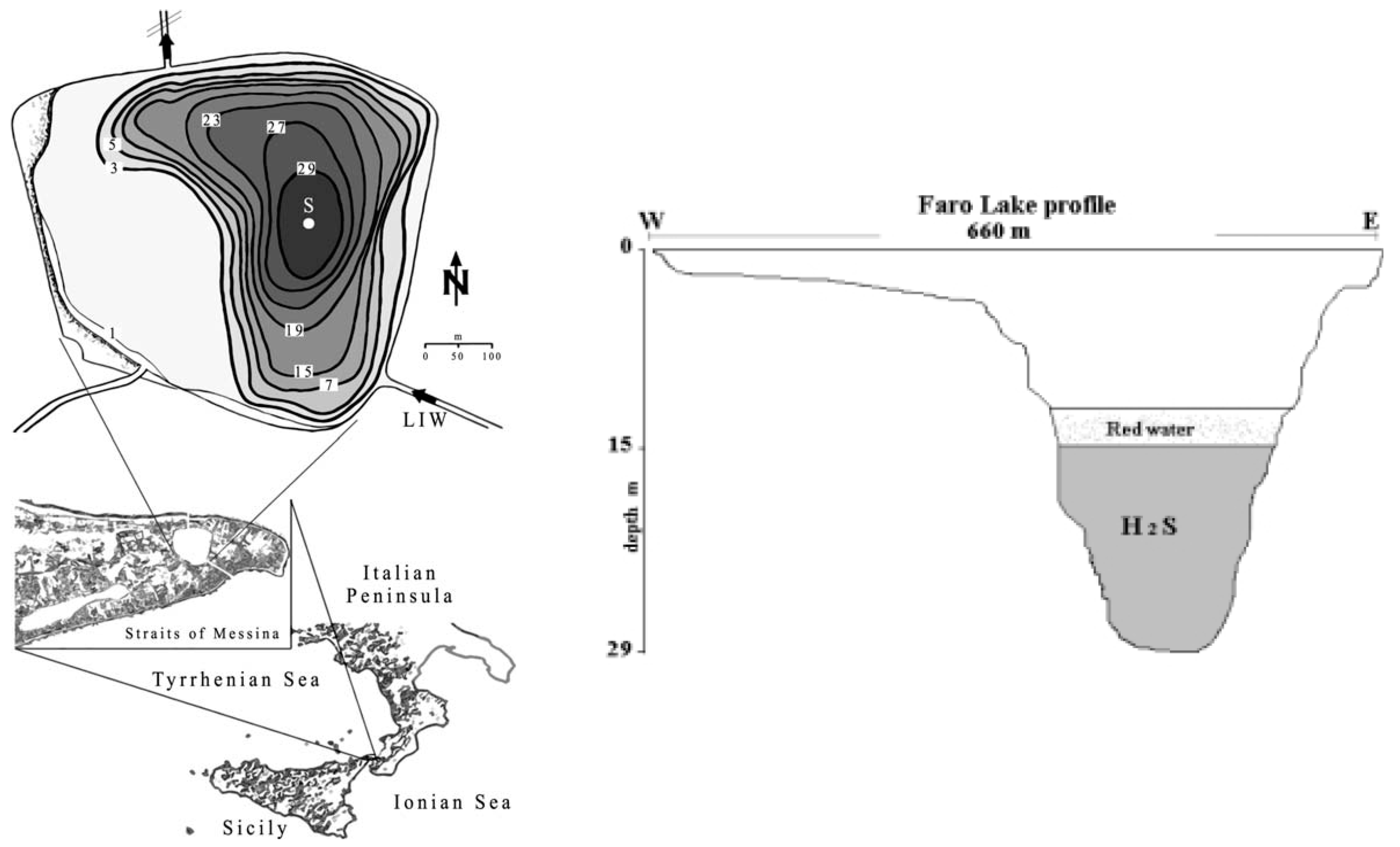
2.1. Study Area
2.2. Water Sample Collection
2.2.1. Nutrient Analyses
2.2.2. Ammonia, Nitrites and Nitrates
2.2.3. Phosphates
2.2.4. TC/TOC/TN Analyses
2.3. Microbial Abundances and Distribution
2.3.1. Total Counts
2.3.2. Automated Ribosomal Intergenic Spacer Analysis (ARISA)
2.3.3. Catalyzed Reporter Deposition–Fluorescence In Situ Hybridization (CARD-FISH)
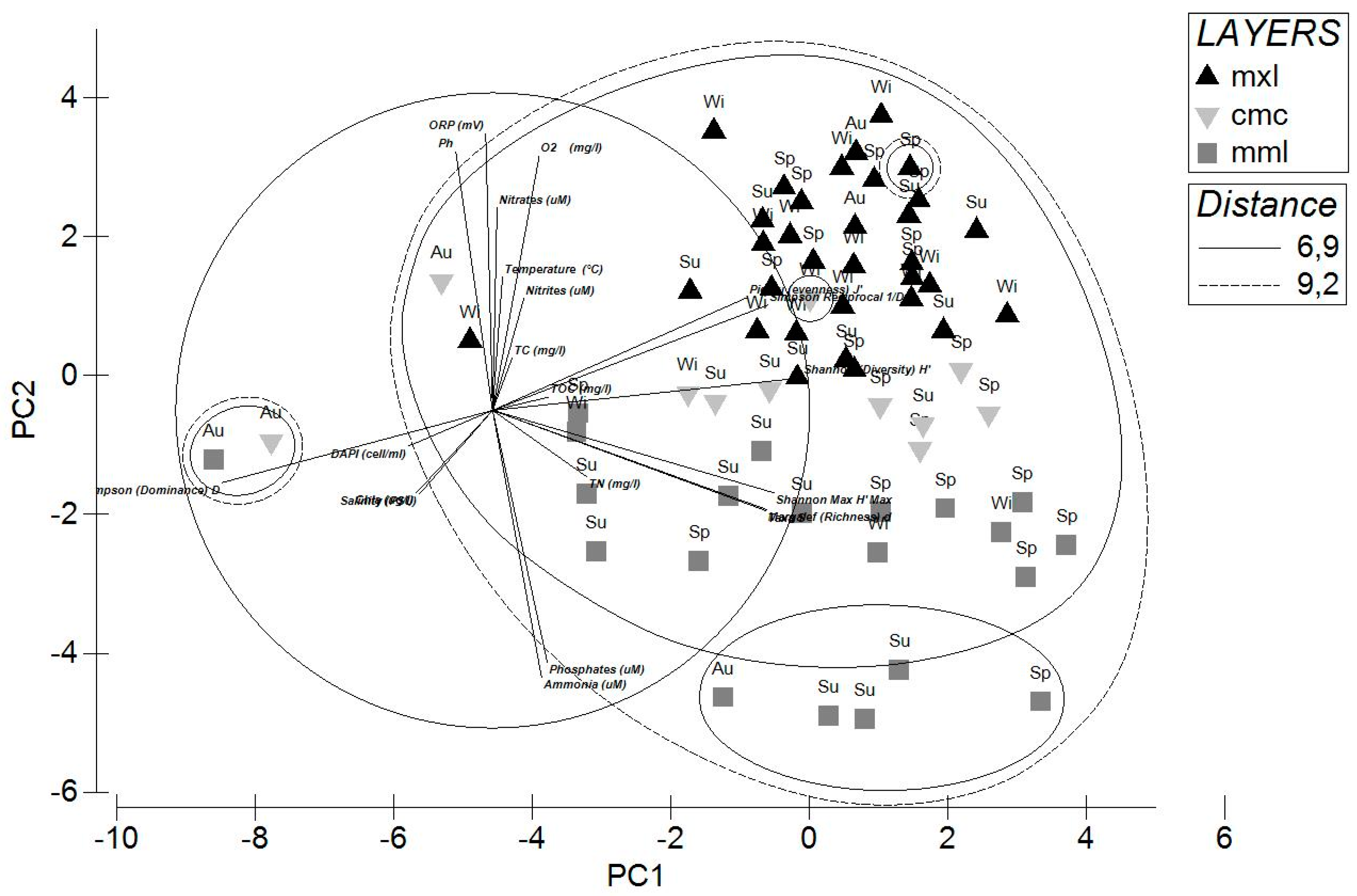
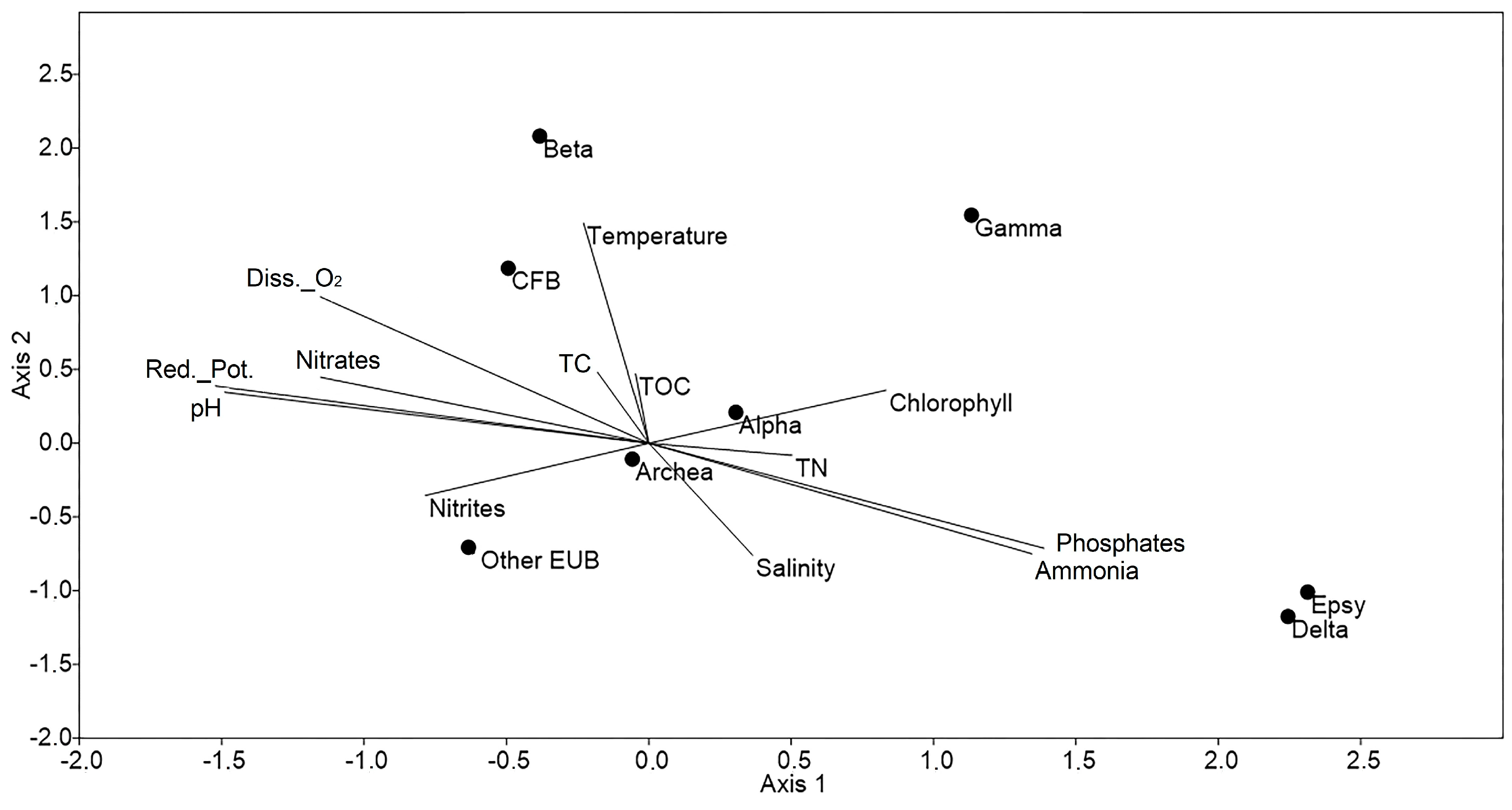
2.4. Statistical Analyses
3. Results
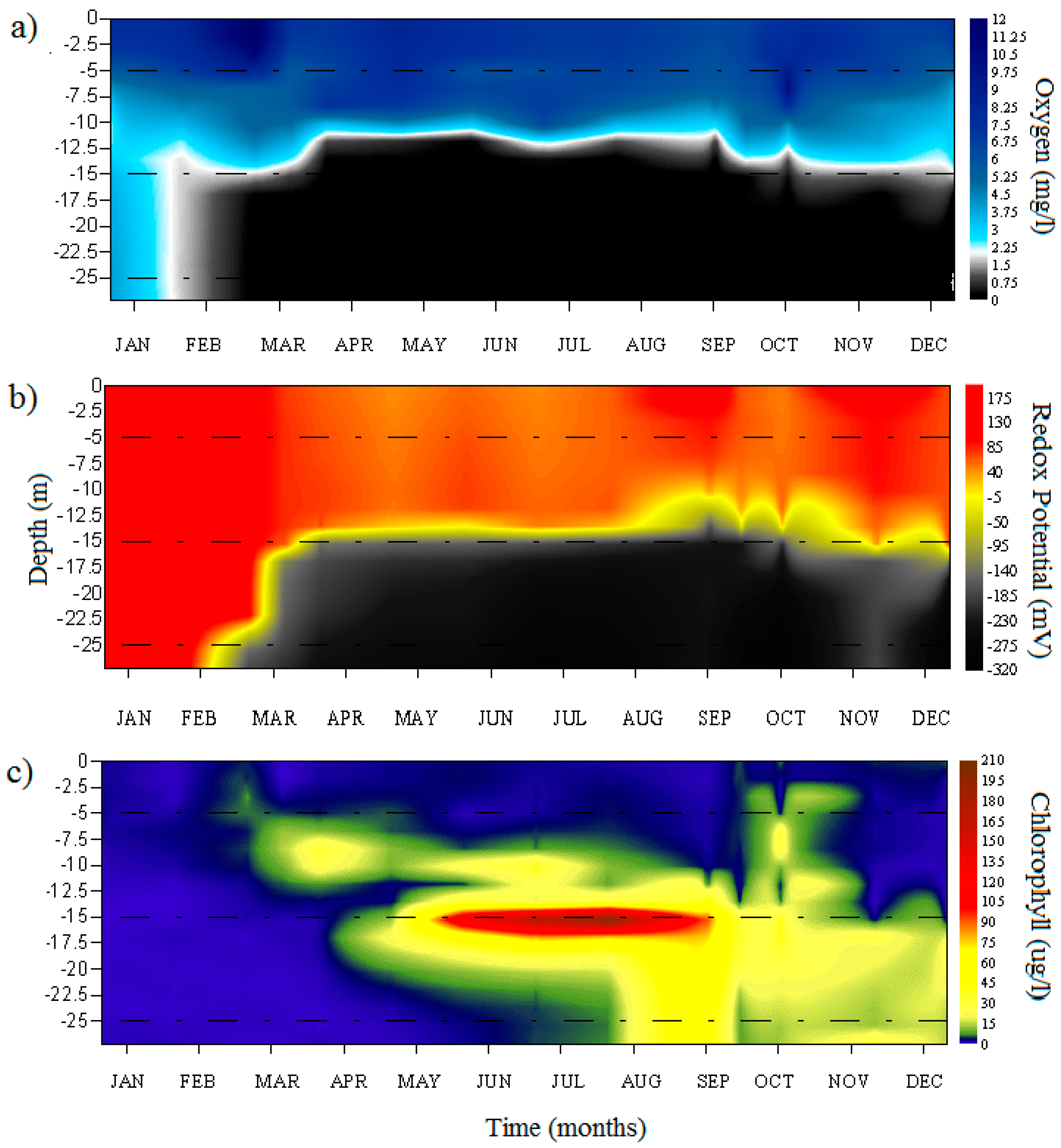
3.1. Environmental Parameters
3.2. Nutrient Concentration
3.2.1. Ammonia, nitrites and nitrates
3.2.2. Phosphates
3.2.3. TC/TOC/TN Analyses
3.3. Microbial Abundances and Distribution
3.3.1. Total Counts
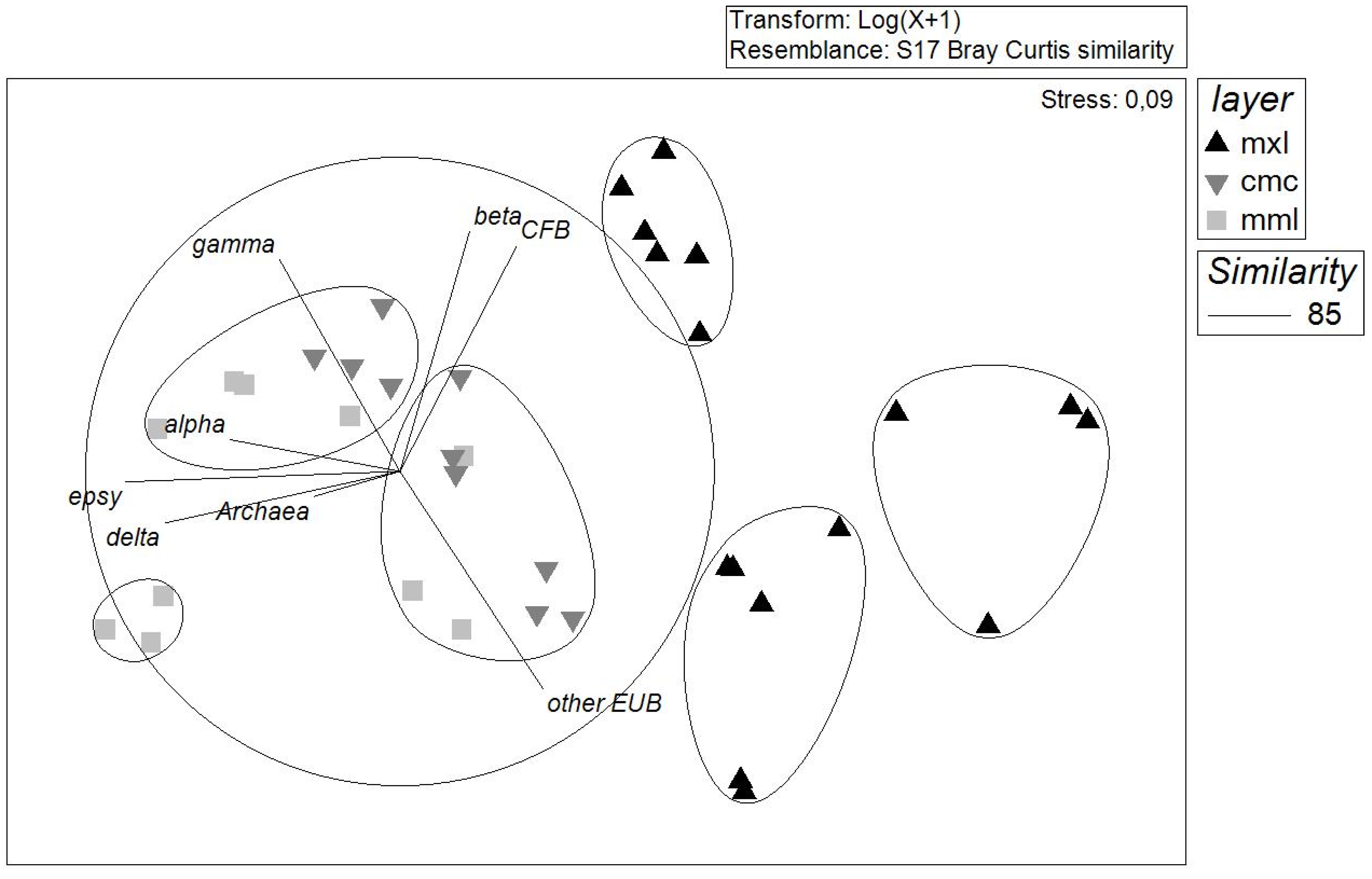
3.3.2. Microbial Community Structure Determined with ARISA
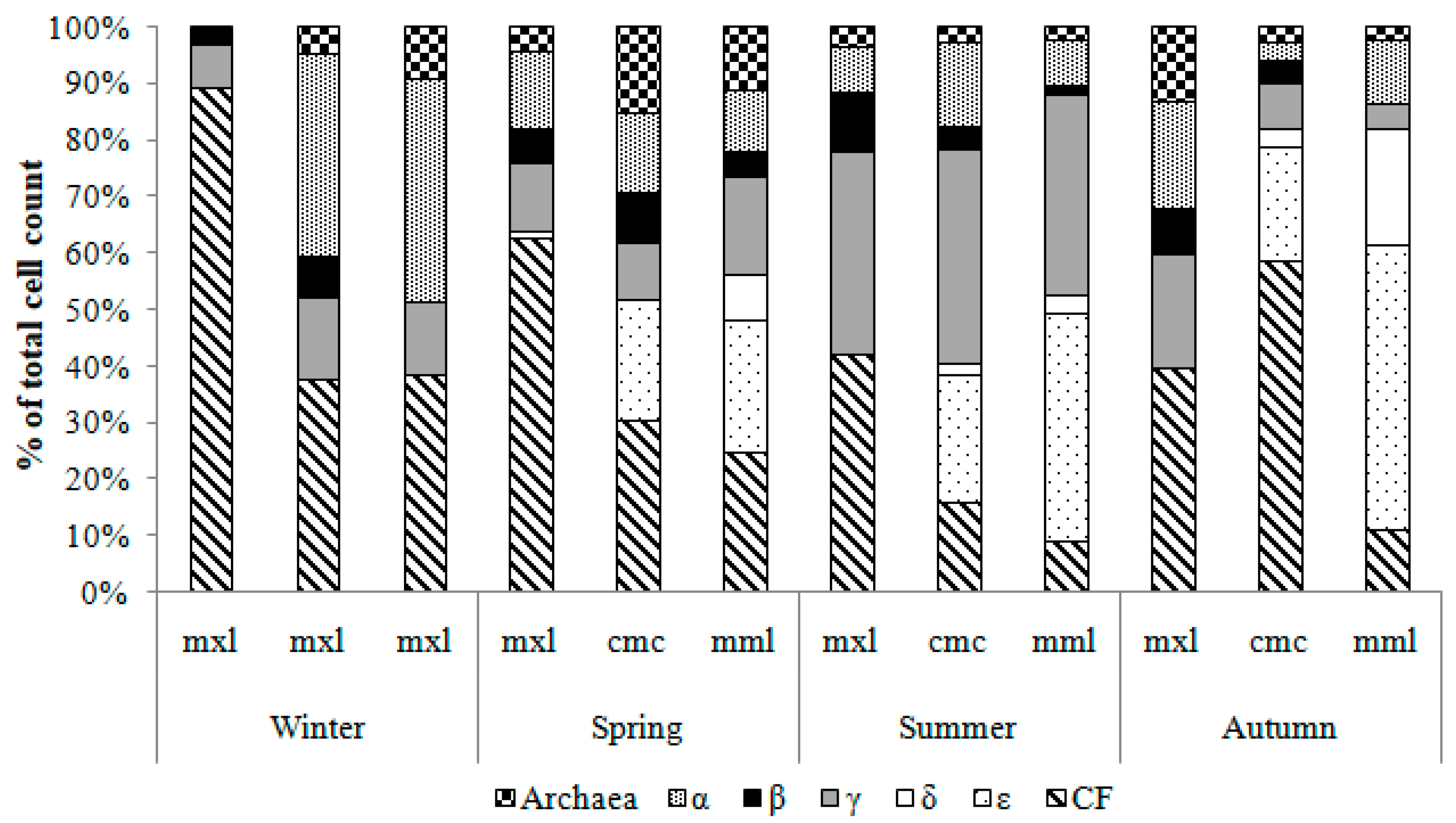
3.3.3. Abundance of the Major Phylogenetic Groups by CARD-FISH Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Andrei, A.Ş.; Robeson, M.S.; Baricz, A.; Coman, C.; Muntean, V.; Ionescu, A.; Etiope, G.; Alexe, M.; Sicora, C.I.; Podar, M.; et al. Contrasting taxonomic stratification of microbial communities in two hypersaline meromictic lakes. ISME J. 2015, 9, 2642–2656. [Google Scholar] [CrossRef] [PubMed]
- Baatar, B.; Chiang, P-W.; Rogozin, D.Y.; Wu, Y.T.; Tseng, C-H.; Yang, C-Y.; Chiu, H-H.; Oyuntsetseg, B.; Degermendzhy, A.G.; Tang, S-L. Bacterial communities of three saline meromicticlakes in central Asia. PLoS ONE 2016, 11, e0150847. [Google Scholar] [CrossRef] [PubMed]
- Gugliandolo, C.; Lentini, V.; Maugeri, T. Distribution and diversity of bacteria in a saline meromictic lake as determined by PCR-DGGE of 16S rRNAgene fragments. Curr. Microbiol. 2010, 63, 3367–3373. [Google Scholar]
- Lauro, F.M.; DeMaere, M.Z.; Yau, S.; Brown, M.V.; Ng, C.; Wilkins, D.; Raftery, M.J.; Gibson, J.A.; Andrews-Pfannkoch, C.; Lewis, M.; et al. An integrative study of a meromictic lake ecosystem in Antarctica. ISME J. 2010, 5, 879–895. [Google Scholar] [CrossRef] [PubMed]
- Camacho, A.; Erez, J.; Chicote, A.; Florin, M.; Squires, M.M.; Lehmann, C.; Bachofen, R. Microbial microstratification, inorganic carbon photoassimilation and dark carbon fixation at the chemocline of the meromictic Lake Cadagno (Switzerland) and its relevance to the food web. Aquat. Sci. 2001, 63, 91–106. [Google Scholar] [CrossRef]
- Lehours, A.C.; Bardot, C.; Thenot, A.; Debroas, D.; Fonty, G. Anaerobic microbial communities in Lake Pavin, a unique meromictic lake in France. Appl. Environ. Microbiol. 2005, 71, 7389–7400. [Google Scholar] [CrossRef] [PubMed]
- Fenchel, T.; King, G.H.; Blackburn, T.H. Bacterial Biogeochemistry, the Ecophysiology of Mineral Cycling; Elsevier Science, Academic Press: Cambridge, MA, USA, 1998; pp. 307–309. [Google Scholar]
- Nõges, T.; Solovjova, I. The formation and dynamics of deep bacteriochlorophyll maximum in the temperate and partly meromictic Lake Verevi. Hydrobiologia 2005, 547, 73–81. [Google Scholar] [CrossRef]
- Øvreas, L.; Forney, L.; Daae, F.L.; Torsvik, V. Distribution of bacterioplankton in meromictic Lake Saelenvannet, as determined by denaturing gradient gel electrophoresis of PCR-amplified gene fragments coding for 16S rRNA. Appl. Environ. Microbiol. 1997, 63, 3367–3373. [Google Scholar] [PubMed]
- Peura, S.; Eiler, A.; Bertilsson, S.; Nykänen, H.; Tiirola, M.; Jones, R.I. Distinct and diverse anaerobic bacterial communities in boreal lakes dominated by candidate division OD1. ISME J. 2012, 6, 1640–1652. [Google Scholar] [CrossRef] [PubMed]
- Rogozin, D.Y.; Trusova, M.Y.; Khromechek, E.B.; Degermendzhy, A.G. Microbial community of the chemocline of the meromictic Lake Shunet (Khakassia, Russia) during summer stratification. Microbiology 2010, 79, 253–261. [Google Scholar] [CrossRef]
- Comeau, A.M.; Harding, T.; Galand, P.E.; Vincent, W.F.; Lovejoy, C. Vertical distribution of microbial communities in a perennially stratified Arctic lake with saline, anoxic bottom waters. Sci. Rep. 2012, 2, 604. [Google Scholar] [CrossRef] [PubMed]
- Yau, S.; Lauro, F.M.; Williams, T.J.; DeMaere, M.Z.; Brown, M.V.; Rich, J.; Gibson, J.A.; Cavicchioli, R. Metagenomic insights into strategies of carbon conservation and unusual sulfur biogeochemistry in a hypersaline Antarctic lake. ISME J. 2013, 7, 1944–1961. [Google Scholar] [CrossRef] [PubMed]
- Ramsing, N.B.; Fossing, H.; Ferdelmann, T.G.; Andersen, F.; Thamdrup, B. Distribution of bacterial populations in a stratified fjord (Mariager Fjord, Denmark) quantified by in situ hybridization and related to chemical gradients in the water column. Appl. Environ. Microbiol. 1996, 62, 1391–1404. [Google Scholar] [PubMed]
- Teske, A.; Wawer, C.; Muyzer, G.; Ramsing, N.B. Distribution of sulfate-reducing bacteria in a stratified fjord (Mariager Fjord, Denmark) as evaluated by most-probable-number counts and denaturing gradient gel electrophoresis of PCR-amplified ribosomal DNA fragments. Appl. Environ. Microbiol. 1996, 62, 1405–1415. [Google Scholar] [PubMed]
- Tuomi, P.; Torsvik, T.; Heldal, M.; Bratbak, G. Bacterial population dynamics in a meromictic lake. Appl. Environ. Microbiol. 1997, 63, 2181–2188. [Google Scholar] [PubMed]
- Bosshard, P.P.; Santini, Y.; Grüter, D.; Stettler, R.; Bachofen, R. Bacterial diversity and community composition in the chemocline of the meromictic alpine Lake Cadagno as revealed by 16S rDNA analysis. FEMS Microbiol. Ecol. 2000, 31, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Humayoun, S.B.; Bano, N.; Hollibaugh, J.T. Depth distribution of microbial diversity in Mono Lake, a meromictic soda lake in California. Appl. Environ. Microbiol. 2003, 69, 1030–1042. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, Y.; Kojima, H.; Oguri, K.; Kitazato, Y.; Fukui, M. Vertical and temporal shifts in microbial communities in the water column and sediment of saline meromictic Lake Kaiike (Japan), as determined by a 16S rDNA-based analysis, and related to physicochemical gradients. Environ. Microbiol. 2004, 6, 622–637. [Google Scholar] [CrossRef] [PubMed]
- Lehours, A.C.; Evans, P.; Bardot, C.; Joblin, K.; Gérard, F. Phylogenetic diversity of archaea and bacteria in the anoxic zone of a meromictic lake (Lake Pavin, France). Appl. Environ. Microbiol. 2007, 6, 2016–2019. [Google Scholar] [CrossRef] [PubMed]
- Kondo, R.; Nakagawa, A.; Mochizuki, L.; Osawa, K.; Fujioka, Y.; Butani, J. Dominant bacterioplankton populations in the meromictic Lake Suigetsu as determined by denaturing gradient gel electrophoresis of 16S rRNA gene fragments. Limnology 2009, 10, 63–69. [Google Scholar] [CrossRef]
- Saccà, A.; Borrego, C.M.; Renda, R.; Triadò-Margarit, X.; Bruni, V.; Guglielmo, L. Predation impact of ciliated and flagellated protozoa during a summer bloom of brown sulfur bacteria in a meromictic coastal lake. FEMS Microbiol. Ecol. 2009, 70, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Saccà, A.; Guglielmo, L.; Bruni, V. Vertical and temporal microbial community patterns in a meromictic coastal lake influenced by the Straits of Messina upwelling system. Hydrobiologia 2008, 600, 89–104. [Google Scholar] [CrossRef]
- La Cono, V.; La Spada, G.; Arcadi, E.; Placenti, F.; Smedile, F.; Ruggeri, G.; Michaud, L.; Raffa, C.; De Domenico, E.; Sprovieri, M.; et al. Partaking of Archaea to biogeochemical cycling in oxygen-deficient zones of meromictic saline Lake Faro (Messina, Italy). Environ. Microbiol. 2012, 15, 1717–1733. [Google Scholar] [CrossRef] [PubMed]
- Giuffrè, G.; Pezzani, R. Annual nano-microphytoplankton succession in the meromictic brackish Lake Faro (Messina - Sicily) in relation to physical-chemical conditions. Nat. Sicil. 2005, 29, 153–168. [Google Scholar]
- Aminot, A.; Chaussepied, M. Manuel del Analyses Chimiques en Milieu Marin; Centre National pour l’Exploration des Oceans: Brest, France, 1983; pp. 107–118. [Google Scholar]
- Bendschneider, K.; Robinson, R.J. A new spectrophotometric determination of nitrite in sea water. J. Mar. Res. 1952, 11, 87–96. [Google Scholar]
- Wood, E.D.; Armstrong, A.J.; Richards, F.A. Determination of nitrate in sea water by cadmium-copper reduction to nitrite. J. Mar. Biol. Ass. U.K. 1967, 47, 23–31. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. An. Chem. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Porter, K.J.; Feig, Y.S. The use of DAPI for identifying and counting aquatic microflora. Limnol. Oceanogr. 1980, 25, 943–948. [Google Scholar] [CrossRef]
- Stepanauskas, R.; Moran, M.A.; Bergamaschi, B.A.; Hollibaugh, J.T. Covariance of bacterioplankton composition and environmental variables in a temperate delta system. Aquat. Microb. Ecol. 2003, 31, 85–98. [Google Scholar] [CrossRef]
- Luna, G.M.; Dell’Anno, A.; Danovaro, R. DNA extraction procedure: a critical issue for bacterial diversity assessment in marine sediments. Environ. Microbiol. 2006, 8, 308–320. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, M.; Brusetti, L.; Quatrini, P.; Borin, S.; Puglia, A.M.; Rizzi, A.; Zanardini, E.; Sorlini, C.; Corselli, C.; Daffonchio, D. Comparison of different primer sets for use in automated ribosomal intergenic spacer analysis of complex bacterial communities. Appl. Environ. Microbiol. 2004, 70, 6147–6156. [Google Scholar] [CrossRef] [PubMed]
- Ramette, A. Quantitative community fingerprinting methods for estimating the abundance of operational taxonomic units in natural microbial communities. Appl. Environ. Microbiol. 2009, 75, 2495–2505. [Google Scholar] [CrossRef] [PubMed]
- Porporato, E.M.; Lo Giudice, A.; Michaud, L.; De Domenico, E.; Spanò, N. Diversity and antibacterial activity of the bacterial communities associated with two Mediterranean sea pens, Pennatula phosphorea and Pteroeides spinosum (Anthozoa: Octocorallia). Microb. Ecol. 2013, 66, 701–714. [Google Scholar] [CrossRef] [PubMed]
- Dunbar, J.; Ticknor, L.O.; Kuske, C.R. Phylogenetic specificity and reproducibility and new method for analysis of terminal restriction fragment profiles of 16S rRNA genes from bacterial communities. Appl. Environ. Microbiol. 2001, 67, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Amann, R.I.; Binder, B.J.; Olson, R.J.; Chisholm, S.W.; Devereux, R.; Stahl, D.A. Combination of 16S rRNA- targeted oligonucleotide probes with flow cytometry for analyzing mixed microbial populations. Appl. Environ. Microbiol. 1990, 56, 1919–1925. [Google Scholar] [PubMed]
- Daims, H.; Bruhl, A.; Amann, R.; Schleifer, K.H.; Wagner, M. The domain-specific probe EUB338 is insufficient for the detection of all Bacteria: Development and evaluation of a more comprehensive probe set. System. Appl. Microbiol. 1999, 22, 434–444. [Google Scholar] [CrossRef]
- Stahl, D.A.; Amann, R. Development and application of nucleic acid probes. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: Hoboken, NJ, USA, 1991; pp. 205–248. [Google Scholar]
- Neef, A.; Amann, R.; Schlesner, H.; Schleifer, K.H. Monitoring a widespread bacterial group: in situ detection of planctomycetes with 16S rRNA-targeted probes. Microbiology 1998, 144, 3257–3266. [Google Scholar] [CrossRef] [PubMed]
- Manz, W.; Amann, R.; Ludwig, W.; Wagner, M.; Schleifer, K.H. Phylogenetic oligodeoxynucleotide probes for the major subclasses of proteobacteria: problems and solutions. Syst. Appl. Microbiol. 1992, 15, 593–600. [Google Scholar] [CrossRef]
- Loy, A.; Lehner, A.; Lee, N.; Adamczyk, J.; Meier, H.; Ernst, J.; Schleifer, K.H.; Wagner, M. Oligonucleotide microarray for 16S rRNA gene based detection of all recognized lineages of sulfate reducing prokaryotes in the environment. Appl. Environ. Microbiol. 2002, 68, 5064–5081. [Google Scholar] [CrossRef] [PubMed]
- Loy, A.; Maixner, F.; Wagner, M.; Horn, M. probeBase—An online resource for rRNA-targeted oligonucleotide probes: New features. Nucleic Acids Res. 2007, 35, D800–D804. [Google Scholar] [CrossRef] [PubMed]
- Manz, W.; Amann, R.; Ludwig, W.; Vancanneyt, M.; Schleifer, K.H. Application of a suite of 16S rRNA specific oligonucleotide probes designed to investigate bacteria of the phylum Cytophaga-Flavobacter-Bacteroides in the natural environment. Microbiology 1996, 142, 1097–1106. [Google Scholar] [CrossRef] [PubMed]
- Wallner, G.; Amann, R.; Beisker, W. Optimizing fluorescent in situ hybridization of suspended cells with rRNA-targeted oligonucleotide probes for the flow cytometric identification of microorganisms. Cytometry 1993, 14, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Pernthaler, A.; Amann, R. Simultaneous fluorescence in situ hybridization of mRNA and rRNA in environmental bacteria. Appl. Environ. Microbiol. 2004, 70, 5426–5433. [Google Scholar] [CrossRef] [PubMed]
- Danovaro, R.; Luna, G.M.; Dell’Anno, A.; Pietrangeli, B. Comparison of two fingerprinting techniques, terminal restriction fragment length polymorphism and automated ribosomal intergenic spacer analysis, for determination of bacterial diversity in aquatic environments. Appl. Environ. Microbiol. 2006, 72, 5982–5989. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics software for education and data analysis. Palaeontol. Electronica 2001, 4, 1–9. [Google Scholar]
- Rodrigo, M.A.; Miracle, M.R.; Vicente, E. The meromictic Lake La Cruz (Central Spain). Patterns of stratification. Aquat. Sci. 2001, 63, 406–416. [Google Scholar] [CrossRef]
- Crisafi, P. Nuove osservazioni sulla distribuzione dell’idrogeno solforato nel lago di Faro (Messina). Atti Soc. Pelorit. Sci. Fis. Mat. Nat. 1956, 4, 309–314. [Google Scholar]
- Sorokin, Y.I.; Donato, N. On the carbon and sulphur metabolism in the meromictic Lake Faro (Sicily). Hydrobiologia 1975, 47, 241–252. [Google Scholar] [CrossRef]
- Genovese, S. Sul fenomeno dell’ ”acqua rossa” riscontrato nello stagno salmastro di Faro. Atti Soc. Pelorit. Sci. Fis. Mat. Nat. 1961, 7, 269–271. [Google Scholar]
- Trüper, H.G.; Genovese, S. Characterization of photosynthetic sulfur bacteria causing red water in Lake Faro (Messina, Sicily). Limnol. Oceanog. 1968, 13, 225–232. [Google Scholar] [CrossRef]
- Bruni, V.; Grillo, O.C. Ulteriori indagini sulla presenza dell’acqua rossa nel lago di Faro. Atti Soc. Pelorit. Sci. Fis. Mat. Nat. 1981, 24, 291–299. [Google Scholar]
- Acosta Pomar, L.; Bruni, V.; Decembrini, F.; Giuffrè, G.; Maugeri, T.L. Distribution and activity of picophytoplankton in a brackish environment. Progr. Oceanogr. 1988, 21, 129–138. [Google Scholar] [CrossRef]
- Grote, J.; Jost, G.; Labrenz, M. Epsilonproteobacteria represent the major portion of chemoautotrophic bacteria in sulfidic waters of pelagic redoxclines of the Baltic and Black Seas. Appl. Environ. Microbiol. 2008, 74, 7546–7551. [Google Scholar] [CrossRef] [PubMed]
- Dimitriu, P.A.; Pinkart, H.C.; Peyton, B.M.; Mormile, M.R. Spatial and temporal patterns in the microbial diversity of a meromictic soda lake in Washington State. Appl. Environ. Microbiol. 2008, 74, 4877–4888. [Google Scholar] [CrossRef] [PubMed]
- Klepac-Ceraj, V.; Hayes, C.A.; Gilhooly, W.P.; Lyons, T.W.; Kolter, R.; Pearson, A. Microbial diversity under extreme euxinia: Mahoney Lake, Canada. Geobiology 2012, 10, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Bowman, J.P.; McCammon, S.A.; Rea, S.M.; McMeekin, T.A. The microbial composition of three limnologically disparate hypersaline Antarctic lakes. FEMS Microbiol. Lett. 2000, 183, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Casamayor, E.O.; Triadó-Margarit, X.; Castañeda, C. Microbial biodiversity in saline shallow lakes of the Monegros Desert, Spain. FEMS Microbiol. Ecol. 2013, 85, 503–518. [Google Scholar] [CrossRef] [PubMed]
- Tiodjio, R.E.; Sakatoku, A.; Nakamura, A.; Tanaka, D.; Fantong, W.Y.; Tchakam, K.B. Bacterial and archaeal communities in Lake Nyos (Cameroon, Central Africa). Sci. Rep. 2014, 4, 6151. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Wakeham, S.G.; Putnam, I.F.; Astor, Y.M.; Scranton, M.I.; Chistoserdov, A.Y.; Taylor, G.T. Comparison of vertical distributions of prokaryotic assemblages in the anoxic Cariaco Basin and Black Sea by use of fluorescence in situ hybridization. Appl. Environ. Microbiol. 2006, 4, 2679–2690. [Google Scholar] [CrossRef] [PubMed]
- Grote, J.; Labrenz, M.; Pfeiffer, B; Jost, G.; Jürgens, K. Quantitative distributions of Epsilonproteobacteria and a Sulfurimonas subgroup in pelagic redoxclines of the Central Baltic Sea. Appl. Environ. Microbiol. 2007, 73, 7155–7161. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.J.; Craig Cary, S. Abundance of reverse tricarboxylic acid cycle genes in free-living microorganisms at deep-sea hydrothermal vents. Appl. Environ. Microbiol. 2004, 70, 6282–6289. [Google Scholar] [CrossRef] [PubMed]
- Takai, K.; Campbell, B.J.; Cary, S.C.; Suzuki, M.; Oida, H.; Nunoura, T.; Hirayama, H.; Nakagawa, S.; Suzuki, Y.; Inagaki, F.; et al. Enzymatic and genetic characterization of carbon and energy metabolisms by deep-sea hydrothermal chemolithoautotrophic isolates of Epsilonproteobacteria. Appl. Environ. Microbiol. 2005, 71, 7310–7320. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Kato, C.; Horikoshi, K. Bacterial diversity in deep-sea sediments from different depths. Biodiv. Conserv. 1999, 8, 659–677. [Google Scholar] [CrossRef]
- Yanagibayashi, M.; Nogi, Y.; Li, L.; Kato, C. Changes in the microbial community in Japan Trench sediment from a depth of 6292 m during cultivation without decompression. FEMS Microbiol. Lett. 1999, 170, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Rath, J.; Wu, K.Y.; Herndl, G.J.; DeLong, E.F. High phylogenetic diversity in a marine-snow-associated bacterial assemblage. Aquat. Microb. Ecol. 1998, 14, 261–269. [Google Scholar] [CrossRef]
- Campbell, B.J.; Stein, J.L.; Craig Cary, S. Evidence of chemolithoautotrophy in the bacterial community associated with Alvinella pompejana, a hydrothermal vent polychaete. Appl. Environ. Microbiol. 2003, 69, 5070–5078. [Google Scholar] [CrossRef] [PubMed]
- Vandamme, P.; De Ley, J. Proposal for a new family, Campylobacteraceae. Int. J. Syst. Bacteriol. 1992, 41, 451–455. [Google Scholar] [CrossRef]
- Noguerola, I.; Picazo, A.; Llirós, M.; Camacho, A.; Borrego, C.M. Diversity of freshwater Epsilonproteobacteria and dark inorganic carbon fixation in the sulphidic redoxcline of a meromictic karstic lake. FEMS Microbiol. Ecol. 2015, 91, 7. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.J.; Engel, A.S.; Porter, M.L. The versatile Epsilonproteobacteria: key players in sulphidic habitats. Nat. Rev. Microbiol. 2006, 4, 458–468. [Google Scholar] [CrossRef] [PubMed]
- Barberán, A.; Casamayor, E.O. Euxinic freshwater hypolimnia promote bacterial endemicity in continental areas. Microb. Ecol. 2010, 61, 465–472. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raffa, C.; Rizzo, C.; Strous, M.; Domenico, E.D.; Sanfilippo, M.; Michaud, L.; Giudice, A.L. Prokaryotic Dynamics in the Meromictic Coastal Lake Faro (Sicily, Italy). Diversity 2019, 11, 37. https://doi.org/10.3390/d11030037
Raffa C, Rizzo C, Strous M, Domenico ED, Sanfilippo M, Michaud L, Giudice AL. Prokaryotic Dynamics in the Meromictic Coastal Lake Faro (Sicily, Italy). Diversity. 2019; 11(3):37. https://doi.org/10.3390/d11030037
Chicago/Turabian StyleRaffa, Carmela, Carmen Rizzo, Marc Strous, Emilio De Domenico, Marilena Sanfilippo, Luigi Michaud, and Angelina Lo Giudice. 2019. "Prokaryotic Dynamics in the Meromictic Coastal Lake Faro (Sicily, Italy)" Diversity 11, no. 3: 37. https://doi.org/10.3390/d11030037
APA StyleRaffa, C., Rizzo, C., Strous, M., Domenico, E. D., Sanfilippo, M., Michaud, L., & Giudice, A. L. (2019). Prokaryotic Dynamics in the Meromictic Coastal Lake Faro (Sicily, Italy). Diversity, 11(3), 37. https://doi.org/10.3390/d11030037