A Comprehensive Study of Orchid Seed Production Relative to Pollination Traits, Plant Density and Climate in an Urban Reserve in Western Australia


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Study Species and Survey Methods
2.3. Analysis of Data
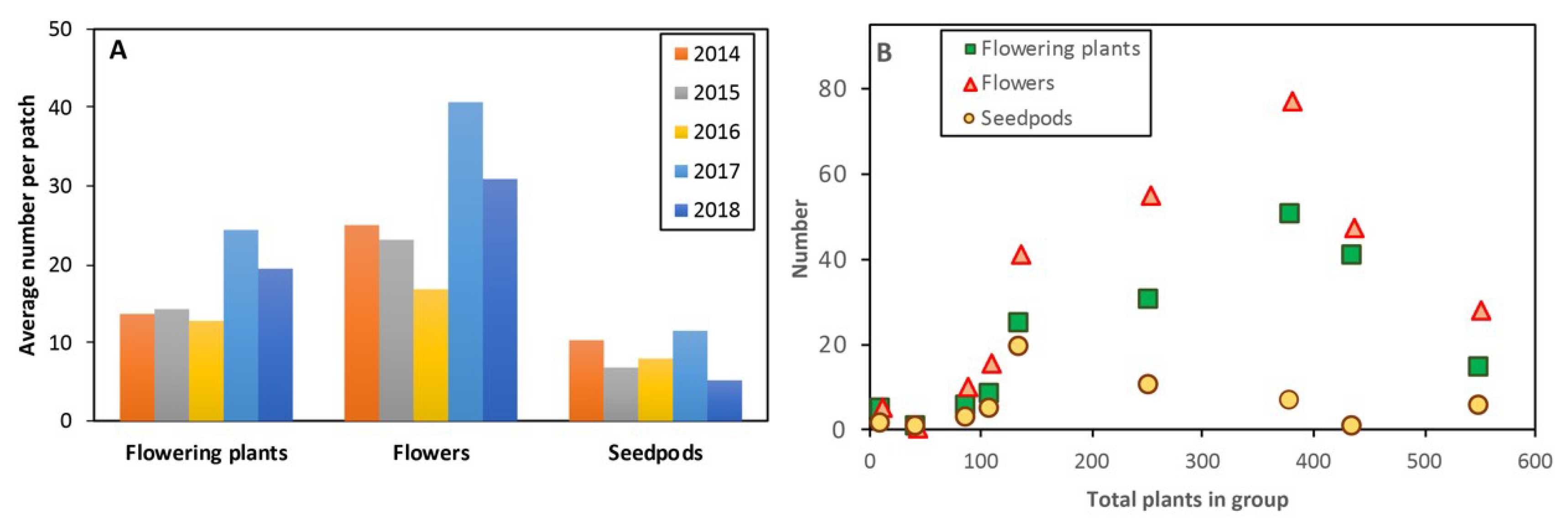
3. Results
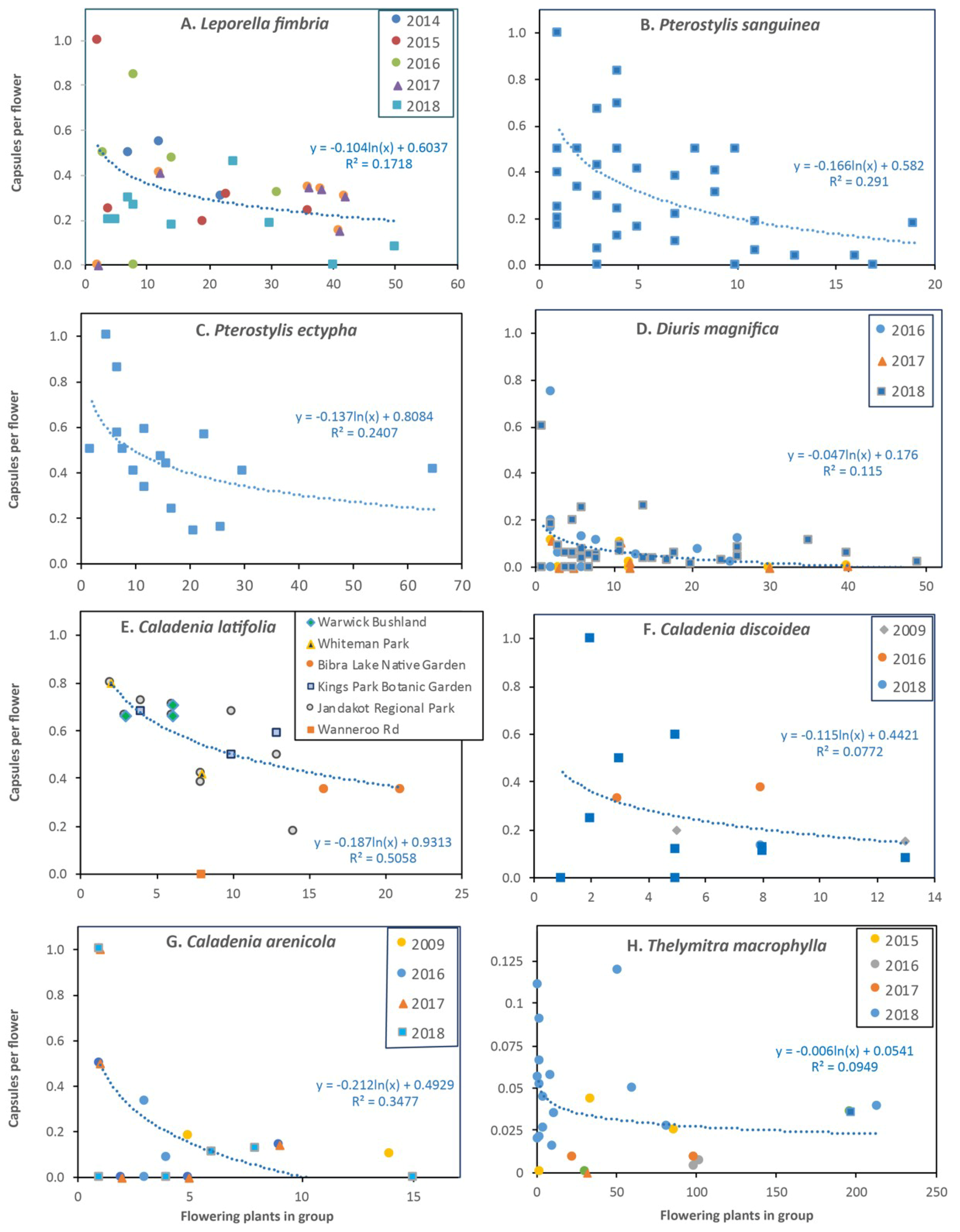
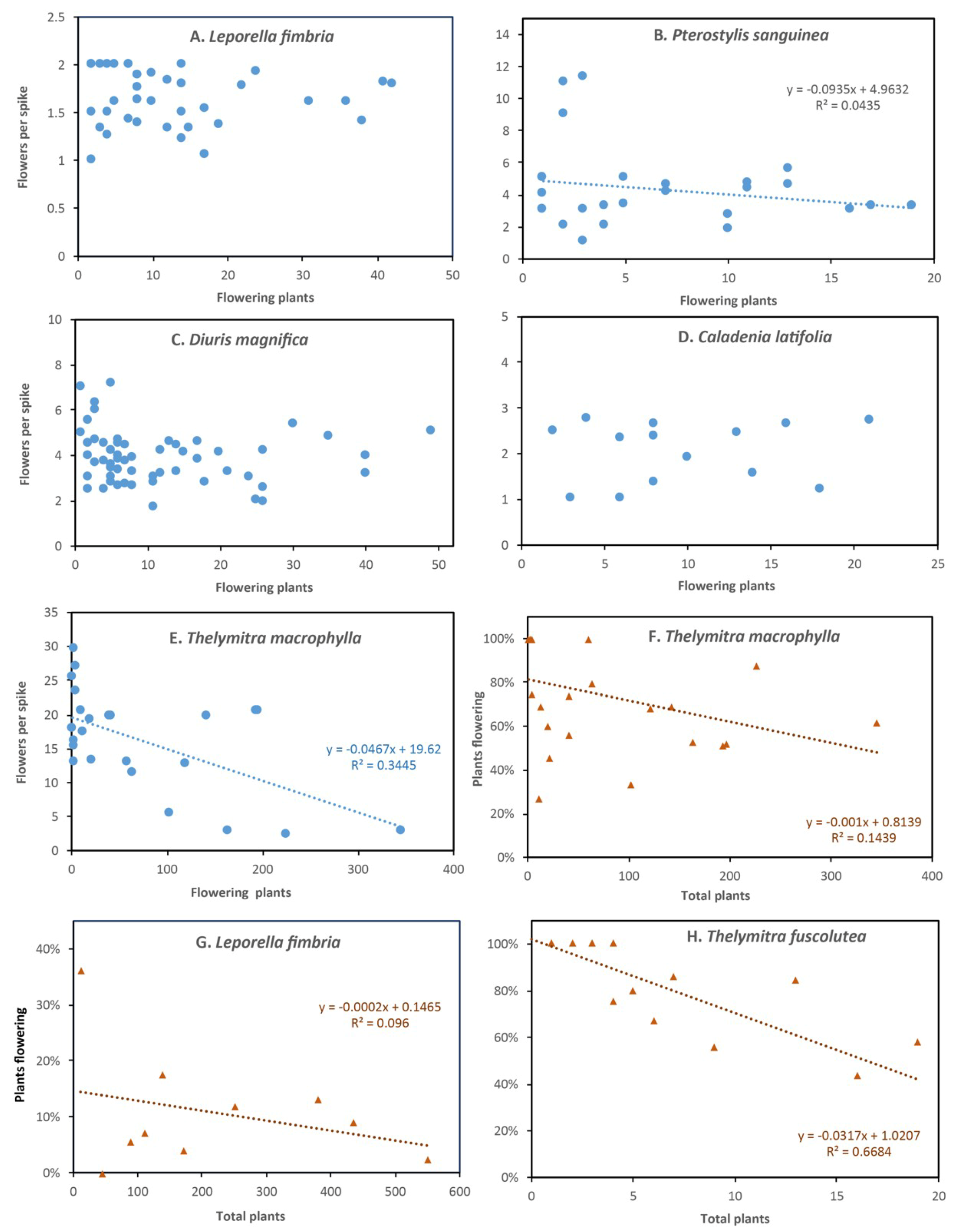
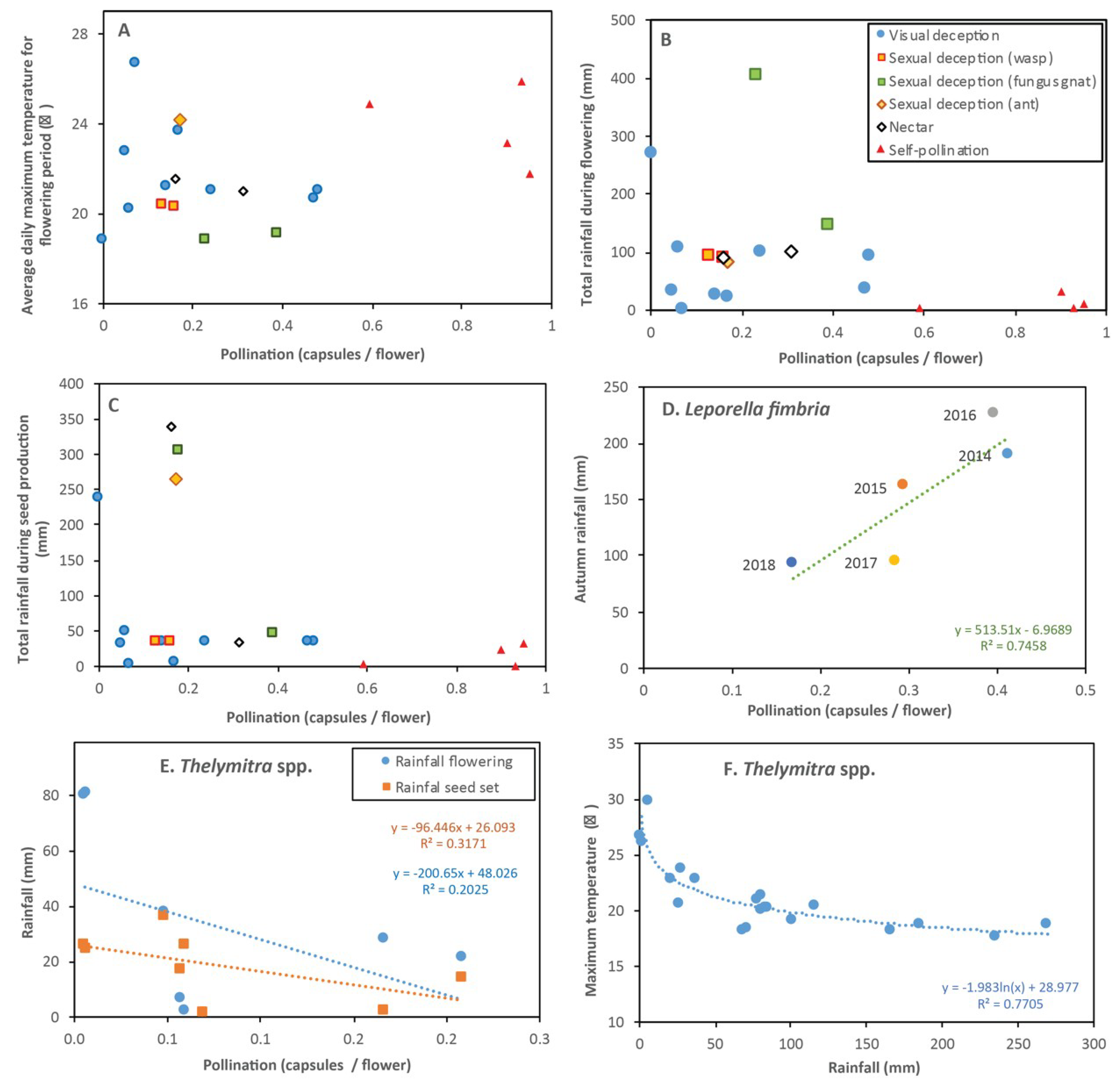
3.1. Leporella fimbriata (Hare Orchid)
3.2. Eriochilus dilatatus (Bunny Orchid)
3.3. Pterostylis sanguinea (Banded Greenhood Orchid)
3.4. Pheladenia deformis (Blue Beard Orchid)
3.5. Pterostylis ectypha (Short-Sepalled Snail Orchid)
3.6. Diuris magnifica (Pansy Orchid)
3.7. Pyrorchis nigricans (Red Beak Orchid)
3.8. Elythranthera brunonis (Purple Enamel Orchid)
3.9. Caladenia flava (Cowslip Orchid)
3.10. Caladenia latifolia (Pink Fairy Orchid)
3.11. Caladenia longicauda (White Spider Orchid)
3.12. Caladenia discoidea (Dancing Bee Orchid)
3.13. Caladenia arenicola (Carousel Spider Orchid)
3.14. Thelymitra vulgaris (Common Sun Orchid)
3.15. Thelymitra macrophylla (Blue Lady Orchid)
3.16. Thelymitra graminea (Slender Blue Sun-Orchid)
3.17. Thelymitra benthamiana (Leopard Orchid)
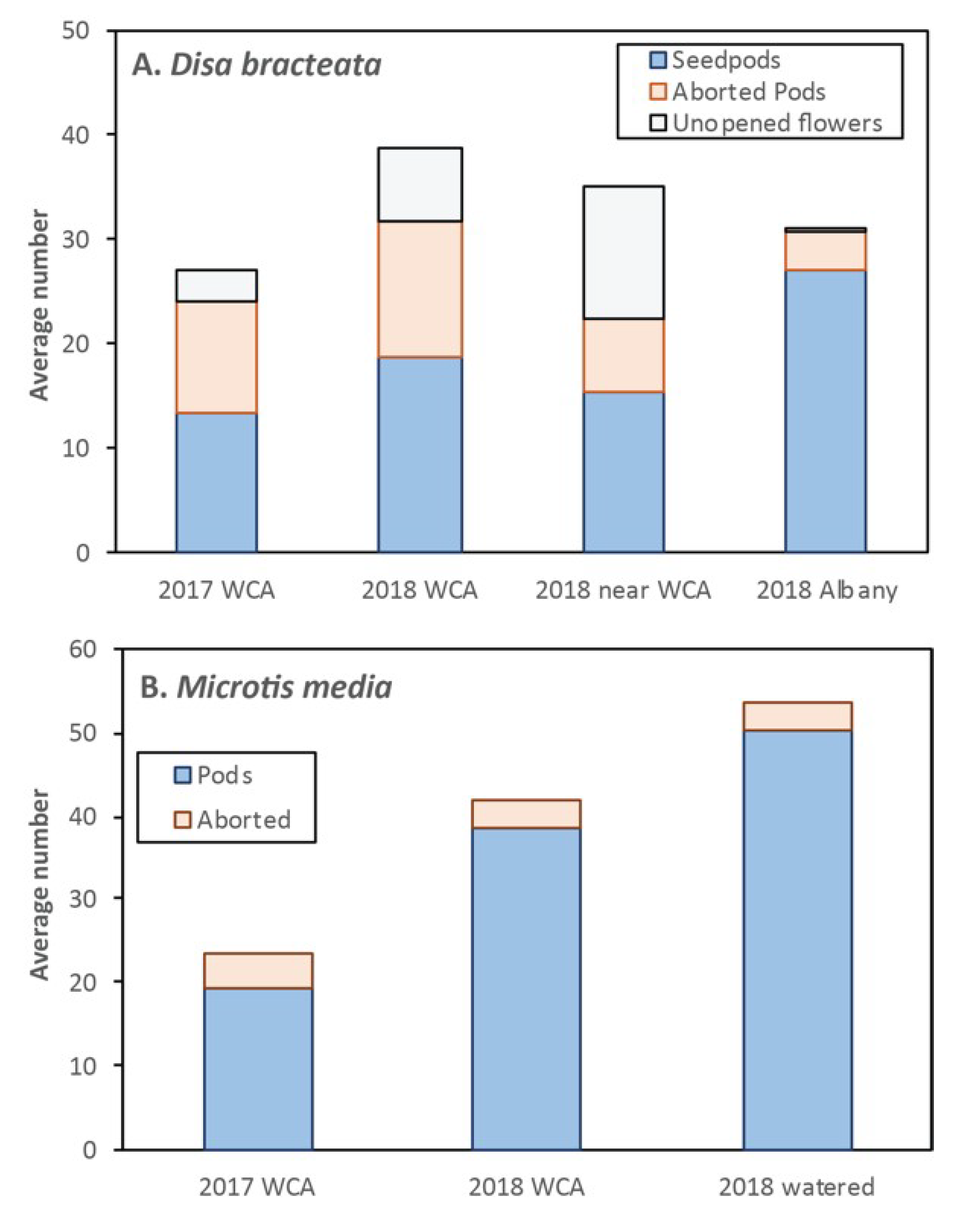
3.18. Disa bracteata (South African Orchid)
3.19. Microtis media (Mignonette Orchid)
3.20. Thelymitra fuscolutea (Chestnut Sun Orchid)
3.21. Other less Common Orchids
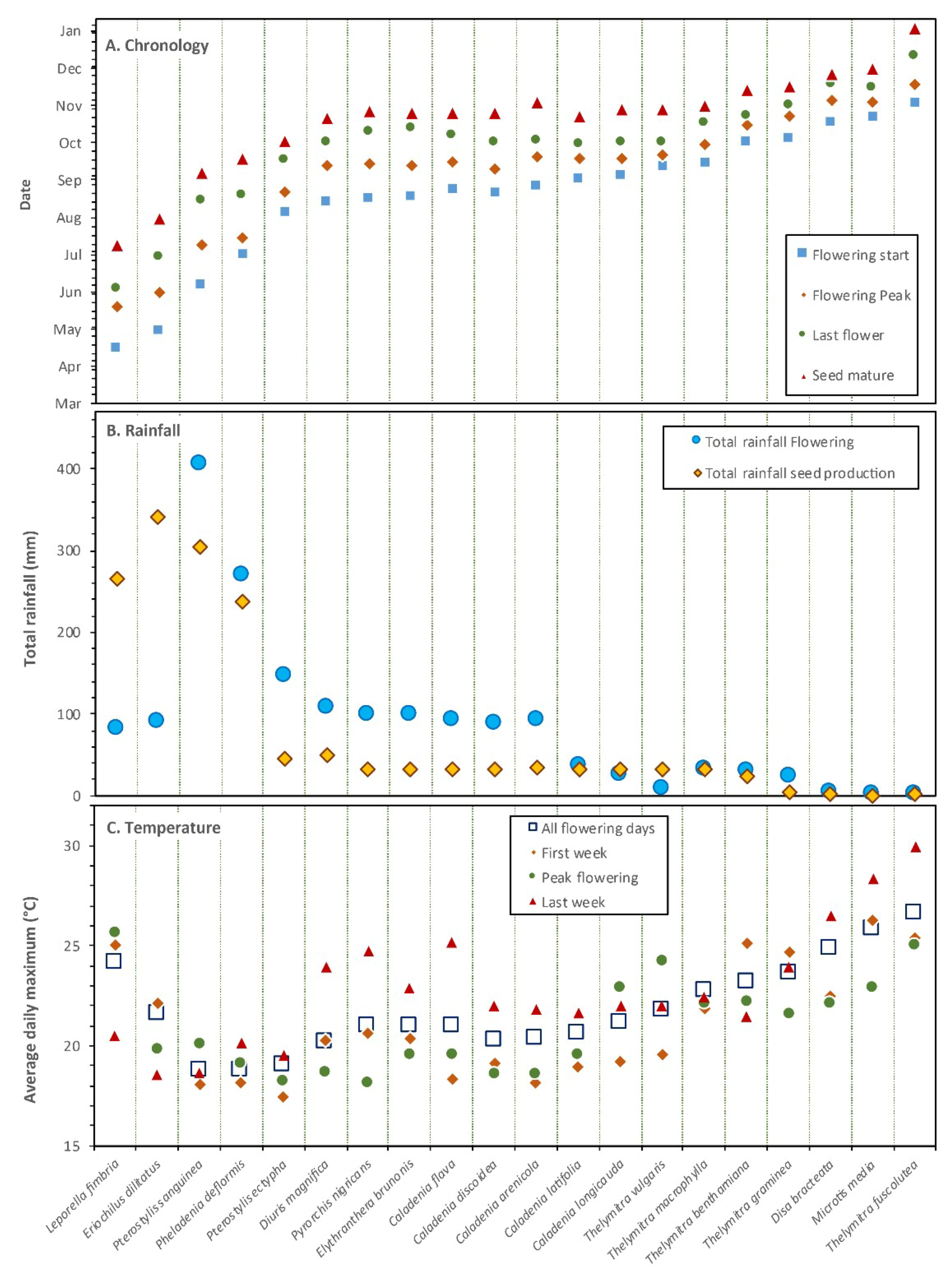
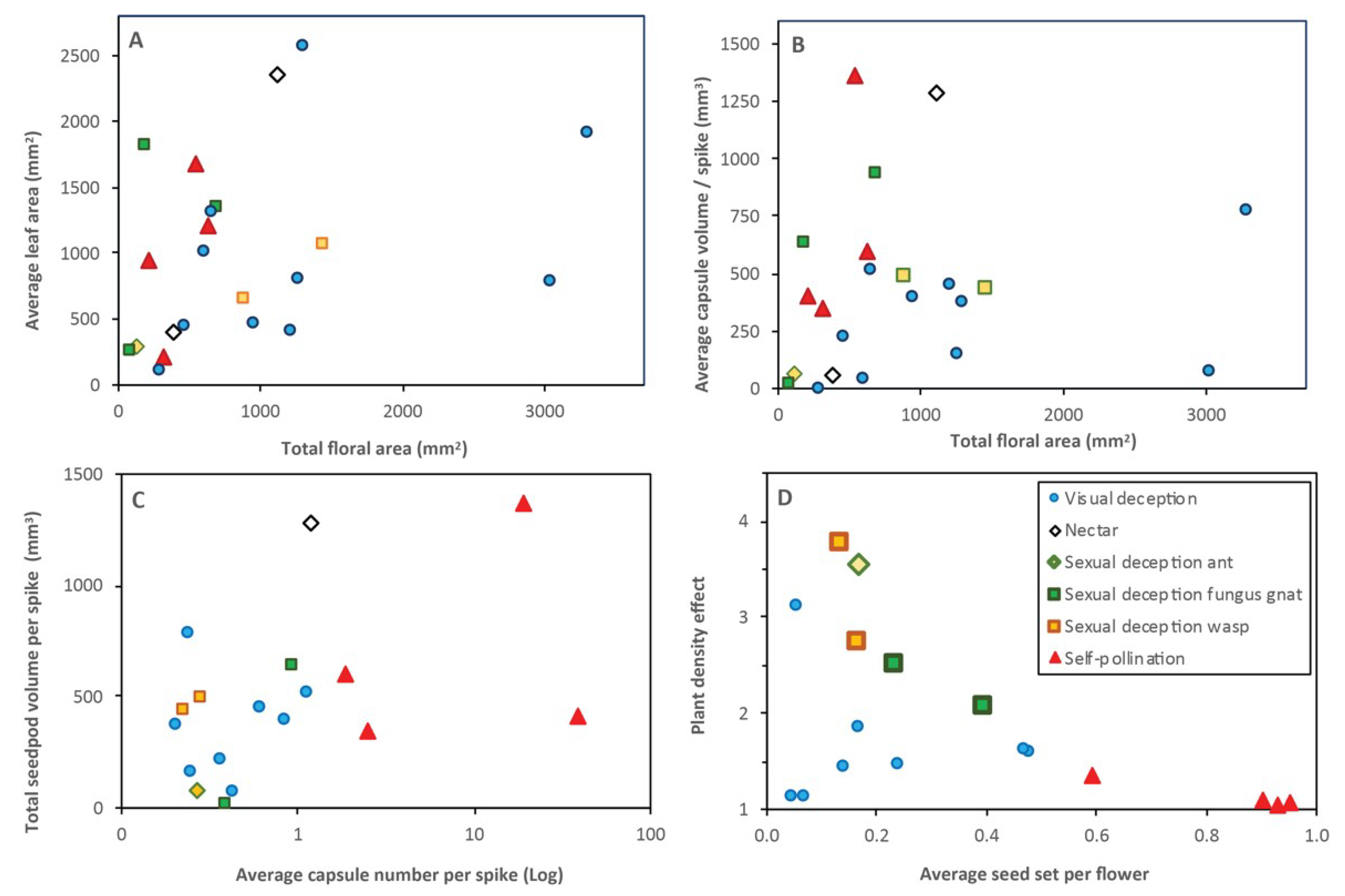
3.22. Pollination Outcomes Relative to Climate and Ecology
4. Discussion
4.1. Climate and Habitat Conditions
4.2. Plant Density Effects and Resource Limitations
4.3. Pollination Outcomes Relative to Orchid Ecology and Evolution
5. Conclusions
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Full Name | Flowering (fls.) Start (date) | Fls. Peak (date) | Fls. End (date) | Seed Ripe (date) | Spike Height (mm) | Leaf no. | Leaf Length (mm) | Leaf Width (mm) | Leaf Area mm3 | Plants With Fls. | Mean Fls. per Spike | Fl. no. Range | Fl. Area (mm3) | Fl. Area per Spike | Fl. Area per Plant (mm3) | Flower Colors | Pollination Syndrome | Observed and Reported Pollinators | References for Pollinators |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Leporella fimbriata (Lindl.) A.S.George | 16–4 | 20–5 | 4–6 | 10–7 | 180 | 1–2 | 16–20 | 4–6 | 300 | 0.09 | 1.6 | 1–3 | 70 | 112 | 9.7 | Green and purple | Sexual deception (SD) | Male ants (Myrmecia spp.) | [55,86] |
| Eriochilus dilatatus Lindl. subsp. dilatatus | 1–5 | 30–5 | 1–7 | 1–8 | 320 | 1 | 10–70 | 4–8 | 400 | 0.77 | 2.4 | 1–8 | 160 | 384 | 295 | White with pink and green markings | Nectar | Native bees, honeybees | [32,44,55] |
| Pterostylis sanguinea D.L.Jones and M.A.Clem. | 7–6 | 11–7 | 16–8 | 9–9 | 210 | 7–13 | 4–14 | 3–6 | 1800 | 0.95 | 3.9 | 1–15 | 50 | 195 | 185.25 | Translucent green, red or brown | SD | Fungus gnats (Mycomya sp.) | [55,88] |
| Pheladenia deformis (R.Br.) D.L.Jones and M.A.Clem. | 2–7 | 17–7 | 7–8 | 20–9 | 130 | 1 | 45–70 | 1–3 | 90 | 0.83 | 1 | 1–2 | 300 | 300 | 249 | Bright blue | Visual deception (VD) | Native bees | [48] |
| Pterostylis ectypha D.L.Jones and C.J.French | 7–8 | 23–8 | 19–9 | 5–10 | 80 | 3–7 | 11–19 | 4–7 | 210 | 0.65 | 1 | 1 | 90 | 90 | 58.5 | Green | SD | Fungus gnats | [55,89] |
| Diuris magnifica D.L.Jones | 15–8 | 15–9 | 4–10 | 24–10 | 480 | 2–5 | 60–280 | 5–10 | 2560 | 0.95 | 3.6 | 2–8 | 360 | 1296 | 1231.2 | Yellow, orange, purple | VD | Moth, bees, flies, beetles | [17,22] |
| Pyrorchis nigricans (R.Br.) D.L.Jones and M.A.Clem. | 19–8 | 16–9 | 12–10 | 30–10 | 180 | 1–2 | 40–100 | 30–80 | 2350 | 0.3 | 3.7 | 2–7 | 300 | 1110 | 333 | Dark red | Some nectar | Bees, flies, etc. | [55] |
| Elythranthera brunonis (Endl.) A.S.George | 20–8 | 15–9 | 16–10 | 24–10 | 330 | 1 | 60–145 | 5–7 | 440 | 0.71 | 1.6 | 1–3 | 290 | 464 | 329.44 | Glossy dark purple | VD | Bees, flies, etc. | [32,64] |
| Caladenia flava R.Br. subsp. flava | 26–8 | 18–9 | 10–10 | 29–10 | 150 | 1 | 50–100 | 5–12 | 450 | 0.24 | 1.8 | 1–4 | 530 | 954 | 228.96 | Bright yellow | VD | Native bees, Beetles | [32,44] |
| Caladenia discoidea Lindl. | 23–8 | 11–9 | 4–10 | 28–10 | 230 | 1 | 75–150 | 8–5 | 650 | 0.62 | 1.8 | 1–4 | 490 | 882 | 546.84 | Orange-green | SD | Thynnine wasp | [45] |
| Caladenia arenicola Hopper and A.P.Br. | 28–8 | 22–9 | 5–10 | 6–11 | 440 | 1 | 100–225 | 8–12 | 1050 | 0.60 | 1.8 | 1–4 | 805 | 1449 | 869.4 | Red and green | SD | Thynnid wasp | [42,77] |
| Caladenia latifolia R.Br. | 4–9 | 20–9 | 24–10 | 7–11 | 300 | 1 | 65–210 | 8–21 | 1300 | 0.64 | 2.5 | 1–5 | 260 | 650 | 416 | Pink | VD | Beetles, bees | See C. flava |
| Caladenia longicauda subsp. calcigena Hopper and A.P.Br. | 7–9 | 20–9 | 4–10 | 1–11 | 320 | 1 | 120–200 | 7–12 | 800 | 0.25 | 1.8 | 1–4 | 700 | 1260 | 315 | White | VD | Flies, beetles, bees | [32,42] |
| Thelymitra vulgaris Jeanes | 14–9 | 23–9 | 4–10 | 1–11 | 315 | 1 | 65–155 | 1–4 | 210 | 0.9 | 2.6 | 1–6 | 120 | 312 | 280.8 | Blue | Self-pollination | none observed | [90] |
| Thelymitra macrophylla Lindl. | 17–9 | 2–10 | 20–10 | 4–11 | 700 | 1 | 140–250 | 7–25 | 1900 | 0.74 | 6.6 | 2–15 | 500 | 3300 | 2442 | Blue | VD | Flies, bees, etc. | [70] |
| Thelymitra benthamiana Rchb.f. | 4–10 | 14–10 | 26–10 | 16–11 | 330 | 1 | 40–210 | 10–40 | 1200 | 0.50 | 2.1 | 1–8 | 300 | 630 | 315 | Yellow and brown | Self-pollination | none observed | [91] |
| Thelymitra graminea Lindl. | 7–10 | 26–10 | 4–11 | 20–11 | 360 | 1 | 55–170 | 8–3 | 400 | 0.97 | 3.8 | 2–8 | 320 | 1216 | 1179.52 | Blue | VD | Hoverflies, etc. | This study |
| Disa bracteata Sw. | 20–10 | 8–11 | 21–11 | 30–11 | 240 | 5–16 | 15–150 | 2–10 | 1670 | 0.95 | 32 | 10–70 | 17 | 544 | 516.8 | Greenish | Self-pollination (some nectar) | none observed | [46] |
| Microtis media R.Br. subsp. media | 25–10 | 7–11 | 18–11 | 25–11 | 250 | 1 | 140–270 | 1.8–2.2 | 950 | 1 | 42 | 10–100 | 5 | 210 | 210 | Green | Self-pollination (some nectar) | ants, micro wasps | [55,92] |
| Thelymitra fuscolutea R.Br. | 6–11 | 21–11 | 14–12 | 7–1 | 380 | 1 | 60–140 | 7–15 | 780 | 0.72 | 6.2 | 2–10 | 490 | 3038 | 2187.36 | Orange and brown | VD | Hover flies, etc. | This study |
| Orchid | Location | Year | Spikes | Flowers | Capsules/Flower | Capsules/Spike | Capsules/Flower SGA | Capsules/Spike SGA | Capsules/Flower LGA | Capsules Measured | Capsules/Spike Range | Average Capsule Length (mm) | Capsule Length Range (mm) | Average Capsule Width (mm) | Capsule Width Range (mm) | Average Capsule Volume (mm3) | Capsule Volume Range | Total Capsule Volume/Spike (mm3) | Total Capsule Volume/Plant (mm3) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Leporella fimbriata | WCA | 2018 | 175 | 278 | 0.17 | 0.27 | 0.60 | 1.2 | 0.2 | 21 | 0–3 | 8.7 | 6–11 | 3 | 2–5 | 45 | 30–90 | 72 | 6 |
| Eriochilus dilatatus | WCA, etc. | 2014–16 | 61 | 149 | 0.16 | 0.59 | ND | ND | ND | 3 | 0–4 | 13 | 11–15 | 4 | 3–4 | 100 | 70–120 | 59 | 45 |
| Pterostylis sanguinea | WCA | 2018 | 173 | 680 | 0.23 | 0.95 | 0.58 | 2.5 | 0.1 | 61 | 0–6 | 13 | 9–16 | 7 | 5–10 | 300 | 100–690 | 630 | 599 |
| Pheladenia deformis | WCA | 2018 | 39 | 39 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 18 | 15–20 | 4.5 | 4–5 | 190 | NA | 0 | 0 |
| Pterostylis ectypha | WCA | 2018 | 265 | 265 | 0.39 | 0.39 | 0.81 | 0.81 | 0.2 | 26 | 0–1 | 11 | 7–20 | 3.5 | 3–5 | 80 | 35–290 | 14 | 9 |
| Pterostylis recurva* | WCA, etc. | 2016–18 | 4 | 8 | 0.5 | 1 | ND | ND | ND | 4 | 0–2 | 16 | 15–19 | 9 | 7.5–10 | 720 | 500–1000 | 936 | 936 |
| Diuris magnifica | WCA | 2018 | 465 | 1690 | 0.058 | 0.21 | 0.18 | 0.76 | 0.00 | 46 | 0–3 | 13 | 12–17 | 7 | 5–8 | 320 | 230–640 | 368 | 350 |
| Pyrorchis nigricans | Marang-aroo | 2018 | 18 | 67 | 0.31 | 1.17 | ND | ND | ND | 9 | 2–3 | 14.5 | 10–19 | 8.2 | 6.8–11.8 | 570 | 220–1400 | 1283 | 385 |
| Elythranthera brunonis | WCA | 2016–18 | 35 | 55 | 0.24 | 0.37 | ND | ND | ND | 6 | 0–2 | 12.2 | 9.1–14.1 | 5.3 | 4.8–6.4 | 180 | 112–246 | 216 | 153 |
| Caladenia flava | WCA | 2018 | 34 | 61 | 0.48 | 0.85 | 0.76 | 1.4 | 0.05 | 10 | 0–2 | 15 | 13–18 | 6 | 4–6.5 | 280 | 130–400 | 392 | 94 |
| Caladenia arenicola | WCA | 2008–18 | 101 | 177 | 0.13 | 0.23 | 0.49 | 0.78 | 0.00 | 12 | 0–2 | 14.9 | 10–20 | 6.7 | 5–9 | 360 | 170–650 | 432 | 259 |
| Caladenia discoidea | WCA | 2015–18 | 52 | 95 | 0.16 | 0.29 | 0.44 | 0.44 | 0.15 | 7 | 0–2 | 15 | 10–21 | 5.8 | 3–9 | 350 | 40–580 | 490 | 304 |
| Caladenia latifolia | WCA | 2018 | 105 | 217 | 0.47 | 1.16 | 0.75 | 2.3 | 0.3 | 16 | 0–2 | 16 | 11–20 | 6 | 4–7.5 | 320 | 80–510 | 512 | 328 |
| Caladenia longicauda | Sorento | 2008–18 | 4 | 7 | 0.14 | 0.25 | ND | ND | ND | 2 | 0–1 | 20 | NA | 7 | NA | 520 | NA | 151 | 38 |
| Leptoceras menziesii* | WCA | 2008 | 6 | 10 | 0.3 | 0.5 | ND | ND | ND | 3 | 0–2 | 9.5 | 9–10 | 4 | 3–5 | 80 | 60–120 | 40 | 12 |
| Thelymitra vulgaris | WCA | 2018 | 29 | 76 | 0.95 | 2.48 | ND | ND | ND | 65 | 0–5 | 10.5 | 8–19 | 4.4 | 3–5.5 | 133 | 30–290 | 346 | 311 |
| Thelymitra macrophylla | WCA | 2018 | 698 | 4574 | 0.048 | 0.24 | 0.054 | 1.2 | 0.04 | 171 | 0–8 | 13 | 21–7 | 5.5 | 3.5–9 | 215 | 40–820 | 774 | 573 |
| Thelymitra benthamiana | WCA | 2018 | 38 | 79 | 0.90 | 1.87 | 0.99 | 2.6 | 0.90 | 36 | 0–5 | 15 | 7–22 | 6 | 4–8 | 300 | 90–660 | 600 | 300 |
| Thelymitra graminea | WCA | 2018 | 64 | 240 | 0.17 | 0.63 | 0.31 | 0.6 | 0.13 | 29 | 0–5 | 10.5 | 17–6 | 4.7 | 7.5–3.5 | 140 | 30–360 | 448 | 435 |
| Disa bracteata | WCA | 2018 | 26 | 821 | 0.59 | 19 | ND | ND | ND | 58 | 3–46 | 11 | 18–7 | 3.4 | 2–4.5 | 72 | 18–138 | 1368 | 1300 |
| Microtis media | WCA | 2018 | 38 | 1583 | 0.93 | 39 | 0.97 | 42 | 0.92 | 96 | 30–67 | 4.1 | 5.7–2.8 | 2.4 | 3.7–1.5 | 14 | 5–40 | 406 | 406 |
| Thelymitra fuscolutea | WCA | 2018 | 102 | 632 | 0.070 | 0.43 | 0.078 | 0.54 | 0.06 | 78 | 0–4 | 11 | 8–14 | 5.2 | 4–7 | 160 | 70–310 | 67 | 48 |
References
- Neiland, M.R.M.; Wilcock, C.C. Fruit set, nectar reward, and rarity in the Orchidaceae. Am. J. Bot. 1998, 85, 1657–1671. [Google Scholar] [CrossRef] [PubMed]
- Arditti, J.; Ghani, A.K.A. Numerical and physical properties of orchid seeds and their biological implications. New Phytol. 2000, 145, 367–421. [Google Scholar] [CrossRef]
- Brundrett, M.C. Scientific approaches to Australian temperate terrestrial orchid conservation. Aust. J. Bot. 2007, 55, 293–307. [Google Scholar] [CrossRef]
- McCormick, M.K.; Jacquemyn, H. What constrains the distribution of orchid populations? New Phytol. 2014, 202, 392–400. [Google Scholar] [CrossRef]
- Phillips, R.D.; Peakall, R.; Hutchinson, M.F.; Linde, C.C.; Xu, T.; Dixon, K.W.; Hopper, S.D. Specialized ecological interactions and plant species rarity: the role of pollinators and mycorrhizal fungi across multiple spatial scales. Biol. Conserv. 2014, 169, 285–295. [Google Scholar] [CrossRef]
- Darwin, C. The Various Contrivances by which British and Foreign Orchids are Fertilised By Insects, 2nd ed.; John Murray: London, UK, 1877; pp. 1–300. [Google Scholar]
- Johnson, S.D.; Nilsson, L.A. Pollen carryover, geitonogamy, and the evolution of deceptive pollination systems in orchids. Ecology 1999, 80, 2607–2619. [Google Scholar] [CrossRef]
- Weston, P.H.; Perkins, A.J.; Indsto, J.O.; Clements, M.A. Phylogeny of Orchidaceae tribe Diurideae and its implications for the evolution of pollination systems. In Darwin’s Orchids: Then and Now; University of Chicago Press: Chicago, IL, USA, 2014; pp. 91–154. [Google Scholar]
- Dafni, A. Mimicry and deception in pollination. Annu. Rev. Ecol. Syst. 1984, 15, 259–278. [Google Scholar] [CrossRef]
- Cozzolino, S.; Widmer, A. Orchid diversity: an evolutionary consequence of deception? Trends Ecol. Evol. 2005, 20, 487–494. [Google Scholar] [CrossRef]
- Vereecken, N.J.; Dafni, A.; Cozzolino, S. Pollination syndromes in Mediterranean orchids—implications for speciation, taxonomy and conservation. Bot. Rev. 2010, 76, 220–240. [Google Scholar] [CrossRef]
- Schiestl, F.P. On the success of a swindle: Pollination by deception in orchids. Naturwissenschaften 2005, 92, 255–264. [Google Scholar] [CrossRef]
- Tremblay, R.L.; Ackerman, J.D.; Zimmerman, J.K.; Calvo, R.N. Variation in sexual reproduction in orchids and its evolutionary consequences: a spasmodic journey to diversification. Biol. J. Linn. Soc. 2005, 84, 1–54. [Google Scholar] [CrossRef]
- Jersáková, J.; Johnson, S.D.; Kindlmann, P. Mechanisms and evolution of deceptive pollination in orchids. Biol. Rev. 2006, 81, 219–235. [Google Scholar] [CrossRef] [PubMed]
- Dafni, A.; Bernhhardt, P. Pollination of terrestrial orchids of Southern Australia and the Mediterranean region. Systematics, ecological and evolutionary implications. Evol. Biol. 1990, 24, 193–252. [Google Scholar]
- Herberstein, M.E.; Baldwin, H.J.; Gaskett, A.C. Deception down under: is Australia a hot spot for deception? Behav. Ecol. 2013, 25, 12–16. [Google Scholar] [CrossRef] [Green Version]
- Beardsell, D.; Clements, M.; Hutchinson, J.; Williams, E. Pollination of Diuris maculata R Br (Orchidaceae) by floral mimicry of the native legumes Daviesia spp. and Pultenaea scabra R Br. Aust. J. Bot. 1986, 34, 165–173. [Google Scholar] [CrossRef]
- Elliott, C.P.; Ladd, P.G. Pollen limitation of fruit set in Western Australian terrestrial orchids. J. R. Soc. West. Aust. 2002, 85, 165–168. [Google Scholar]
- Edens-Meier, R.; Bernhardt, P. The Sun Orchids (Thelymitra) Then and Now: Large Flowers versus Small Flowers and Their Evolutionary Implications. Darwins Orchids Then Now 2014, 173–200. [Google Scholar]
- Gaskett, A.C.; Endler, J.A.; Phillips, R.D. Convergent evolution of sexual deception via chromatic and achromatic contrast rather than colour mimicry. Evol. Ecol. 2017, 31, 205–227. [Google Scholar] [CrossRef]
- Cropper, S.C.; Calder, D.M. The floral biology of Thelymitra epipactoides (Orchidaceae), and the implications of pollination by deceit on the survival of this rare orchid. Plant Syst. Evol. 1990, 170, 11–27. [Google Scholar] [CrossRef]
- Indsto, J.O.; Weston, P.H.; Clements, M.A.; Dyer, A.G.; Batley, M.; Whelan, R.J. Pollination of Diuris maculata (Orchidaceae) by male Trichocolletes venustus bees. Aust. J. Bot. 2006, 54, 669–679. [Google Scholar] [CrossRef]
- Scaccabarozzi, D.; Cozzolino, S.; Guzzetti, L.; Galimberti, A.; Milne, L.; Dixon, K.W.; Phillips, R.D. Masquerading as pea plants: behavioural and morphological evidence for mimicry of multiple models in an Australian orchid. Ann. Bot. 2018, 122, 1061–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kagawa, K.; Takimoto, G. Inaccurate color discrimination by pollinators promotes evolution of discrete color polymorphism in food-deceptive flowers. Am. Nat. 2016, 187, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Adams, P.B.; Lawson, S.D. Pollination in Australian orchids: a critical–assessment of the literature 1882–1992. Aust. J. Bot. 1993, 41, 553–575. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Hopper, S.D.; Gioia, P. The southwest Australian floristic region: evolution and conservation of a global hot spot of biodiversity. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 623–650. [Google Scholar] [CrossRef]
- Lambers, H.; Brundrett, M.C.; Raven, J.A.; Hopper, S.D. Plant mineral nutrition in ancient landscapes: high plant species diversity on infertile soils is linked to functional diversity for nutritional strategies. Plant Soil 2011, 348, 7. [Google Scholar] [CrossRef]
- Groom, P.K.; Lamont, B. Plant Life of Southwestern Australia: Adaptations for Survival; Walter de Gruyter GmbH & Co KG: Berlin, Germany, 2015; pp. 1–258. [Google Scholar]
- Brundrett, M.C. Distribution and evolution of mycorrhizal types and other specialised roots in Australia. In Biogeography of Mycorrhizal Symbiosis; Springer: Cham, Switzerland, 2017; Volume 230, pp. 361–394. [Google Scholar]
- Phillips, R.D.; Barrett, M.D.; Dixon, K.W.; Hopper, S.D. Do mycorrhizal symbioses cause rarity in orchids? J. Ecol. 2011, 99, 858–869. [Google Scholar] [CrossRef]
- Brundrett, M. Identification and Ecology of Southwest Australian Orchids; Western Australian Naturalists’ Club Inc.: Perth, WA, Australia, 2014; p. 424. [Google Scholar]
- Banksia Woodlands of the Swan Coastal Plain: a nationally-protected ecological community. Available online: https://www.environment.gov.au/biodiversity/threatened/publications/banksia-woodlands-swan-coastal-plain-guide (accessed on 16 September 2016).
- Ramalho, C.E.; Laliberte, E.; Poot, P.; Hobbs, R.J. Complex effects of fragmentation on remnant woodland plant communities of a rapidly urbanizing biodiversity hotspot. Ecology 2014, 95, 2466–2478. [Google Scholar] [CrossRef] [Green Version]
- Government of Western Australia Directory of Bush Forever Sites. In Bush Forever: Keeping the Bush in the City; Department of Environmental Protection: Perth, Western Australia, Australia, 2000; Volume 2, pp. 1–530.
- Friends of Warwick Bushland Warwick Bushland. Available online: Friendsofwarwickbushland.com (accessed on 24 June 2019).
- Bonnardeaux, Y.; Brundrett, M.; Batty, A.; Dixon, K.; Koch, J.; Sivasithamparam, K. Diversity of mycorrhizal fungi of terrestrial orchids: compatibility webs, brief encounters, lasting relationships and alien invasions. Mycol. Res. 2007, 111, 51–61. [Google Scholar] [CrossRef]
- Scade, A.; Brundrett, M.; Batty, A.; Dixon, K.; Sivasithamparam, K. Survival of transplanted terrestrial orchid seedlings in urban bushland habitats with high or low weed cover. Aust. J. Bot. 2006, 54, 383–389. [Google Scholar] [CrossRef]
- Tao, Z.-B.; Ren, Z.-X.; Bernhardt, P.; Wang, W.-J.; Liang, H.; Li, H.-D.; Wang, H. Nocturnal hawkmoth and noctuid moth pollination of Habenaria limprichtii (Orchidaceae) in sub-alpine meadows of the Yulong Snow Mountain (Yunnan, China). Bot. J. Linn. Soc. 2018, 187, 483–498. [Google Scholar] [CrossRef]
- Petit, S.; Jusaitis, M.; Bickerton, D. Effect of pollen load, self-pollination and plant size on seeds and germination in the endangered pink-lipped spider orchid, Caladenia behrii. Aust. J. Bot. 2009, 57, 307–314. [Google Scholar] [CrossRef]
- Bureau of Meteorology Climate data online. Available online: www.bom.gov.au (accessed on 18 January 2019).
- Stoutamire, W.P. Wasp-pollinated species of Caladenia (Orchidaceae) in south-western Australia. Aust. J. Bot. 1983, 31, 383–394. [Google Scholar] [CrossRef]
- Phillips, R.; Peakall, R.; Retter, B.; Montgomery, K.; Menz, M.; Davis, B.; Hayes, C.; Brown, G.; Swarts, N.; Dixon, K. Pollinator rarity as a threat to a plant with a specialized pollination system. Bot. J. Linn. Soc. 2015, 179, 511–525. [Google Scholar] [CrossRef] [Green Version]
- Erickson, R. Orchids of the west; Paterson, Brokensha, Pty. Ltd.: Perth, Western Australia, Australia, 1951; pp. 1–109. [Google Scholar]
- Pollinator Photos by Mark Brundrett & Orchid Photographers Identifications by Graham Brown. Available online: https://www.researchgate.net/publication/269280032_Wasp_Pollinator_Photos_By_Mark_Brundrett_Orchid_Photographers_Identifications_by_Graham_Brown (accessed on 9 December 2014).
- Johnson, S.D.; Linder, H.P.; Steiner, K.E. Phylogeny and radiation of pollination systems in Disa (Orchidaceae). Am. J. Bot. 1998, 85, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Light, M.H.S.; MacConaill, M. Climatic influences on flowering and fruiting of Cypripedium parviflorum var. pubescens. In Trends and Fluctuations and Underlying Mechanisms in Terrestrial Orchid Populations; Backhuys Publishers: Leiden, Netherland, 2002; pp. 85–97. [Google Scholar]
- Rogers, R.S. Pollination of Caladenia deformis, R. Br. Trans. Proc. R. Soc. S. Aust. 1931, 55, 143–146. [Google Scholar]
- Brown, J.; York, A. Fire, food and sexual deception in the neighbourhood of some Australian orchids. Austral Ecol. 2017, 42, 468–478. [Google Scholar] [CrossRef]
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PloS One 2017, 12, e0185809. [Google Scholar] [CrossRef]
- Ridsdill Smith, T. The behaviour of Hemithynnus hyalinatus (Hymenoptera: Tiphiidae), with notes on some other Thynninae. Aust. J. Entomol. 1970, 9, 196–208. [Google Scholar]
- Menz, M.H.; Phillips, R.D.; Dixon, K.W.; Peakall, R.; Didham, R.K. Mate-searching behaviour of common and rare wasps and the implications for pollen movement of the sexually deceptive orchids they pollinate. PLoS One 2013, 8, e59111. [Google Scholar] [CrossRef]
- Jersáková, J.; Malinová, T. Spatial aspects of seed dispersal and seedling recruitment in orchids. New Phytol. 2007, 176, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Woodland Restoration Project Annual Report 6 January - December 2017. Available online: https://www.researchgate.net/publication/325965309_Banksia_Woodland_Restoration_Project_Annual_Report_6_January_-_December_2017 (accessed on 25 June 2018).
- Kuiter, R.H. Orchid Pollinators of Victoria, 3rd ed.; Aquatic Photographics: Seaford, Victoria, VIC, Australia, 2015; pp. 252–525. [Google Scholar]
- Brundrett, M.C. Using vital statistics and core-habitat maps to manage critically endangered orchids in the Western Australian wheatbelt. Aust. J. Bot. 2016, 64, 51–64. [Google Scholar] [CrossRef]
- Petit, S.; Dickson, C.R. Grass-tree (Xanthorrhoea semiplana, Liliaceae) facilitation of the endangered pink-lipped spider orchid (Caladenia syn. Arachnorchis behrii, Orchidaceae) varies in South Australia. Aust. J. Bot. 2005, 53, 455–464. [Google Scholar] [CrossRef]
- Ong, J.; Li, H.; Sivasithamparam, K.; Dixon, K.W.; Jones, M.; Wylie, S.J. Novel and divergent viruses associated with Australian orchid-fungus symbioses. Virus Res. 2018, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.; Brundrett, M. Recovery of terrestrial orchids in natural ecosystems after severe disturbance. In Mining in Ecologically Sensitive Landscapes; CSIRO PUBLISHING: Victoria, Australia, 2015; pp. 141–158. [Google Scholar]
- Brundrett, M.; Scade, A.; Batty, A.; Dixon, K.; Sivasithamparam, K. Development of in situ and ex situ seed baiting techniques to detect mycorrhizal fungi from terrestrial orchid habitats. Mycol. Res. 2003, 107, 1210–1220. [Google Scholar] [CrossRef] [PubMed]
- Jacquemyn, H.; Brys, R.; Hermy, M.; Willems, J.H. Does nectar reward affect rarity and extinction probabilities of orchid species? An assessment using historical records from Belgium and the Netherlands. Biol. Conserv. 2005, 121, 257–263. [Google Scholar] [CrossRef]
- Zimmerman, J.K.; Aide, T.M. Patterns of fruit production in a neotropical orchid: Pollinator vs. resource limitation. Am. J. Bot. 1989, 76, 67–73. [Google Scholar] [CrossRef]
- Vallius, E. Position-dependent reproductive success of flowers in Dactylorhiza maculata (Orchidaceae). Funct. Ecol. 2000, 14, 573–579. [Google Scholar] [CrossRef]
- Tremblay, R.L.; Bateman, A.P.; Brown, A.; Hachandourian, M.; Hutchings, M.J.; Kell, S.; Koopowitz, H.; Lehnebach, C.; Whigham, D.F. Density induced rates of pollinaria removal and deposition in the purple enamel orchid Elythranthera brunonis (Endl.). AS George. 2007, 7, 229–239. [Google Scholar] [CrossRef]
- Peakall, R. Responses of male Zaspilothynnus trilobatus Turner wasps to females and the sexually deceptive orchid it pollinates. Funct. Ecol. 1990, 159–167. [Google Scholar] [CrossRef]
- Dafni, A.; Calder, D. Pollination by deceit and floral mimesis in Thelymitra antennifera (Orchidaceae). Plant Syst. Evol. 1987, 158, 11–22. [Google Scholar] [CrossRef]
- Jacquemyn, H.; Brys, R.; Honnay, O. Large population sizes mitigate negative effects of variable weather conditions on fruit set in two spring woodland orchids. Biol. Lett. 2009, 5, 495–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brys, R.; Jacquemyn, H.; Hermy, M. Pollination efficiency and reproductive patterns in relation to local plant density, population size, and floral display in the rewarding Listera ovata (Orchidaceae). Bot. J. Linn. Soc. 2008, 157, 713–721. [Google Scholar] [CrossRef]
- Perez-Hernandez, H.; Damon, A.; Valle-Mora, J.; Sanchez-Guillen, D. Orchid pollination: specialization in chance? Bot. J. Linn. Soc. 2011, 165, 251–266. [Google Scholar] [CrossRef]
- Edens-Meier, R.M.; Raguso, R.A.; Westhus, E.; Bernhardt, P. Floral fraudulence: Do blue Thelymitra species (Orchidaceae) mimic Orthrosanthus laxus (Iridaceae)? Telopea 2014, 17, 15–28. [Google Scholar] [CrossRef]
- Western Australian Local Government Association 2013 Native vegetation on the Swan Coastal Plain. Available online: http://pbp.walga.asn.au/Publications.html (accessed on 2 April 2019).
- Ackerman, J.D.; Cuevas, A.A.; Hof, D. Are deception-pollinated species more variable than those offering a reward? Plant Syst. Evol. 2011, 293, 91–99. [Google Scholar] [CrossRef]
- Sonkoly, J.E.; Vojtkó, A.; Tökölyi, J.; Török, P.; Sramkó, G.; Illyés, Z.; Molnár, V.A. Higher seed number compensates for lower fruit set in deceptive orchids. J. Ecol. 2016, 104, 343–351. [Google Scholar] [CrossRef]
- Calvo, R.N. Evolutionary demography of orchids: intensity and frequency of pollination and the cost of fruiting. Ecology 1993, 74, 1033–1042. [Google Scholar] [CrossRef]
- Givnish, T.J.; Spalink, D.; Ames, M.; Lyon, S.P.; Hunter, S.J.; Zuluaga, A.; Iles, W.J.; Clements, M.A.; Arroyo, M.T.; Leebens-Mack, J. Orchid phylogenomics and multiple drivers of their extraordinary diversification. Proc R Soc B 2015, 282, 20151553. [Google Scholar] [CrossRef]
- Nauheimer, L.; Schley, R.J.; Clements, M.A.; Micheneau, C.; Nargar, K. Australasian orchid biogeography at continental scale: molecular phylogenetic insights from the sun orchids (Thelymitra, Orchidaceae). Mol. Phylogenet. Evol. 2018, 127, 304–319. [Google Scholar] [CrossRef]
- Phillips, R.D.; Faast, R.; Bower, C.C.; Brown, G.R.; Peakall, R. Implications of pollination by food and sexual deception for pollinator specificity, fruit set, population genetics and conservation of Caladenia (Orchidaceae). Aust. J. Bot. 2009, 57, 287–306. [Google Scholar] [CrossRef]
- Overview of Pterostylis Pollination (Orchidaceae) in Victoria. Available online: https://www.researchgate.net/publication/318282443_Overview_of_Pterostylis_Pollination_Orchidaceae_in_Victoria (accessed on 8 July 2017).
- Kearns, C.A.; Inouye, D.W.; Waser, N.M. Endangered mutualisms: The conservation of plant-pollinator interactions. Annu. Rev. Ecol. Syst. 1998, 29, 83–112. [Google Scholar] [CrossRef]
- Williams, M.R. Habitat resources, remnant vegetation condition and area determine distribution patterns and abundance of butterflies and day-flying moths in a fragmented urban landscape, south-west Western Australia. J. Insect Conserv. 2011, 15, 37–54. [Google Scholar] [CrossRef]
- Light, M.H.; MacConaill, M. The role of common orchids in appreciating the complexity of biodiversity conservation. Lankesteriana Int. J. Orchid. 2011, 11, 293–330. [Google Scholar] [CrossRef]
- Catling, P.M. Auto-pollination in the Orchidaceae. Timber Press 1990, 5, 121–158. [Google Scholar]
- Newman, B.J.; Ladd, P.; Brundrett, M.; Dixon, K.W. Effects of habitat fragmentation on plant reproductive success and population viability at the landscape and habitat scale. Biol. Conserv. 2013, 159, 16–23. [Google Scholar] [CrossRef]
- Orchid Rescue Project: Case Studies of Collaborative Orchid Conservation in Western Australia; University of Western Australia. Available online: https://www.researchgate.net/publication/259500490_Wheatbelt_Orchid_Rescue_Project_Case_Studies_of_Collaborative_Orchid_Conservation_in_Western_Australia (accessed on 2 January 2011).
- Sydes, M.A.; Calder, D.M. Comparative reproductive biology of two sun-orchids; the vulnerable Thelymitra circumsepta and the widespread T. ixioides (Orchidaceae). Aust. J. Bot. 1993, 41, 577–589. [Google Scholar] [CrossRef]
- Peakall, R. The unique pollination of Leporella fimbriata (Orchidaceae): pollination by pseudocopulating male ants (Myrmecia urens, Formicidae). Plant Syst. Evol. 1989, 167, 137–148. [Google Scholar] [CrossRef]
- Batty, A.L.; Dixon, K.W.; Brundrett, M.; Sivasithamparam, K. Constraints to symbiotic germination of terrestrial orchid seed in a mediterranean bushland. New Phytol. 2001, 152, 511–520. [Google Scholar] [CrossRef]
- Phillips, R.D.; Scaccabarozzi, D.; Retter, B.A.; Hayes, C.; Brown, G.R.; Dixon, K.W.; Peakall, R. Caught in the act: pollination of sexually deceptive trap-flowers by fungus gnats in Pterostylis (Orchidaceae). Ann. Bot. 2013, 113, 629–641. [Google Scholar] [CrossRef]
- Hoffman, N.; Brown, A. Orchids of South-West Australia; Noel Hoffman: Perth, Western Australia, Australia, 2011; pp. 1–512. [Google Scholar]
- Jeanes, J.A. A revision of the Thelymitra pauciflora R. Br. (Orchidaceae) complex in Australia. Muelleria 2004, 19, 19–79. [Google Scholar]
- Rogers, R. Mechanism of pollination in certain Australian Orchids. Trans. Proc. R. Soc. S. Aust. 1913, 37, 48–61. [Google Scholar]
- Bates, R. The genus Microtis R. Br. (Orchidaceae): A taxonomic revision with notes on biology. J. Adel. Bot. Gard. 1984, 7, 45–89. [Google Scholar]













| No | Strategy | Insect Group | Season | Orchid Genus | Average Gruit Set per Flower |
|---|---|---|---|---|---|
| 1A | Visual deception | bees, etc. | winter | Pheladenia Diuris | P. deformis—0%* D. brumalis—2% [23] |
| 1B | Visual deception | bees, beetles, flies, etc. | spring | Caladenia | C. flava—most flowers [44],—25% [18], —18% [83], —32% [77], —48%*; C. latifolia—47%*; C. polychroma—16% [77]; C. melanema R—8% [84]; |
| Diuris | D. corymbosa—most flowers [44]; D. magnifica—11% [83], —6%*; D. maculata—15% [22]; | ||||
| 1C | Visual deception | Flies, bees, etc. | spring - early summer | Thelymitra | T. antennifera—3.2% [66]; T. ixioides—28% [85]; T. macrophylla—4.8%*; T. graminea—17%*; T. fuscolutea (drought stressed)—7%* |
| 2A | Some nectar | bees | autumn | Eriochilus | E. dilatatus—95% pollen deposition [44], —16%* |
| 2B | Some nectar | bees | spring | Pyrorchis | P. nigricans—31%* |
| 3A | Sexual deception | male ant | autumn | Leporella | L. fimbria—18% [86], —17%* |
| 3B | Sexual deception | fungus gnats | winter or spring | Pterostylis | P. recurva—50%, [18]), —50%*; P. sanguinea—44%, [83], —23%*; P. ectypha—39%* |
| 3C | Sexual deception | thynnine wasps | spring | Caladenia | C. arenicola—4% [87], —10% [77], —18% [83], —13%*; C. discoidea—16%*; C. huegelii R—7% [77], —2% (Brundrett unpublished); C. graniticola R—6% [84]; C. williamsiae R—15% [84] |
| Drakaea | D. glyptodon—0 to 59% 9 sites [65]; D. elastica R—5% (Brundrett unpublished) | ||||
| 4A | Self–pollination | none | spring | Thelymitra | T. vulgaris—95%*; T. benthamiana—90%*; T. circumsepta R—96% [85] |
| 4B | Self–pollination (some nectar) | none (usually) | spring – summer | Microtis Disa | M. media—93%*; D. bracteata (drought stressed)—59%* |
1. Pollinator abundance is the key limiting factor to pollination success.
|
2. Pollination is primarily limited by learned avoidance responses by insects in deceptive species.
|
3. Pollinator abundance is strongly limited by seasonal climatic factors.
|
4. Flowering and seedpod development are limited by resources available to the plant.
|
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brundrett, M.C. A Comprehensive Study of Orchid Seed Production Relative to Pollination Traits, Plant Density and Climate in an Urban Reserve in Western Australia. Diversity 2019, 11, 123. https://doi.org/10.3390/d11080123
Brundrett MC. A Comprehensive Study of Orchid Seed Production Relative to Pollination Traits, Plant Density and Climate in an Urban Reserve in Western Australia. Diversity. 2019; 11(8):123. https://doi.org/10.3390/d11080123
Chicago/Turabian StyleBrundrett, Mark C. 2019. "A Comprehensive Study of Orchid Seed Production Relative to Pollination Traits, Plant Density and Climate in an Urban Reserve in Western Australia" Diversity 11, no. 8: 123. https://doi.org/10.3390/d11080123
APA StyleBrundrett, M. C. (2019). A Comprehensive Study of Orchid Seed Production Relative to Pollination Traits, Plant Density and Climate in an Urban Reserve in Western Australia. Diversity, 11(8), 123. https://doi.org/10.3390/d11080123