1. Introduction
Federal recovery plans for imperiled freshwater mussels identify the quantification of demographic characteristics—such as population size, age-class structure, and survival rates—as key to assessing species recovery. Estimation of demographic parameters is vital to understanding species-specific population dynamics and, ultimately, assessing population viability [
1,
2]. In recent years, reintroductions of mussel species into historical habitats where they were extirpated, and augmentations of extant but generally declining populations were conducted to recover imperiled species and to prevent future losses [
3]. These recovery efforts require post-release monitoring of demographic vital rates to assess restoration success and evaluate whether down or delisting criteria have been met. Data from post-release monitoring studies help biologists make informed decisions to adaptively manage populations [
1,
4,
5,
6].
Probability-based, quadrat surveys are a common quantitative sampling approach used to collect demographic data for mussel population assessments. Systematic random sampling is more efficient than simple random sampling and is considered an appropriate design for rare, clustered populations when auxiliary information is not available for stratification [
7,
8,
9]. Fundamentals of this approach involve conducting a complete census typically within 0.25 m
2 sampling units (i.e., quadrats) that have been randomly placed—according to sampling design—within a study site and extrapolating the findings across the site to evaluate parameters, such as diversity, population size and density, growth rates, sex ratios, age-class structures, and evidence of recruitment [
3,
6,
10]. From the length data on live mussels encountered and shells collected during quadrat sampling, catch-curve and shell thin-sectioning analyses can be used to estimate survival rates for demographic models. However, the underlying assumptions (e.g., constant recruitment, mortality, and survival rates across age-classes) of these techniques for survival analyses are rarely met by natural populations [
2,
11]. Alternatively, following uniquely marked individuals or cohorts through time supports improved estimates of survival based on the fates of recaptured individuals. The use of model-based mark-recapture estimators have a long history in wildlife ecology [
12,
13,
14,
15,
16,
17,
18,
19,
20,
21] and are commonly used in studies of many other taxa [
22,
23,
24]. While not traditionally used to assess mussel population dynamics, mark-recapture studies on mussels are increasing in frequency [
25,
26,
27,
28,
29,
30,
31].
In a capture-mark-recapture (CMR) study, repeated sampling of a population is required to separate nondetection from true absence. During the first sampling event, all individuals captured are uniquely marked, recorded, and released back into the population. During subsequent, independent sampling occasions, identities of recaptured individuals are recorded, and all unmarked captures are uniquely marked before release [
20,
21]. After the final sampling event, capture data can be compiled to create an encounter history for each individual. In the simplest form of CMR, population size can be estimated from a single recapture event based on the proportion of marked to unmarked individuals encountered [
9,
12,
13,
17,
32]. Data from marking and recapturing individuals over multiple sampling events can also be used to investigate factors influencing capture and site fidelity probabilities, temporary emigration, and population growth, survival, mortality, and recruitment rates [
17,
18,
21,
26,
27,
32,
33,
34,
35].
Capture-mark-recapture models fall into two broad classes: closed- and open-population models. Generally, closed-population estimators are used to estimate population size when capture-recapture sampling occasions occur over a relatively short period of time (e.g., days to weeks, or time relative to the species’ life-history) to ensure demographic and geographic closure during the study period. Simple single-recapture (e.g., Lincoln–Petersen) and multiple-recapture (e.g., Schnabel) estimators, to more complex multiple-recapture estimators [
12,
13,
20], are available for closed-population sampling designs. Open-population estimators (e.g., Cormack–Jolly–Seber) require multiple recapture events and are frequently used for CMR data collected over longer periods of time (e.g., months to years) to obtain estimates of recruitment, survival, mortality or site fidelity. A common assumption of CMR models is that all animals—marked and unmarked—are equally likely to be caught during any sampling occasion (i.e., the equal catchability assumption). However, the assumptions underlying closed- and open-population estimators are frequently violated in CMR field studies by the inherent variability in capture probabilities due to individual heterogeneity, trap response, time effects, temporal emigration, and combinations of these and other factors [
16,
17,
18,
20,
21,
32,
33]. To cope with capture variability and account for incomplete detection, CMR models have been developed that support parameter estimations that incorporate such factors [
20,
21,
32,
33,
34,
36,
37,
38,
39].
Mussel populations are often sampled without accounting for imperfect detection, with the expectation being that all individuals within a defined sampling unit (e.g., quadrat) are detected. Still, perfect detectability within a quadrat unit is not always obtainable as recruits and young mussels (<10 mm) are inherently difficult to detect, even when excavation and sieving methods are employed. Complete and constant detectability is not realistic in practice, as capture rates can vary temporally, spatially, and by various other—potentially interacting—biotic and abiotic factors (e.g., species, age, sex, size, habitat type, sampling conditions) [
9,
21,
27,
29]. Although mussels are relatively sedentary animals, their ability to move vertically in the substrate can make them temporarily unavailable for detection on the substrate surface [
26,
27]. Much like underwater fish surveys, visual search efficiencies for mussels can be strongly influenced by sampling depths, flows, substrate composition, vegetation, and visibility [
27,
40,
41,
42]. While quadrat-based designs for mussels can investigate variation in detectability on the surface (sampling efficiency at a particular point in time) through excavation of sampling units, they are limited in their ability to capture all sources of variability in detection over time and space; particularly site to site differences (e.g., community assemblages, hydrogeomorphology, habitat heterogeneity). Although CMR can be data intensive depending on project objectives, it offers an approach that allows for the incorporation and investigation of underlying sources of capture biases. Monitoring designs that fail to incorporate differences in detectability can result in biased parameter estimates, leading to false inferences of population status and trends [
26,
27,
29,
34,
43]. Hence, the capability to assess species recovery efforts with confidence and to make informed management decisions relies on the ability of monitoring programs to accurately quantify population demographics with precision.
We chose a recently reintroduced mussel population in the Upper Clinch River, Virginia, that was in need of follow-up monitoring to examine the use of model-based CMR and probability-based quadrat sampling designs for estimating mussel population parameters. Once common throughout the Upper Clinch River, the federally endangered
Epioblasma capsaeformis has experienced significant declines over the past half century due to various anthropogenic impacts on habitat and water quality. By the mid-1980s, the native Upper Clinch River population in Virginia had declined to virtually undetectable levels. Improvements to habitat and water quality over the last 20 years have allowed mussel and fish populations to recover in portions of the river and, in 2002, the Virginia Department of Game and Inland Fisheries (VDGIF) designated a 19.3 km reach of the Upper Clinch River as suitable for mussel population recovery efforts [
3,
44,
45,
46,
47]. In a multiagency collaboration with VDGIF’s Aquatic Wildlife Conservation Center (AWCC) and the U.S. Fish and Wildlife Service (USFWS), Virginia Tech’s Freshwater Mollusk Conservation Center (FMCC) has been working to restore
E. capsaeformis across this reach since 2005. Prior to the start of recovery efforts, the last
E. capsaeformis within this reach were found in 1985 [
48].
One of the population restoration sites within the reach, Cleveland Islands, has received extensive
E. capsaeformis reintroduction efforts since 2006. By 2011, over 4000 individuals were reintroduced to the site using translocation and captive propagation methods. To evaluate the success of these reintroduction efforts at Cleveland Islands, follow-up monitoring was initiated in 2011 [
3,
28]. This population restoration site presented an ecological opportunity to increase knowledge of species-specific demographic rates and to compare the relative performance of CMR to conventional quadrat sampling designs because all reintroduced
E. capsaeformis were uniquely marked. Using systematic quadrat and CMR sampling methods, we collected data on
E. capsaeformis and two nonlisted, naturally occurring species—
Actinonaias pectorosa and
Medionidus conradicus—at Cleveland Islands over two years (2011–2012) to estimate and compare abundance and precision, and relative sampling efficiencies, between sampling designs. In addition, we used CMR models to investigate factors influencing capture probabilities and to assess reintroduced
E. capsaeformis survival rates.
4. Discussion
Our study has shown that E. capsaeformis population restoration efforts since 2006 were successful in the Upper Clinch River at Cleveland Islands, Virginia, and that CMR offers more additional applications for the inference of demographic parameters for mussels relative to quadrat data. Evidence of recruitment was documented by both sampling methods, indicating that natural reproduction is occurring for this restored E. capsaeformis population. Although other studies have used mark-recapture methods to assess mussel populations and investigate factors influencing detectability, this is the first study to directly compare the reliability and precision of CMR population parameter estimates relative to those obtained through quadrat sampling for freshwater mussels. Our comparisons showed that population parameter estimates were, generally, similar between CMR and systematic quadrat sampling approaches, but that there was considerable variability in the level of precision obtained around parameter estimates between sampling designs, study years, and among species. Although CMR was approximately four times more time-intensive (person-hours effort) than quadrat sampling, we processed nearly five times the number of mussels per person-hour effort and encountered over four to eleven times the number of unique LPSA and TA E. capsaeformis individuals, respectively, than we did from systematic quadrat sampling.
Estimating species-specific demographic vital rates is essential to assessing reintroduction success, evaluating whether delisting criteria have been met, and developing effective management plans [
1,
3,
26,
27]. The common methodology for collecting mussel demographic data is through probability-based quadrat sampling designs, which have been used in the Clinch River since the mid-1970s [
46,
66,
67]. Systematic quadrat sampling is a probability-based survey method for assessing rare or clustered populations, is simple to execute in the field, and offers effective spatial coverage [
7,
10,
43,
50]. In addition, with probability-based sampling, the probability that a species is present at a specified mean density even if it were not detected can be estimated [
10,
68,
69].
Because not all mussels are available at the substrate surface at any point in time (temporary vertical emigration) during which a quadrat survey is being conducted, population parameter estimates may be biased if excavation is not executed [
27,
41,
50,
70]. Even when excavation is applied to minimize biases associated with temporary emigration, it is unknown whether excavation disrupts substrate composition and stability, causes increased mortality, disrupts feeding and reproduction, or causes significant displacement of individuals (i.e., permanent emigration) [
41]. In addition to possible biological disturbances, excavation can be resource intensive. Obtaining reliable and comparable estimates of density using quadrat methods requires some level of excavation effort to account for sources of variability in detection on the substrate surface [
41,
50]. Conversely, substrate excavation is not necessary for CMR to obtain unbiased population estimates as models can account for incomplete detection at the substrate surface over time. Excavating or not, quadrat surveys are often problematic to implement in deep-water and high-velocity habitats [
27] and often are inefficient at detecting the presence of rare species [
41,
69]. Although useful for detecting population trends within a site, quadrat sampling provides only broad estimates of diversity, growth rates, age-class structures, and periodic survival and recruitment rates—particularly when target species are at low densities (
Table 6).
As it is becoming increasingly important to understand and monitor species-specific population dynamics for conservation management, the incorporation of CMR into mussel monitoring studies has steadily been expanding [
25,
26,
27,
29,
30,
31,
71]. Similar to quadrat surveys, CMR is useful for estimating and detecting population trends, but in addition, it can: (1) offer improved precision in population parameter estimates; (2) provide reliable estimates of vital rates (i.e., survival, mortality, recruitment); (3) investigate factors influencing vital rates and detectability; and (4) be used to validate and improve species-specific demographic models—particularly for species occurring at lower densities [
10,
17,
18,
26,
32]. Improved estimates of population parameters are partly a result of high numbers of captures and recaptures of individuals (i.e., increasing sample size). Additionally, determining what factors are important predictors of capture provides biologists with guidelines for species-specific encounter rates through time (e.g., in relation to temperature, discharge, reproductive condition), and thus informs more efficient monitoring plans [
26,
27].
The probability that a mussel is captured at the substrate surface is a function of its availability for detection and its detectability by a surveyor. Mussels generally exhibit seasonal (time) and species-specific reproductive behavior patterns of vertical migration in substrate that are influenced by environmental variables [
70,
72,
73]. Particularly in the absence of excavating and sieving substrates, these patterns can strongly affect the probability of detection during a survey. Several CMR mussel studies have examined (and accounted for) abiotic and biotic factors influencing vertical migration patterns and capture probabilities. These studies found that vertical migration patterns varied by species and season, and determined that capture probabilities were influenced by reproductive behavior, shell length, water temperature, and habitat type [
26,
27]. Similarly, a separate study found that individual recapture probabilities were influenced by shell length, which varied by time and species [
29]. In addition, larger and older individuals tend to be more epibenthic than juveniles and smaller individuals, even during warmer months, suggesting that age and size influence their availability for detection at the substrate surface [
70,
74].
Given that a mussel is available for detection, the likelihood it is encountered can be influenced by factors such as species-specific reproductive behaviors (e.g., spawning, displaying mantle lures, lying on top of the substrate), shell length, aperture size and appearance, habitat type, survey conditions (e.g., turbidity, water depth), and can vary among surveyors [
26,
27,
29]. For example,
Pleurobema collina CMR surveys in the upper James River basin, Virginia, have observed discharge effects on capture rates as a result of reduced visibility associated with turbid sampling conditions (B. Watson, VDGIF, unpublished data). Differences in ability to detect individuals among surveyors can be attributed to years of experience, familiarity with target species or the study area, visual acuity, dedication, and mental or physical fatigue [
26,
27,
29]. Although surveyor ability to detect adults at the substrate surface is often positively associated with increasing mussel shell length, CMR studies have demonstrated that this relationship can vary with substrate size and habitat type [
26,
27,
31]. To make things more complex, many of these factors influencing availability and detectability can have interacting effects and vary by species.
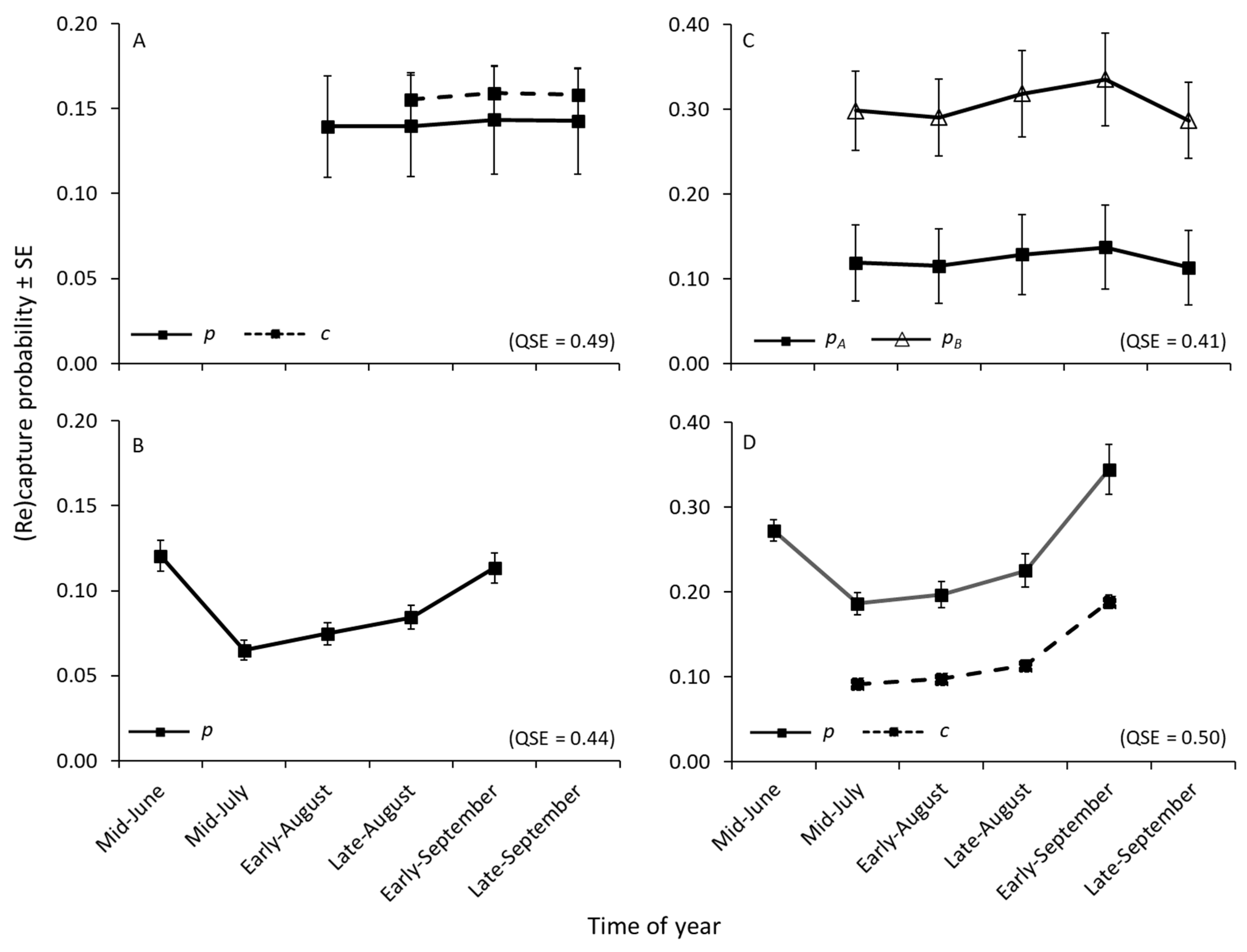
Our results were in agreement with those of previous CMR studies and indicated that capture probabilities varied among species, by time, and were associated with shell length. Although our mean capture probability estimates for
E. capsaeformis (2–6%) were lower than those reported for other species during the warmer months by Villella et al. [
26] (7–19%), Meador et al. [
27] (8–20%), and Watson et al. [
75] (10–30%), our estimated capture probabilities for
A. pectorosa (10–34%) and
M. conradicus (6–16%) were similar. The lower capture rates of
E. capsaeformis may reflect the difficulty of detecting a species existing at much lower densities (<0.4/m
2) than our other two study species, or be related to species-specific behavior or smaller shell size.
Actinonaias pectorosa had the highest capture probabilities, presumably because individuals were comparatively larger in length and aperture size relative to other target species. Interestingly, intra-annual temporal trends in capture were similar across all three study species. Capture probabilities were higher in mid-June, and then declined by mid-July before steadily increasing through late September. Because all three species are long-term brooders—spawning in late summer and autumn, gravid through winter, and releasing glochidia the following late spring to early summer—capture probabilities may have reflected vertical migration patterns associated with reproductive behavior. Because species-specific mean capture probabilities were different between years, but intra-annual trends were similar among species, inter-annual variability in environmental conditions (e.g., temperature, stream discharge) additionally may have played a role.
Obtaining unbiased and precise estimates of specific-specific vital rates and identifying factors influencing mussel survival are important for developing effective conservation plans. Previous CMR studies have reported that annual mussel survival can be influenced by several factors, including age, shell length, habitat type, stream discharge, and invasive unionid densities [
25,
26,
27,
29,
71]. Over a four-year CMR study, Villella et al. [
26] estimated high annual apparent survival rates (>90%) for adult
Elliptio complanata,
E. fisheriana, and
Lampsilis cariosa, and found that survival was time- and size-dependent. Using a Passive Integrated Transponder-tag CMR methodology, Hua et al. [
30] documented high monthly survival rates (>98%) for Cumberlandian combshell (
Epioblasma brevidens) in the Powell River, Tennessee, over a two-year period. Additionally, during a one-year Robust Design CMR study, Meador et al. [
27] reported that variations in survival differed among habitat types and were positively associated with shell length. Similar to these other CMR study findings of high adult annual survival rates (>90%), our results indicated that
E. capsaeformis exhibited high annual apparent survival probabilities (>96%). By understanding the factors affecting survival, reintroduction plan details (e.g., release site habitat characteristics; age, size, sex ratios of released individuals) can be identified and recommended to optimize survival of released individuals, and ultimately, long-term restoration success.
In addition to providing important insights into the ecological relationships affecting vital rates and capture probabilities, CMR can provide invaluable data on age and growth for improving predicted length-at-age growth models, assessing age-class distributions, evaluating spatial and temporal variations in growth, predicting chance of species recovery, and estimating species risk of extinction [
26,
71,
76,
77]. Furthermore, mark-recapture studies can be used to validate previous conclusions on vital rates estimated through other approaches to assessing population parameters, such as quantitative quadrats sampling, length-at-age catch-curves or shell thin-sectioning age and growth analyses. By following unique individuals through time, we were able to estimate survival rates based on fates of individuals captured. Although original aging of uniquely marked TAs was estimated using predicted length-at-age growth curves, our study was able to assess post-release survival by integrating 2011–2012 capture histories with 2006–2011 reintroduction data (i.e., known time since release). Similarly, tagged LPSAs were of known age at release and provided concrete age-specific data for estimating annual survival rates. The results of our study were in agreement with previous predictions—high annual survival for subadult and adult age-classes—estimated using shell thin-sectioning analyses and modeling of length-at-age data of mussels collected from systematic quadrat surveys in the Clinch River, Tennessee [
2].
When combined, length-at-release reintroduction data and measurements taken during the 2011–2012 study period provided a considerable amount of data on absolute growth for
E. capsaeformis. However, a von Bertalanffy growth curve [
78] could not accurately be fitted to our presently available growth data for several reasons: (1) the ages of TAs were extrapolated estimates based on predicted length-at-age growth curves in Jones and Neves [
6], (2) LPSA growth data only represented younger age-classes (≤3 years-old), and (3) shell thin-sectioning was not conducted. Even though LPSA data provided known—as opposed to estimated— length-at-age, using the available data for 1–3-year-olds likely would have resulted in biased estimates of growth parameters. Through shell thin-sectioning and future monitoring at our study site, a complete age and growth data set for reintroduced
E. capsaeformis can be compiled and a predicted length-at-age growth curve can be computed to compare to that of Jones and Neves [
6]. Consequently, further data from marked individuals can be used to assess the accuracy and precision of shell thin-sectioning, test the assumptions of shell annuli formation for
E. capsaeformis, and examine disturbance ring deposition for reintroduced individuals [
79,
80].
Also of concern is whether sampling and monitoring efforts can cause declines in abundance and density due to disturbance. Mussels are known to lay down disturbance rings, which represent brief cessations of growth due to factors such as handling or natural disturbances, but it is unknown how much disturbance—through the excavation of substrate or removal of mussels from substrate for processing—influences mortality rates or increases displacement of individuals [
80]. In this study, CMR estimators indicated a significant decline in
A. pectorosa abundance between 2011 and 2012; a trend not detected by the systematic quadrat survey. It is likely that displacement from the site—rather than natural or induced mortality—was responsible for this estimated decline in abundance. Other multiyear studies suggested low to no mortality from presumed similar levels of handling stress, nor did they reveal related declines in abundance [
26,
80,
81]. Even though all mussels were returned to the area where they were found in the substrate, the average size of
A. pectorosa individuals was larger than our other study species, and these mussels may have had a more difficult time reburrowing into the substrate after handling. This, in combination with surveyors moving about on the streambed and high flow events after surveying, may have displaced some
A. pectorosa individuals downstream out of the survey area, resulting in the decline in abundance noted from the effective sampling area. Further examination is needed to assess whether this decline in abundance, estimated through CMR, can be attributed to mortality (natural or handling associated) or due to displacement outside the study area.
Despite the large number of mussel population restoration projects that have been conducted over the last century [
73], few have determined the long-term success of these efforts [
82]. Detecting population trends, estimating species-specific vital rates, identifying factors influencing capture and survival, and long-term monitoring of population dynamics are essential to developing effective conservation plans and to determine long-term success of reintroduction efforts [
5]. By performing and reporting post-restoration population monitoring, projects can provide insight into the relative success of method-specific restoration efforts and population viability. Both systematic quadrat and CMR sampling techniques have useful applications in population monitoring—and towards assessing population viability—but are dependent on project objectives.
Our results indicated that CMR has advantages over quadrat sampling for quantitatively monitoring mussel populations. Incorporating appropriate CMR methodologies into monitoring studies can provide greater insight into species-specific population dynamics through the ability to account for imperfect detection, increase sample size, and ultimately produce more reliable and precise population parameter estimates. However, CMR sampling can be considerably more resource-intensive than other probability-based designs, depending on the scope of the project and CMR study design. While our study was sampling-intensive in order to thoroughly compare CMR to systematic quadrat sampling population size estimators for mussels, there are many different—and less resource-intensive—sampling and data collection approaches, data analysis strategies, and statistical models available for biologists to consider when designing a CMR study. Developing an effective, efficient, and feasible monitoring program that can achieve desired goals requires identifying clearly defined and quantifiable objectives to guide informed survey design decisions [
10,
43,
69].
In addition to taking project objectives and availability of resources into consideration, the selection of an appropriate sampling design for monitoring and analysis should consider other factors such as study site characteristics and species’ expected densities and distributions [
8]. For example, biologists interested in employing CMR sampling across large study areas (e.g., >2000 m
2), or in difficult-to-sample habitats, could consider stratifying reaches (e.g., habitat type, species’ densities, distributions) and allocating efforts to randomly chosen sampling units (e.g., line-transects) within randomly selected reaches because it would be less resource intensive and could still provide improved reliability and precision in population estimates relative to quadrat sampling [
27]. Alternatively, if a study area is relatively small (e.g., <500 m
2), a CMR design surveying the entire substrate surface area could easily be implemented in a cost-effective manner. Finally, a Robust Design (integrated closed- and open-population parameter estimators) CMR study is recommended for long-term (≥3 years) monitoring as it allows for estimation of abundance and true survival with improved precision, (temporary) emigration, and recruitment rates—which is crucial to evaluating population viability [
2,
21,
26,
27,
31,
33,
83,
84]. Overall, CMR approaches are appropriate and feasible for monitoring populations within or across a few study sites. However, further research is needed on the applicability and feasibility of CMR sampling to watershed-wide (i.e., spatially extensive) assessments compared to other alternative, less intensive, designs.
We recommend that monitoring projects use quadrat sampling approaches when the objective is to simply estimate and detect trends in population size for established, or restored, species of moderate to high densities (>0.2/m
2). Capture-mark-recapture should be used or incorporated into existing monitoring programs when objectives include assessing restored populations of reintroduced or augmented species at low to moderate densities, obtaining reliable and precise estimates of population parameters (e.g., survival, recruitment), or producing unbiased estimates of abundance with precision for species occurring at low to moderate densities (≤0.2/m
2). Future mussel restoration efforts should uniquely mark released individuals and incorporate a CMR monitoring and analysis component to the project to improve our understanding of species-specific demographic characteristics as well as to assess likelihood of population restoration success (
Table 6).
Even though there was not enough data on recruited
E. capsaeformis for CMR models, our observations confirmed that natural recruitment is occurring—a measure of short-term reintroduction success. We believe long-term reintroduction success could not be assessed in our 2011–2012 surveys due to the time frame of the project, i.e., monitoring immediately followed reintroductions and consequential natural recruitment can take several years before it is self-sustaining and evident. Continued long-term monitoring efforts will be essential to evaluating
E. capsaeformis recruitment rates at the restoration site and determine if they are reaching self-sustaining levels or are in need of additional augmentations. Results from this follow-up monitoring study, and future monitoring efforts, will improve our understanding of
E. capsaeformis vital rates, and provide data on effective population sizes and demographic structures required to make informed decisions for future recovery projects. In accordance with the recovery plan for
E. capsaeformis [
1], we recommend biennial CMR monitoring efforts at Cleveland Islands, Virginia to reveal whether these recovery efforts were ultimately successful at restoring a long-term viable deme of
E. capsaeformis to the Upper Clinch River—information essential for effective future management and recovery plans.

 and
and 

{kind=link}
{kind=link}
{kind=link}


