Endemic Infection of Batrachochytrium dendrobatidis in Costa Rica: Implications for Amphibian Conservation at Regional and Species Level

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Species Assessment
2.2. Field Dataset
2.3. Combined Dataset
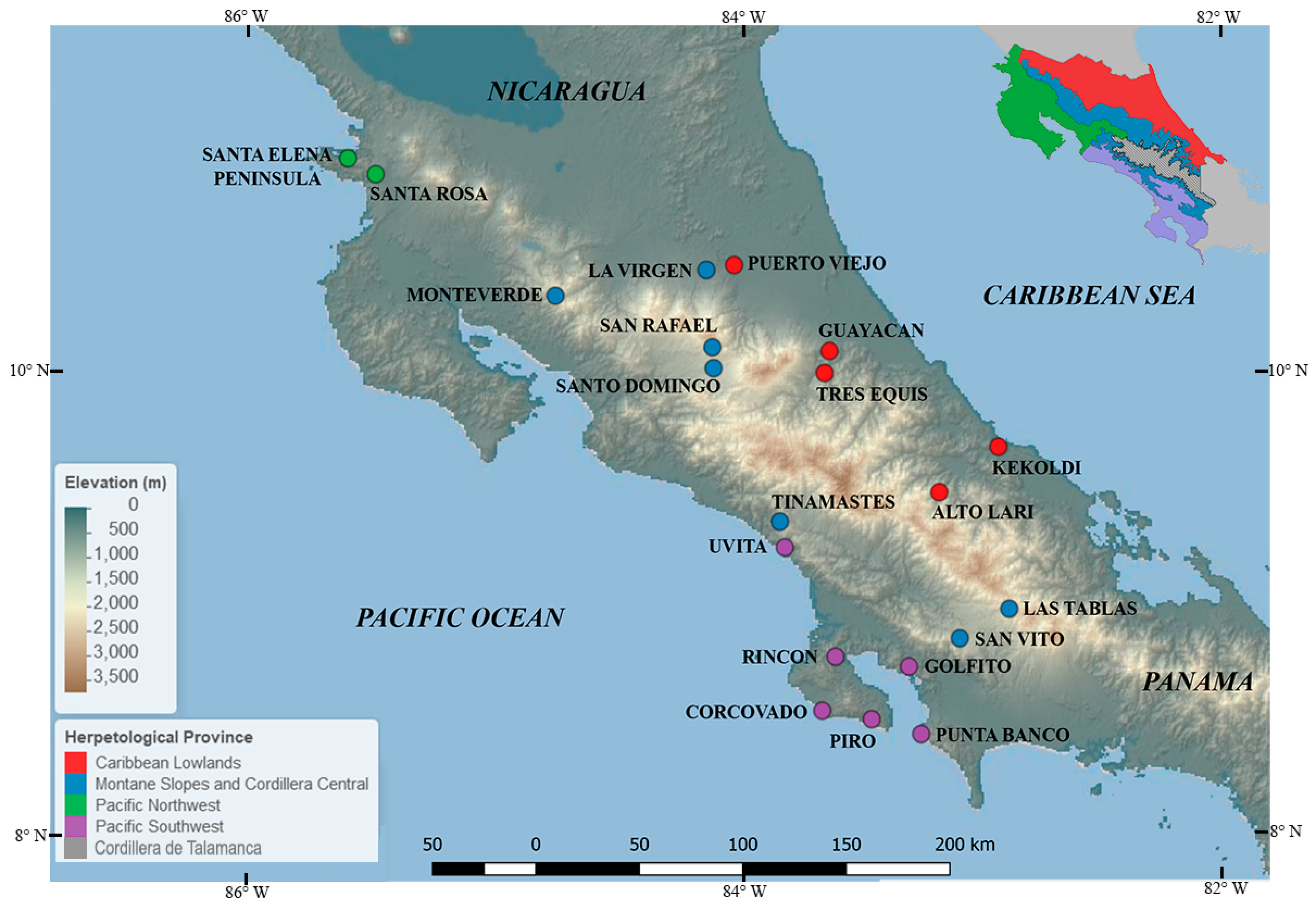
2.3.1. Herpetological Provinces
2.3.2. Altitudinal Belt
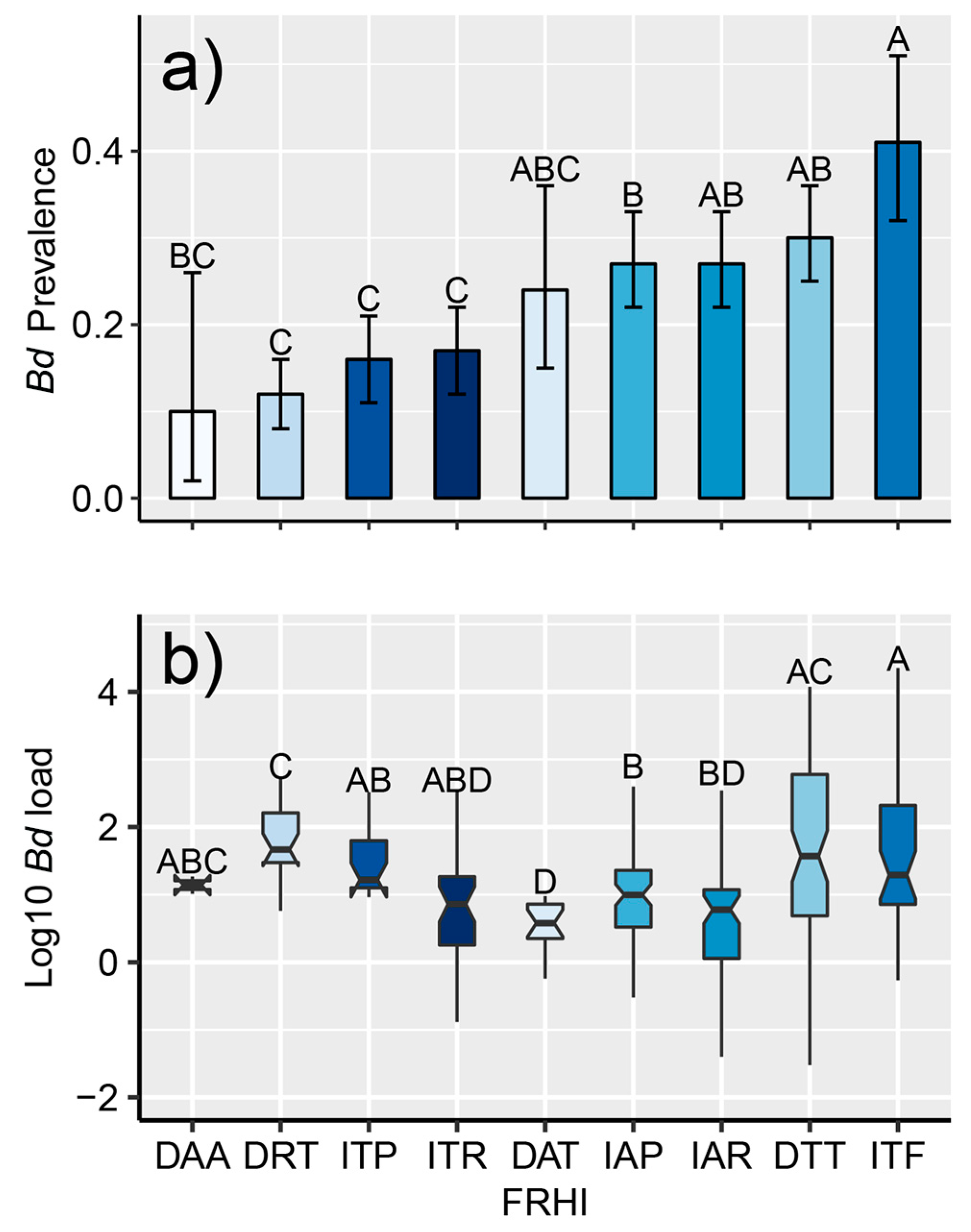
2.3.3. Foraging-Reproduction Habitat Index
2.4. Statistical Analysis
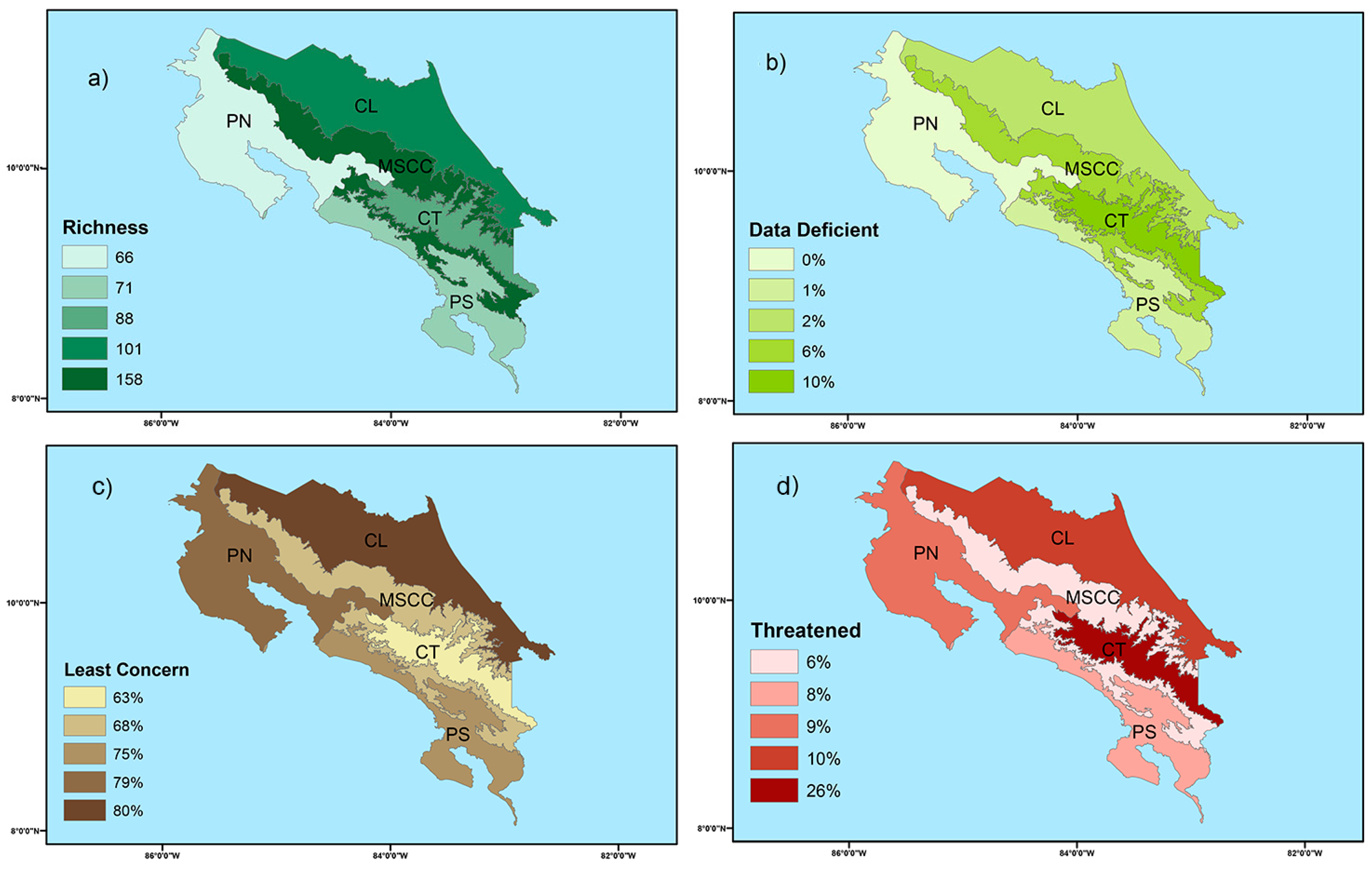
3. Results
3.1. Species Assessment
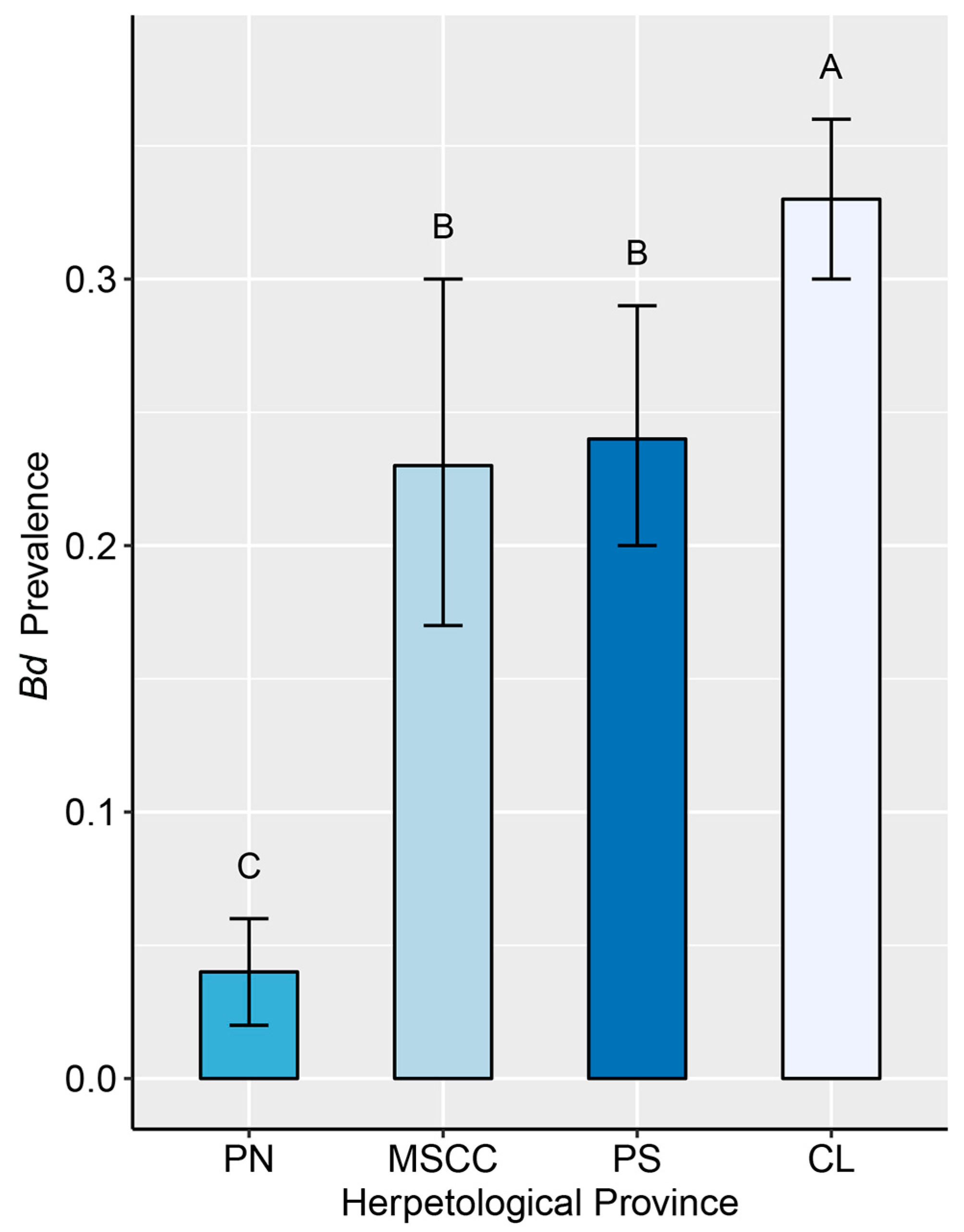
3.2. Endemic Dynamics
4. Discussion
4.1. Species Assessment
4.2. Post-Decline Dynamics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Novacek, M.J.; Cleland, E.E. The current biodiversity extinction event: Scenarios for mitigation and recovery. Proc. Natl. Acad. Sci. USA 2001, 98, 5466–5470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnosky, A.D.; Matzke, N.; Tomiya, S.; Wogan, G.O.U.; Swartz, B.; Quental, T.B.; Marshall, C.; McGuire, J.L.; Lindsey, E.L.; Maguire, K.C.; et al. Has the Earth’s sixth mass extinction already arrived? Nature 2011, 471, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Wake, D.B.; Vredenburg, V.T. Are we in the midst of the sixth mass extinction? A view from the world of amphibians. Proc. Natl. Acad. Sci. USA 2008, 105, 11466–11473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monastersky, R. Life—A status report. Nature 2014, 516, 158–161. [Google Scholar] [CrossRef] [PubMed]
- Stuart, S.N.; Chanson, J.; Cox, N.A.; Young, B.E.; Rodrigues, A.S.L.; Fischman, D.; Waller, R. Status and trends of amphibian declines and extinctions worldwide. Science 2004, 306, 1783–1786. [Google Scholar] [CrossRef]
- Catenazzi, A. State of the World’s Amphibians. Annu. Rev. Environ. Resour. 2015, 40, 91–119. [Google Scholar] [CrossRef]
- Daszak, P.; Berger, L.; Cunningham, A.A.; Hyatt, A.D.; Green, D.E.; Speare, R. Emerging infectious diseases and amphibian population declines. Emerg. Infect. Dis. 1999, 5, 735–748. [Google Scholar] [CrossRef] [PubMed]
- Collins, J.P. Amphibian decline and extinction: What we know and what we need to learn. Dis. Aquat. Organ. 2010, 92, 93–99. [Google Scholar] [CrossRef] [PubMed]
- La Marca, E.; Lips, K.R.; Lotters, S.; Puschendorf, R.; Ibanez, R.; Rueda-Almonacid, J.V.; Schulte, R.; Marty, C.; Castro, F.; Manzanilla-Puppo, J.; et al. Catastrophic population declines and extinctions in Neotropical harlequin frogs (Bufonidae: Atelopus). Biotropica 2005, 37, 190–201. [Google Scholar] [CrossRef]
- Scheele, B.C.; Pasmans, F.; Skerratt, L.F.; Berger, L.; Martel, A.; Beukema, W.; Acevedo, A.A.; Burrowes, P.A.; Carvalho, T.; Catenazzi, A.; et al. Amphibian fungal panzootic causes catastrophic and ongoing loss of biodiversity. Science 2019, 363, 1459–1463. [Google Scholar] [CrossRef]
- Gerber, B.D.; Converse, S.J.; Muths, E.; Crockett, H.J.; Mosher, B.A.; Bailey, L.L. Identifying species conservation strategies to reduce disease-associated declines. Conserv. Lett. 2018, 11, e12393. [Google Scholar] [CrossRef]
- Meredith, H.; Van Buren, C.; Antwis, R.E. Making amphibian conservation more effective. Conserv. Evid. 2016, 13, 1–6. [Google Scholar]
- Savage, J.M. The Amphibians and Reptiles of Costa Rica: A Herpetofauna between Two Continents, between Two Deas; University of Chicago Press: Chicago, IL, USA, 2002. [Google Scholar]
- Frost, D.R. Amphibian Species of the World: An Online Reference. Version 6.0. Available online: http://research.amnh.org/vz/herpetology/amphibia/ (accessed on 27 June 2019).
- Bagley, J.C.; Johnson, J.B. Phylogeography and biogeography of the lower Central American Neotropics: Diversification between two continents and between two seas. Biol. Rev. 2014, 89, 767–790. [Google Scholar] [CrossRef] [PubMed]
- Bolaños, F. Situación de los anfibios de Costa Rica. Biocenosis 2009, 22, 95–108. [Google Scholar]
- Pounds, J.A.; Crump, M.L. Amphibian declines and climate disturbance: The case of the golden toad and the harlequin frog. Conserv. Biol. 1994, 8, 72–85. [Google Scholar] [CrossRef]
- González-Maya, J.F.; Belant, J.L.; Wyatt, S.A.; Schipper, J.; Cardenal, J.; Corrales, D.; Cruz-Lizano, I.; Hoepker, A.; Escobedo-Galván, A.H.; Castañeda, F.; et al. Renewing hope: The rediscovery of Atelopus varius in Costa Rica. Amphib. Reptil. 2013, 34, 573–578. [Google Scholar] [CrossRef]
- Chaves, G.; Zumbado-Ulate, H.; García-Rodríguez, A.; Gómez, E.; Vredenburg, V.T.; Ryan, M.J. Rediscovery of the critically endangered streamside frog, Craugastor taurus (Craugastoridae), in Costa Rica. Trop. Conserv. Sci. 2014, 7, 628–638. [Google Scholar] [CrossRef]
- Abarca, J.; Chaves, G.; García-Rodríguez, A.; Vargas, R. Reconsidering extinction: Rediscovery of Incilius holdridgei (Anura: Bufonidae) in Costa Rica after 25 years. Herpetol. Rev. 2010, 41, 150. [Google Scholar]
- Hero, J.-M.; Williams, S.E.; Magnusson, W.E. Ecological traits of declining amphibians in upland areas of eastern Australia. J. Zool. 2005, 267, 221–232. [Google Scholar] [CrossRef]
- Mendelson, J.R.; Whitfield, S.M.; Sredl, M.J. A recovery engine strategy for amphibian conservation in the context of disease. Biol. Conserv. 2019, 236, 188–191. [Google Scholar] [CrossRef]
- Longcore, J.E.; Pessier, A.P.; Nichols, D.K. Batrachochytrium dendrobatidis gen. et sp. nov., a chytrid pathogenic to amphibians. Mycologia 1999, 91, 219–227. [Google Scholar] [CrossRef]
- Berger, L.; Speare, R.; Daszak, P.; Green, D.E.; Cunningham, A.A.; Goggin, C.L.; Slocombe, R.; Ragan, M.A.; Hyatt, A.D.; McDonald, K.R.; et al. Chytridiomycosis causes amphibian mortality associated with population declines in the rain forests of Australia and Central America. Proc. Natl. Acad. Sci. USA 1998, 95, 9031–9036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lips, K.R.; Green, D.E.; Papendick, R. Chytridiomycosis in wild frogs from southern Costa Rica. J. Herpetol. 2003, 37, 215–218. [Google Scholar] [CrossRef]
- Puschendorf, R.; Bolaños, F.; Chaves, G. The amphibian chytrid fungus along an altitudinal transect before the first reported declines in Costa Rica. Biol. Conserv. 2006, 132, 136–142. [Google Scholar] [CrossRef]
- Piotrowski, J.S.; Annis, S.L.; Longcore, J.E. Physiology of Batrachochytrium dendrobatidis, a chytrid pathogen of amphibians. Mycologia 2004, 96, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Pounds, A.J.; Bustamante, M.R.; Coloma, L.A.; Consuegra, J.A.; Fogden, M.P.L.; Foster, P.N.; La Marca, E.; Masters, K.L.; Merino-Viteri, A.; Puschendorf, R.; et al. Widespread amphibian extinctions from epidemic disease driven by global warming. Nature 2006, 439, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Zumbado-Ulate, H.; Bolaños, F.; Willink, B.; Soley-Guardia, F. Population status and natural history notes on the critically endangered stream dwelling frog Craugastor ranoides (Craugastoridae) in a Costa Rican tropical dry forest. Herpetol. Conserv. Biol. 2011, 6, 455–464. [Google Scholar]
- Puschendorf, R.; Carnaval, A.C.; VanDerWal, J.; Zumbado-Ulate, H.; Chaves, G.; Bolaños, F.; Alford, R.A. Distribution models for the amphibian chytrid Batrachochytrium dendrobatidis in Costa Rica: Proposing climatic refuges as a conservation tool. Divers. Distrib. 2009, 15, 401–408. [Google Scholar] [CrossRef]
- Whitfield, S.M.; Bell, K.E.; Philippi, T.; Sasa, M.; Bolaños, F.; Chaves, G.; Savage, J.M.; Donnelly, M.A. Amphibian and reptile declines over 35 years at La Selva, Costa Rica. Proc. Natl. Acad. Sci. USA 2007, 104, 8352–8356. [Google Scholar] [CrossRef] [Green Version]
- Christie, M.R.; Searle, C.L. Evolutionary rescue in a host–pathogen system results in coexistence not clearance. Evol. Appl. 2018, 11, 681–693. [Google Scholar] [CrossRef]
- Retallick, R.W.R.; Miera, V. Strain differences in the amphibian chytrid Batrachochytrium dendrobatidis and non-permanent, sub-lethal effects of infection. Dis. Aquat. Organ. 2007, 75, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Briggs, C.J.; Vredenburg, V.T.; Knapp, R.A.; Rachowicz, L.J. Investigating the population-level effects of chytridiomycosis: An emerging infectious disease of amphibians. Ecology 2005, 86, 3149–3159. [Google Scholar] [CrossRef]
- Rachowicz, L.J.; Knapp, R.A.; Morgan, J.A.T.; Stice, M.J.; Vredenburg, V.T.; Parker, J.M.; Briggs, C.J. Emerging infectious disease as a proximate cause of amphibian mass mortality. Ecology 2006, 87, 1671–1683. [Google Scholar] [CrossRef]
- Retallick, R.W.R.; McCallum, H.; Speare, R. Endemic infection of the amphibian chytrid fungus in a frog community post-decline. PLoS Biol. 2004, 2, e351. [Google Scholar] [CrossRef] [PubMed]
- Searle, C.L.; Biga, L.M.; Spatafora, J.W.; Blaustein, A.R. A dilution effect in the emerging amphibian pathogen Batrachochytrium dendrobatidis. Proc. Natl. Acad. Sci. USA 2011, 108, 16322–16326. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, V.A.; Moore, A.T.; Young, G.R.; Komar, N.; Reisen, W.K.; Brown, C.R. An enzootic vector-borne virus is amplified at epizootic levels by an invasive avian host. Proc. R. Soc. B Biol. Sci. 2011, 278, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Briggs, C.J.; Knapp, R.A.; Vredenburg, V.T. Enzootic and epizootic dynamics of the chytrid fungal pathogen of amphibians. Proc. Natl. Acad. Sci. USA 2010, 107, 9695–9700. [Google Scholar] [CrossRef] [Green Version]
- Miller, C.A.; Tasse Taboue, G.C.; Ekane, M.M.P.; Robak, M.; Sesink Clee, P.R.; Richards-Zawacki, C.; Fokam, E.B.; Fuashi, N.A.; Anthony, N.M. Distribution modeling and lineage diversity of the chytrid fungus Batrachochytrium dendrobatidis (Bd) in a central African amphibian hotspot. PLoS ONE 2018, 13, e0199288. [Google Scholar] [CrossRef]
- Hitchman, S.M.; Mather, M.E.; Smith, J.M.; Fencl, J.S. Identifying keystone habitats with a mosaic approach can improve biodiversity conservation in disturbed ecosystems. Glob. Chang. Biol. 2018, 24, 308–321. [Google Scholar] [CrossRef]
- Vredenburg, V.T.; Knapp, R.A.; Tunstall, T.S.; Briggs, C.J. Dynamics of an emerging disease drive large-scale amphibian population extinctions. Proc. Natl. Acad. Sci. USA 2010, 107, 9689–9694. [Google Scholar] [CrossRef] [Green Version]
- Brem, F.; Lips, K. Batrachochytrium dendrobatidis infection patterns among Panamanian amphibian species, habitats and elevations during epizootic and enzootic stages. Dis. Aquat. Organ. 2008, 81, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Kriger, K.M.; Hero, J.-M. The chytrid fungus Batrachochytrium dendrobatidis is non-randomly distributed across amphibian breeding habitats. Divers. Distrib. 2007, 13, 781–788. [Google Scholar] [CrossRef]
- Heard, G.W.; Scroggie, M.P.; Ramsey, D.S.L.; Clemann, N.; Hodgson, J.A.; Thomas, C.D. Can habitat management mitigate disease impacts on threatened amphibians? Conserv. Lett. 2018, 11, e12375. [Google Scholar] [CrossRef]
- Scheele, B.C.; Hunter, D.A.; Grogan, L.F.; Berger, L.; Kolby, J.E.; Mcfadden, M.S.; Marantelli, G.; Skerratt, L.F.; Driscoll, D.A. Interventions for reducing extinction risk in chytridiomycosis-threatened amphibians. Conserv. Biol. 2014, 28, 1195–1205. [Google Scholar] [CrossRef] [PubMed]
- Bolanos, F.; Savage, J.M.; Chaves, G. Anfibios y Reptiles de Costa Rica. Listas Zoológicas Actualizadas UCR. Available online: http://museo.biologia.ucr.ac.cr/Listas/Anteriores/HerpCREsp.htm (accessed on 27 June 2019).
- Savage, J.M.; Bolaños, F. A checklist of the amphibians and reptiles of Costa Rica: Additions and nomenclatural revisions. Zootaxa 2009, 2005, 1–23. [Google Scholar] [CrossRef]
- Sasa, M.; Chaves, G.; Porras, L.W. The Costa Rican herpetofauna: Conservation status and future perspectives. In Conservation of Mesoamerican Amphibians and Reptiles; Townsend, J.H., Johnson, J.D., Eds.; Eagle Mountain Press: Salt Lake City, UT, USA, 2010; pp. 510–603. [Google Scholar]
- Chaves, G.; Bolaños, F.; Rodríguez, J.E.; Matamoros, Y. Actualización de las Listas Rojas Nacionales de Costa Rica. Anfibios y Reptiles; Conservation Breeding Specialist Group (SSC/IUCN)/CBSG Mesoamerica): San José, Costa Rica, 2014. [Google Scholar]
- IUCN The IUCN Red List of Threatened Species. Version 2019-1. Available online: https://www.iucnredlist.org/en (accessed on 27 June 2019).
- Wilson, L.D.; McCranie, J.R. The conservation status of the herpetofauna of Honduras. Amphib. Reptile Conserv. 2004, 3, 6–33. [Google Scholar] [PubMed]
- Voyles, J.; Young, S.; Berger, L.; Campbell, C.; Voyles, W.F.; Dinudom, A.; Cook, D.; Webb, R.; Alford, R.A.; Skerratt, L.F.; et al. Pathogenesis of chytridiomycosis, a cause of catastrophic amphibian declines. Science 2009, 326, 582–585. [Google Scholar] [CrossRef]
- Kriger, K.M.; Hines, H.B.; Hyatt, A.D.; Boyle, D.G.; Hero, J.-M. Techniques for detecting chytridiomycosis in wild frogs: Comparing histology with real-time Taqman PCR. Dis. Aquat. Organ. 2006, 71, 141–148. [Google Scholar] [CrossRef]
- Skerratt, L.; Berger, L.; Hines, H.; McDonald, K.; Mendez, D.; Speare, R. Survey protocol for detecting chytridiomycosis in all Australian frog populations. Dis. Aquat. Organ. 2008, 80, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Boyle, D.G.; Boyle, D.B.; Olsen, V.; Morgan, J.A.T.; Hyatt, A.D. Rapid quantitative detection of chytridiomycosis (Batrachochytrium dendrobatidis) in amphibian samples using real-time Taqman PCR assay. Dis. Aquat. Organ. 2004, 60, 141–148. [Google Scholar] [CrossRef]
- Hyatt, A.D.; Boyle, D.G.; Olsen, V.; Boyle, D.B.; Berger, L.; Obendorf, D.; Dalton, A.; Kriger, K.; Hero, M.; Hines, H.; et al. Diagnostic assays and sampling protocols for the detection of Batrachochytrium dendrobatidis. Dis. Aquat. Organ. 2007, 73, 175–192. [Google Scholar] [CrossRef] [PubMed]
- Kriger, K.M.; Hero, J.-M.; Ashton, K.J. Cost efficiency in the detection of chytridiomycosis using PCR assay. Dis. Aquat. Organ. 2006, 71, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Picco, A.M.; Collins, J.P. Fungal and viral pathogen occurrence in Costa Rican amphibians. J. Herpetol. 2007, 41, 746–749. [Google Scholar] [CrossRef]
- Goldberg, C.S.; Hawley, T.J.; Waits, L.P. Local and regional patterns of amphibian chytrid prevalence on the Osa Peninsula, Costa Rica. Herpetol. Rev. 2009, 40, 309–311. [Google Scholar]
- Whitfield, S.M.; Geerdes, E.; Chacon, I.; Ballestero Rodriguez, E.; Jimenez, R.; Donnelly, M.; Kerby, J. Infection and co-infection by the amphibian chytrid fungus and ranavirus in wild Costa Rican frogs. Dis. Aquat. Organ. 2013, 104, 173–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zumbado-Ulate, H.; Bolaños, F.; Gutiérrez-Espeleta, G.; Puschendorf, R. Extremely low prevalence of Batrachochytrium dendrobatidis in frog populations from Neotropical dry forest of Costa Rica supports the existence of a climatic refuge from disease. EcoHealth 2014, 11, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Zumbado-Ulate, H.; García-Rodríguez, A.; Vredenburg, V.T.; Searle, C.L. Infection with Batrachochytrium dendrobatidis is common in tropical lowland habitats: Implications for amphibian conservation. Ecol. Evol. 2019, 9, 4917–4930. [Google Scholar] [CrossRef] [PubMed]
- Saenz, D.; Adams, C.K.; Pierce, J.B.; Laurencio, D. Occurrence of Batrachochytrium dendrobatidis in an anuran community in the southeastern Talamanca region of Costa Rica. Herpetol. Rev. 2009, 40, 311–313. [Google Scholar]
- Abarca, J.G. Quitridiomicosis en Costa Rica: Aislamiento y Descripción de Cepas Circulantes del Patógeno y Análisis de la Microbiota del Hospedero como Posible Factor en la Incidencia de la Enfermedad. Master’s Thesis, Universidad de Costa Rica, Heredia, Costa Rica, 2018. [Google Scholar]
- Lips, K.R.; Diffendorfer, J.; Mendelson, J.R.; Sears, M.W. Riding the wave: Reconciling the roles of disease and climate change in amphibian declines. PLoS Biol. 2008, 6, e72. [Google Scholar] [CrossRef]
- Lips, K.R.; Burrowes, P.A.; Mendelson, J.R.; Parra-Olea, G. Amphibian declines in Latin America: Widespread population declines, extinctions, and impacts. Biotropica 2005, 37, 163–165. [Google Scholar] [CrossRef]
- Whitfield, S.M.; Lips, K.R.; Donnelly, M.A. Amphibian decline and conservation in Central America. Copeia 2016, 104, 351–379. [Google Scholar] [CrossRef]
- Arias, E.; Chaves, G. 140 years after William M. Gabb’s climb to Cerro Pico Blanco. Mesoam. Herpetol. 2014, 1, 176–180. [Google Scholar]
- Holdridge, L.R. Life Zone Ecology; Tropical Science Center: San Jose, Costa Rica, 1967. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.r-project.org/ (accessed on 22 February 2019).
- Burnham, K.P.; Anderson, D.R. Multimodel inference: Understanding AIC and BIC in model selection. Sociol. Methods Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- Ryan, M.; Lips, K.; Giermakowski, J.T. New species of Pristimantis (Anura: Terrarana: Strabomantinae) from lower Central America. J. Herpetol. 2010, 44, 193–200. [Google Scholar] [CrossRef]
- Batista, A.; Hertz, A.; Köhler, G.; Mebert, K.; Vesely, M. Morphological variation and phylogeography of frogs related to Pristimantis caryophyllaceus (Anura: Terrarana: Craugastoridae) in Panama. Salamandra 2014, 50, 155–171. [Google Scholar]
- Kubicki, B.; Salazar, S.; Puschendorf, R. A new species of glassfrog, genus Hyalinobatrachium (Anura: Centrolenidae), from the Caribbean foothills of Costa Rica. Zootaxa 2015, 3920, 069–084. [Google Scholar] [CrossRef]
- Arias, E.; Chaves, G.; Parra-Olea, G. A new species of Craugastor (Anura: Craugastoridae) from the montane rainforest of the Cordillera de Talamanca, Costa Rica. Phyllomedusa J. Herpetol. 2018, 17, 211–232. [Google Scholar] [CrossRef]
- Arias, E.; Chaves, G.; Crawford, A.J.; Parra-Olea, G. A new species of the Craugastor podiciferus species group (Anura: Craugastoridae) from the premontane forest of southwestern Costa Rica. Zootaxa 2016, 4132, 347–363. [Google Scholar] [CrossRef]
- Arias, E.; Hertz, A.; Parra-Olea, G. Taxonomic assessment of Craugastor podiciferus (Anura: Craugastoridae) in lower Central America with the description of two new species. Amphib. Reptile Conserv. 2019, 13, 173–197. [Google Scholar]
- Arias, E.; Chaves, G.; Salazar, S.; Salazar-Zúñiga, J.A.; García-Rodríguez, A. A new species of dink frog, genus Diasporus (Anura: Eleutherodactylidae), from the Caribbean foothills of the Cordillera de Talamanca, Costa Rica. Zootaxa 2019, 4609, 269–288. [Google Scholar] [CrossRef]
- Barquero, M.D.; Araya, M.F. First record of the Greenhouse frog, Eleutherodactylus planirostris (Anura: Eleutherodactylidae), in Costa Rica. Herpetol. Notes 2016, 9, 145–147. [Google Scholar]
- Kubicki, B.; Salazar, S. Discovery of the golden-eyed fringe-limbed treefrog, Ecnomiohyla bailarina (Anura: Hylidae), in the Caribbean foothills of southeastern Costa Rica. Mesoam. Herpetol. 2015, 2, 76–86. [Google Scholar]
- McCranie, J.R. Morphological and systematic comments on the Caribbean lowland population of Smilisca baudinii (Anura: Hylidae: Hylinae) in northeastern Honduras, with the resurrection of Hyla manisorum. Mesoam. Herpetol. 2017, 4, 513–526. [Google Scholar]
- Gray, A.R. Review of the genus Cruziohyla (Anura: Phyllomedusidae), with description of a new species. Zootaxa 2018, 4450, 401–426. [Google Scholar] [CrossRef] [PubMed]
- Kubicki, B.; Arias, E. A beautiful new yellow salamander, genus Bolitoglossa (Caudata: Plethodontidae), from the northeastern slopes of the Cordillera de Talamanca, Costa Rica. Zootaxa 2016, 4184, 329–346. [Google Scholar] [CrossRef] [PubMed]
- Boza-Oviedo, E.; Rovito, S.M.; Chaves, G.; Garcia-Rodriguez, A.; Artavia, L.G.; Bolaños, F.; Wake, D.B. Salamanders from the eastern Cordillera de Talamanca, Costa Rica, with descriptions of five new species (Plethodontidae: Bolitoglossa, Nototriton, and Oedipina) and natural history notes from recent expeditions. Zootaxa 2012, 3309, 36–61. [Google Scholar] [CrossRef]
- Arias, E.; Kubicki, B. A new moss salamander, genus Nototriton (Caudata: Plethodontidae), from the Cordillera de Talamanca, in the Costa Rica-Panama border region. Zootaxa 2018, 4369, 487–500. [Google Scholar] [CrossRef] [PubMed]
- Kubicki, B. A new species of salamander (Caudata: Plethodontidae: Oedipina) from the central Caribbean foothills of Costa Rica. Mesoam. Herpetol. 2016, 3, 819–840. [Google Scholar]
- Kubicki, B.; Arias, E. Vulcan’s Slender Caecilian, Caecilia volcani, in Costa Rica. Mesoam. Herpetol. 2017, 4, 488–492. [Google Scholar]
- Jiménez, R.; Alvarado, G. Craugastor escoces (Anura: Craugastoridae) reappears after 30 years: Rediscovery of an “extinct” Neotropical frog. Amphib. Reptil. 2017, 38, 257–259. [Google Scholar] [CrossRef]
- García-Rodríguez, A.; Chaves, G.; Benavides-Varela, C.; Puschendorf, R. Where are the survivors? Tracking relictual populations of endangered frogs in Costa Rica. Divers. Distrib. 2012, 18, 204–212. [Google Scholar] [CrossRef]
- Lewis, C.H.R.; Richards-Zawacki, C.L.; Ibáñez, R.; Luedtke, J.; Voyles, J.; Houser, P.; Gratwicke, B. Conserving Panamanian harlequin frogs by integrating captive-breeding and research programs. Biol. Conserv. 2019, 236, 180–187. [Google Scholar] [CrossRef]
- Searle, C.L.; Gervasi, S.S.; Hua, J.; Hammond, J.I.; Relyea, R.A.; Olson, D.H.; Blaustein, A.R. Differential host susceptibility to Batrachochytrium dendrobatidis, an emerging amphibian pathogen. Conserv. Biol. 2011, 25, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Young, B.E.; Lips, K.R.; Reaser, J.K.; Ibáñez, R.; Salas, A.W.; Cedeño, J.R.; Coloma, L.A.; Ron, S.; La Marca, E.; Meyer, J.R.; et al. Population declines and priorities for amphibian conservation in Latin America. Conserv. Biol. 2001, 15, 1213–1223. [Google Scholar] [CrossRef]
- Martel, A.; Spitzen-van der Sluijs, A.; Blooi, M.; Bert, W.; Ducatelle, R.; Fisher, M.C.; Woeltjes, A.; Bosman, W.; Chiers, K.; Bossuyt, F.; et al. Batrachochytrium salamandrivorans sp. nov. causes lethal chytridiomycosis in amphibians. Proc. Natl. Acad. Sci. USA 2013, 110, 15325–15329. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, S.M.; Kerby, J.; Gentry, L.R.; Donnelly, M.A. Temporal variation in infection prevalence by the amphibian chytrid fungus in three species of frogs at La Selva, Costa Rica. Biotropica 2012, 44, 779–784. [Google Scholar] [CrossRef]
- Whitfield, S.M.; Zoo Miami, FL, USA. Personal Communication, 2019.
- Whitfield, S.M.; Alvarado, G.; Abarca, J.; Zumbado-Ulate, H.; Zuñiga, I.; Wainwright, M.; Kerby, J. Differential patterns of Batrachochytrium dendrobatidis infection in relict amphibian populations following severe disease-associated declines. Dis. Aquat. Organ. 2017, 126, 33–41. [Google Scholar] [CrossRef]
- Puschendorf, R.; Hoskin, C.J.; Cashins, S.D.; McDonald, K.; Skerratt, L.F.; Vanderwal, J.; Alford, R.A. Environmental refuge from disease-driven amphibian extinction. Conserv. Biol. 2011, 25, 956–964. [Google Scholar] [CrossRef]
- Perez, R.; Richards-Zawacki, C.L.; Krohn, A.R.; Robak, M.; Griffith, E.J.; Ross, H.; Gratwicke, B.; Ibanez, R.; Voyles, J. Field surveys in Western Panama indicate populations of Atelopus varius frogs are persisting in regions where Batrachochytrium dendrobatidis is now enzootic. Amphib. Reptile Conserv. 2014, 8, 30–35. [Google Scholar]
- Woodhams, D.C.; Kilburn, V.L.; Reinert, L.K.; Voyles, J.; Medina, D.; Ibáñez, R.; Hyatt, A.D.; Boyle, D.G.; Pask, J.D.; Green, D.M.; et al. Chytridiomycosis and amphibian population declines continue to spread eastward in Panama. EcoHealth 2008, 5, 268–274. [Google Scholar] [CrossRef]
- Kilburn, V.L.; Ibáñez, R.; Sanjur, O.; Bermingham, E.; Suraci, J.P.; Green, D.M. Ubiquity of the pathogenic chytrid fungus, Batrachochytrium dendrobatidis, in anuran communities in Panamá. EcoHealth 2010, 7, 537–548. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Chaves, V.J.; Madrigal-Elizondo, V.; Chaves, G.; Morera-Chacón, B.; Garcia-Rodriguez, A.; Bolaños, F. View of shifts in the diversity of an amphibian community from a premontane forest of San Ramón, Costa Rica. Rev. Biol. Trop. 2019, 67, 259–273. [Google Scholar]
- Garner, T.W.J.; Walker, S.; Bosch, J.; Leech, S.; Marcus Rowcliffe, J.; Cunningham, A.A.; Fisher, M.C. Life history tradeoffs influence mortality associated with the amphibian pathogen Batrachochytrium dendrobatidis. Oikos 2009, 118, 783–791. [Google Scholar] [CrossRef]
- Kolby, J.E.; Ramirez, S.D.; Berger, L.; Richards-Hrdlicka, K.L.; Jocque, M.; Skerratt, L.F. Terrestrial dispersal and potential environmental transmission of the amphibian chytrid fungus (Batrachochytrium dendrobatidis). PLoS ONE 2015, 10, e0125386. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.J.; Lips, K.R.; Eichholz, M.W. Decline and extirpation of an endangered Panamanian stream frog population (Craugastor punctariolus) due to an outbreak of chytridiomycosis. Biol. Conserv. 2008, 141, 1636–1647. [Google Scholar] [CrossRef]
- Campbell, J.A.; Savage, J.M. Taxonomic reconsideration of Middle American frogs of the Eleutherodactylus rugulosus group (Anura: Leptodactylidae): A reconnaissance of subtle nuances among frogs. Herpetol. Monogr. 2000, 14, 186–292. [Google Scholar] [CrossRef]
- Lips, K.R.; Reeve, J.D.; Witters, L.R. Ecological traits predicting amphibian population declines in Central America. Conserv. Biol. 2003, 17, 1078–1088. [Google Scholar] [CrossRef]
- Köhler, G.; Batista, A.; Carrizo, A.; Hertz, A. Field notes on Craugastor azueroensis (Savage, 1975) (Amphibia: Anura: Craugastoridae). Herpetol Notes 2012, 5, 157. [Google Scholar]
- Hanken, J.; Wake, D.B.; Savage, J.M. A solution to the large black salamander problem (genus Bolitoglossa) in Costa Rica and Panamá. Copeia 2005, 2005, 227–245. [Google Scholar] [CrossRef]
- Sunyer, J.; Wake, D.B.; Obando, L. Distributional data for Bolitoglossa (Amphibia, Caudata, Plethodontidae) from Nicaragua and Costa Rica. Herpetol. Rev. 2012, 43, 564–568. [Google Scholar]
- AlMutairi, B.S.; Grossmann, I.; Small, M.J. Climate model projections for future seasonal rainfall cycle statistics in Northwest Costa Rica. Int. J. Climatol. 2019, 39, 2933–2946. [Google Scholar] [CrossRef]
- Grenyer, R.; Orme, C.D.L.; Jackson, S.F.; Thomas, G.H.; Davies, R.G.; Davies, T.J.; Jones, K.E.; Olson, V.A.; Ridgely, R.S.; Rasmussen, P.C.; et al. Global distribution and conservation of rare and threatened vertebrates. Nature 2006, 444, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Woodhams, D.C.; Bosch, J.; Briggs, C.J.; Cashins, S.; Davis, L.R.; Lauer, A.; Muths, E.; Puschendorf, R.; Schmidt, B.R.; Sheafor, B.; et al. Mitigating amphibian disease: Strategies to maintain wild populations and control chytridiomycosis. Front. Zool. 2011, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Garner, T.W.J.; Schmidt, B.R.; Martel, A.; Pasmans, F.; Muths, E.; Cunningham, A.A.; Weldon, C.; Fisher, M.C.; Bosch, J. Mitigating amphibian chytridiomycosis in nature. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20160207. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Site (Elevation m) and Herpetological Provinces | Sampling Period | % of Infection (n Sampled) | Altitudinal Belt | Reference |
|---|---|---|---|---|
| Monteverde (1400–2000), MSCC | July 2005 | 12.2 (41) | Lower montane | [59] |
| San Vito de Coto Brus (1120–1385), MSCC | 9.3 (43) | Premontane | ||
| Rincón de Osa (0–100), PS | May–June 2006 | 0.1 (91) | Lowland | [60] |
| Piro (0–100), PS | 0.0 (62) | Lowland | ||
| Corcovado (0–100), PS | 0.1 (25) | Lowland | ||
| Kekoldi (0–100), CL | January 2008 | 7.9 (126) | Lowland | [64] |
| La Virgen de Sarapiquí (0–200), CL | January–March 2011 | 21.3 (253) | Lowland | [61] |
| Santa Elena Peninsula (0–200), PN | January–March 2007–2008 | 0.0 (310) | Lowland | [62] |
| Santa Rosa (0–200), PN | 9.0 (100) | Lowland | ||
| Punta Banco–Burica (0–100), PS | November–December 2011 | 68.6 (35) | Lowland | [63] |
| Rincón de Osa (0–100), PS | 0.0 (25) | Lowland | ||
| Puerto Viejo de Sarapiquí (0–200), CL | 67.4 (144) | Lowland | ||
| Guayacán de Siquirres (400–600), CL | 47.9 (144) | Lowland | ||
| San Vito de Coto Brus (1120–1385), MSCC | Unknown/not indicated | 10.5 (19) | Lowland | [65] |
| Punta Banco–Burica (0–100), PS | 0.0 (20) | Lowland | ||
| Guayacán de Siquirres (400–600), CL | 5.3 (19) | Lowland | ||
| San Rafael de Heredia (1800), MSCC | 66.7 (15) | Lower montane | ||
| Santo Domingo de Heredia (1000–1200), MSCC | 45.5 (11) | Premontane | ||
| Las Tablas (1350), MSCC | 28.6 (14) | Lower montane |
| FRHI | Species | Taxonomic Group (Examples) |
|---|---|---|
| DAA | 2 | Diasporus spp. (dink frogs, e.g., Diasporus diastema) |
| DAT | 5 | Pristimantis spp. (rain frogs, e.g., Pristimantis cerasinus) |
| DRT | 3 | Craugastor punctariolus clade (robber frogs, e.g., Craugastor taurus) |
| C. fitzingeri (Pacific side) | ||
| DTT | 13 | Craugastor spp. (leaf-litter frogs. e.g., Craugastor bransfordi) |
| C. fitzingeri (Caribbean side) | ||
| Plethodontidae (e.g., Oedipina gracilis) | ||
| IAP | 17 | Hylidae (pond-breeding treefrogs, e.g., Boana rufitela) |
| IAR | 15 | Centrolenidae (glass frogs, e.g., Teratohyla pulverata) |
| Hylidae (stream-breeding treefrogs, e.g., Duellmanohyla rufioculis) | ||
| ITF | 4 | Dendrobatidae (Poison-dart frogs, e.g., Oophaga pumilio) |
| ITP | 12 | Leptodactylidae (Leptodactylid frogs, e.g., Leptodactylus melanonotus) |
| Microhylidae (sheep frogs, e.g., Hypopachus variolosus) | ||
| Ranidae (Ranid frogs, e.g., Lithobates forreri) | ||
| Bufonidae (toads, e.g., Incilius coccifer) | ||
| ITR | 7 | Bufonidae (river toads, e.g., Rhaebo haematiticus) |
| Order | Family | Species | Source |
|---|---|---|---|
| Anura | Centrolenidae | Hyalinobatrachium dianae | [75] |
| Craugastoridae | Craugastor aenigmaticus | [76] | |
| Craugastor gabbi | [77] | ||
| Craugastor zunigai | [78] | ||
| Eleutherodactylidae | Diasporus amirae | [79] | |
| Eleutherodactylus planirostris * | [80] | ||
| Hylidae | Ecnomiohyla bailarina | [81] | |
| Ecnomiohyla veraguensis | Unpublished | ||
| Smilisca manisorum | [82] | ||
| Phyllomedusidae | Cruziohyla sylviae | [83] | |
| Caudata | Plethodontidae | Bolitoglossa aurae | [84] |
| Bolitoglossa aureogularis | [85] | ||
| Bolitoglossa kamuk | [85] | ||
| Bolitoglossa pygmaea | Unpublished | ||
| Bolitoglossa splendida | [85] | ||
| Nototriton costaricense | [86] | ||
| Nototriton matama | [85] | ||
| Oedipina berlini | [87] | ||
| Oedipina nimaso | [85] | ||
| Gymnophiona | Caeciliidae | Caecilia volcani | [88] |
| Species (FRHI) | Bd + Swabs | Bd Load Average (SE) | Log10 Bd Load Average (SE) |
|---|---|---|---|
| Agalychnis callidryas (IAP) | 4 | 8.19 (3.81) | 0.77 (0.21) |
| Agalychnis spurrelli (IAP) | 5 | 39.83 (32.47) | 1.10 (0.30) |
| Boana rufitela (IAP) | 8 | 8.41 (4.12) | 0.53 (0.23) |
| Bolitoglossa colonnea (DTT) | 1 | 1.83 (0.00) | 0.26 (0.00) |
| Cochranella granulosa (IAR) | 1 | 3.95 (0.00) | 0.60 (0.00) |
| Craugastor bransfordi (DTT) | 23 | 1007.06 (483.50) | 1.78 (0.25) |
| Craugastor crassidigitus (DTT) | 5 | 1636.74 (1583.43) | 1.69 (0.64) |
| Craugastor fitzingeri (DTT, DRT) | 44 | 951.48 (310.27) | 1.97 (0.17) |
| Craugastor megacephalus (DTT) | 1 | 0.62 (0.00) | −0.21 (0.00) |
| Craugastor mimus (DTT) | 9 | 125.48 (74.69) | 1.01 (0.46) |
| Craugastor ranoides (DRT) | 3 | 187.40 (174.13) | 1.65 (0.55) |
| Craugastor stejnegerianus (DTT) | 2 | 2.18 (0.95) | 0.29 (0.20) |
| Craugastor taurus (DRT) | 12 | 11,632.50 (6564.67) | 2.51 (0.41) |
| Dendropsophus ebraccatus (IAP) | 34 | 315.85 (194.09) | 1.00 (0.18) |
| Diasporus diastema (DAA) | 2 | 14.44 (4.12) | 1.14 (0.13) |
| Duellmanohyla rufioculis (IAR) | 1 | 3.65 (0.00) | 0.56 (0.00) |
| Engystomops pustulosus (ITP) | 11 | 34.83 (13.26) | 1.11 (0.3) |
| Espadarana prosoblepon (IAR) | 3 | 3691.59 (3684.73) | 1.06 (1.75) |
| Hyalinobatrachium colymbiphyllum (IAR) | 1 | 0.01 (0.00) | −2.00 (0.00) |
| Hyalinobatrachium valerioi (IAR) | 2 | 8.38 (2.04) | 0.91 (0.11) |
| Incilius melanochlorus (ITR) | 2 | 23.27 (19.93) | 1.08 (0.56) |
| Leptodactylus melanonotus (ITP) | 4 | 11.86 (0.66) | 1.07 (0.02) |
| Leptodactylus poecilochilus (ITP) | 1 | 1073.45 (0.00) | 3.03 (0.00) |
| Leptodactylus savagei (ITP) | 1 | 33.49 (0.00) | 1.52 (0.00) |
| Lithobates forreri (ITP) | 2 | 569.24 (241.10) | 2.71 (0.20) |
| Lithobates warszewitschii (ITR) | 14 | 978.92 (801.60) | 1.47 (0.31) |
| Oophaga granulifera (ITF) | 9 | 23.92 (11.31) | 1.20 (0.11) |
| Oophaga pumilio (ITF) | 34 | 1765.81 (778.67) | 1.71 (0.25) |
| Pristimantis cerasinus (DAT) | 9 | 14.82 (10.97) | 0.47 (0.32) |
| Pristimantis ridens (DAT) | 7 | 48.37 (32.34) | 0.69 (0.50) |
| Rhaebo haematiticus (ITR) | 22 | 239.20 (178.56) | 0.70 (0.26) |
| Scinax boulengeri (IAP) | 1 | 195.20 (0.00) | 2.29 (0.00) |
| Scinax elaeochroa (IAP) | 5 | 1384.15 (1350.51) | 1.78 (0.58) |
| Smilisca phaeota (IAP) | 4 | 37.25 (19.15) | 1.44 (0.18) |
| Smilisca sordida (IAP) | 46 | 14.96 (9.27) | 0.24 (0.16) |
| Teratohyla pulverata (IAR) | 2 | 34.53 (22.92) | 1.41 (0.35) |
| Teratohyla spinosa (IAR) | 5 | 937.99 (825.54) | 1.90 (0.57) |
| Tlalocohyla loquax (IAP) | 11 | 144.66 (107.28) | 1.22 (0.30) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zumbado-Ulate, H.; Nelson, K.N.; García-Rodríguez, A.; Chaves, G.; Arias, E.; Bolaños, F.; Whitfield, S.M.; Searle, C.L. Endemic Infection of Batrachochytrium dendrobatidis in Costa Rica: Implications for Amphibian Conservation at Regional and Species Level. Diversity 2019, 11, 129. https://doi.org/10.3390/d11080129
Zumbado-Ulate H, Nelson KN, García-Rodríguez A, Chaves G, Arias E, Bolaños F, Whitfield SM, Searle CL. Endemic Infection of Batrachochytrium dendrobatidis in Costa Rica: Implications for Amphibian Conservation at Regional and Species Level. Diversity. 2019; 11(8):129. https://doi.org/10.3390/d11080129
Chicago/Turabian StyleZumbado-Ulate, Héctor, Kiersten N. Nelson, Adrián García-Rodríguez, Gerardo Chaves, Erick Arias, Federico Bolaños, Steven M. Whitfield, and Catherine L. Searle. 2019. "Endemic Infection of Batrachochytrium dendrobatidis in Costa Rica: Implications for Amphibian Conservation at Regional and Species Level" Diversity 11, no. 8: 129. https://doi.org/10.3390/d11080129
APA StyleZumbado-Ulate, H., Nelson, K. N., García-Rodríguez, A., Chaves, G., Arias, E., Bolaños, F., Whitfield, S. M., & Searle, C. L. (2019). Endemic Infection of Batrachochytrium dendrobatidis in Costa Rica: Implications for Amphibian Conservation at Regional and Species Level. Diversity, 11(8), 129. https://doi.org/10.3390/d11080129