Genetic Data Suggest Multiple Introductions of the Lionfish (Pterois miles) into the Mediterranean Sea

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
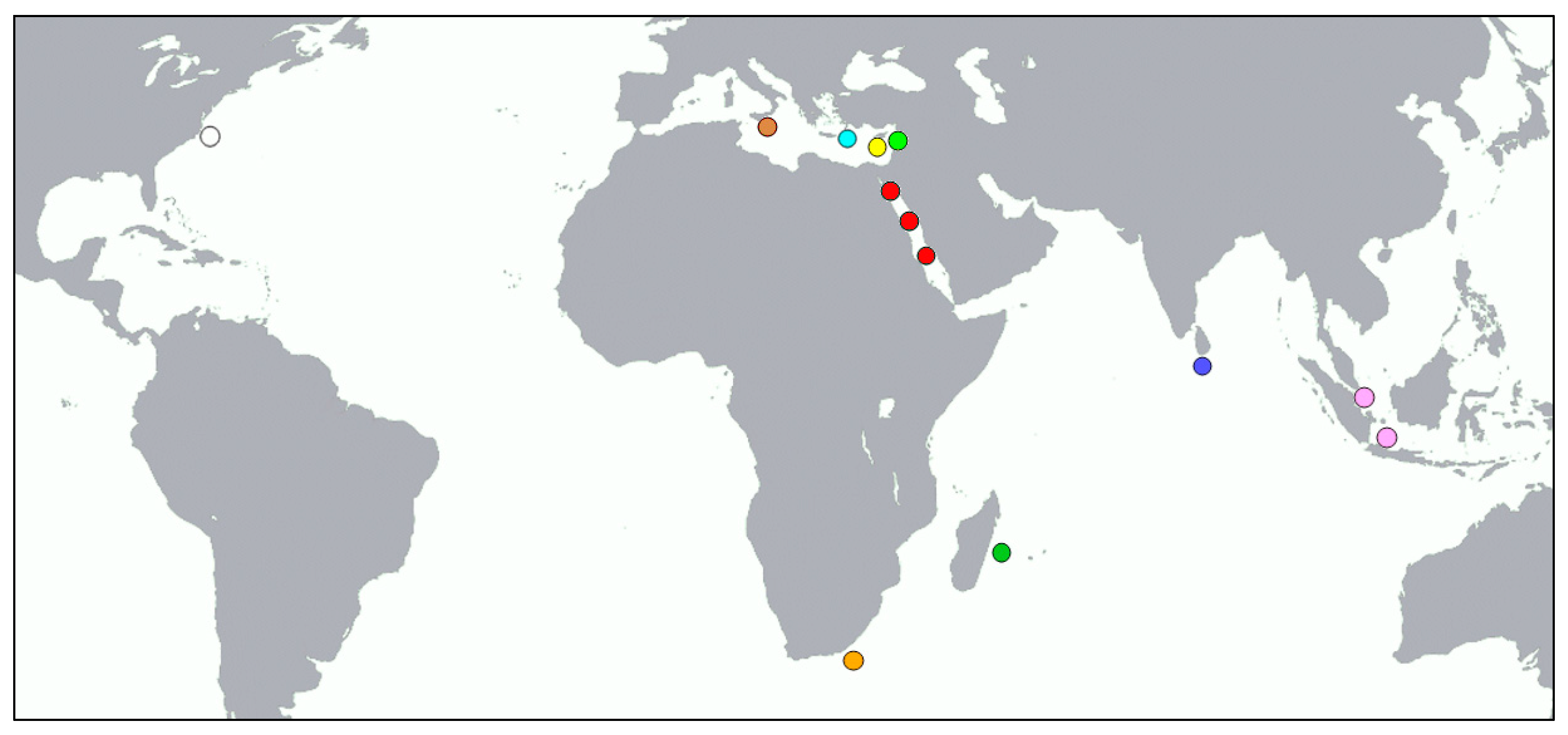
2.1. Sampling—DNA Extraction
2.2. Amplification and Sequencing
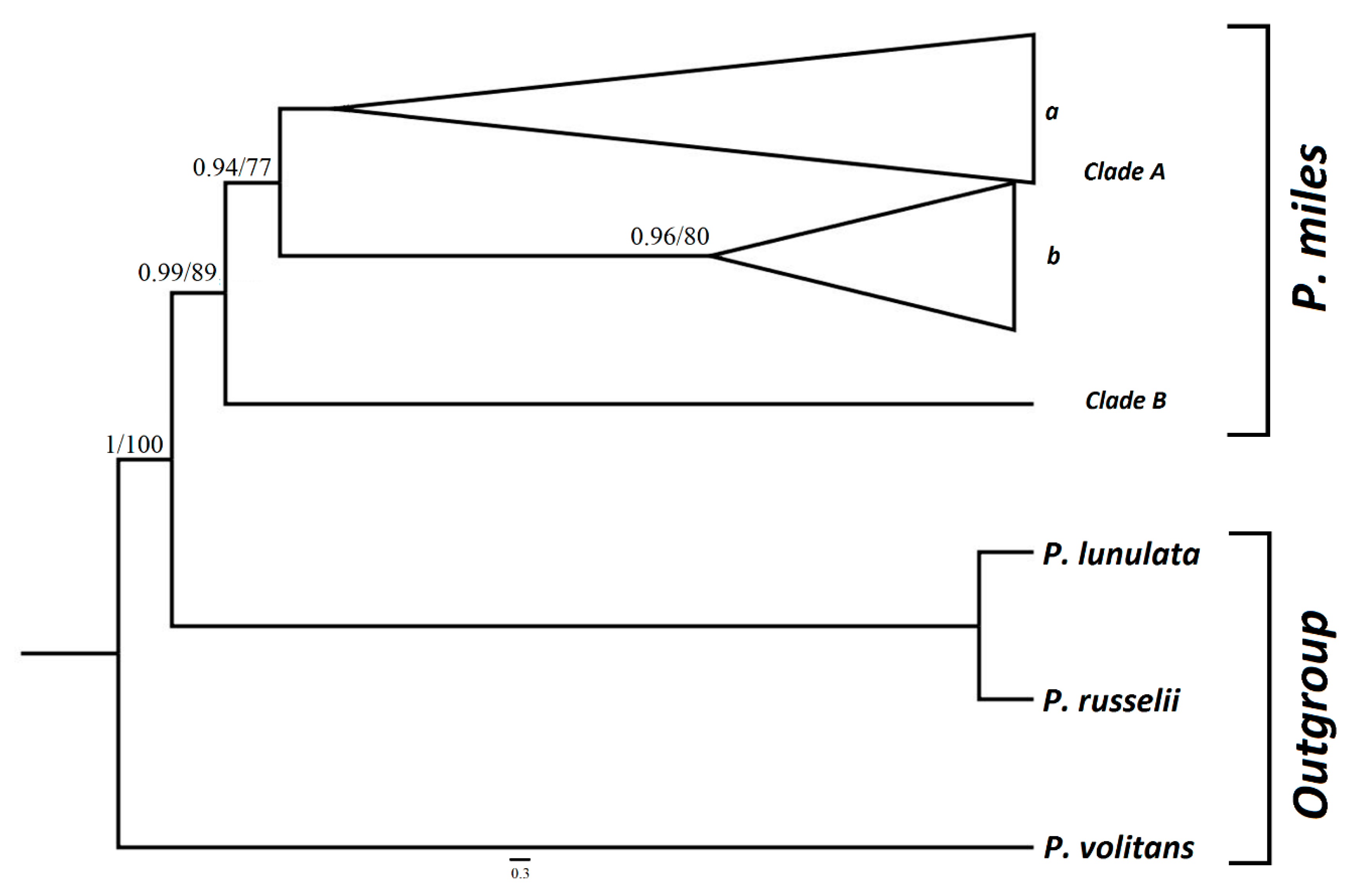
2.3. Data Elaboration and Phylogenetic Analyses
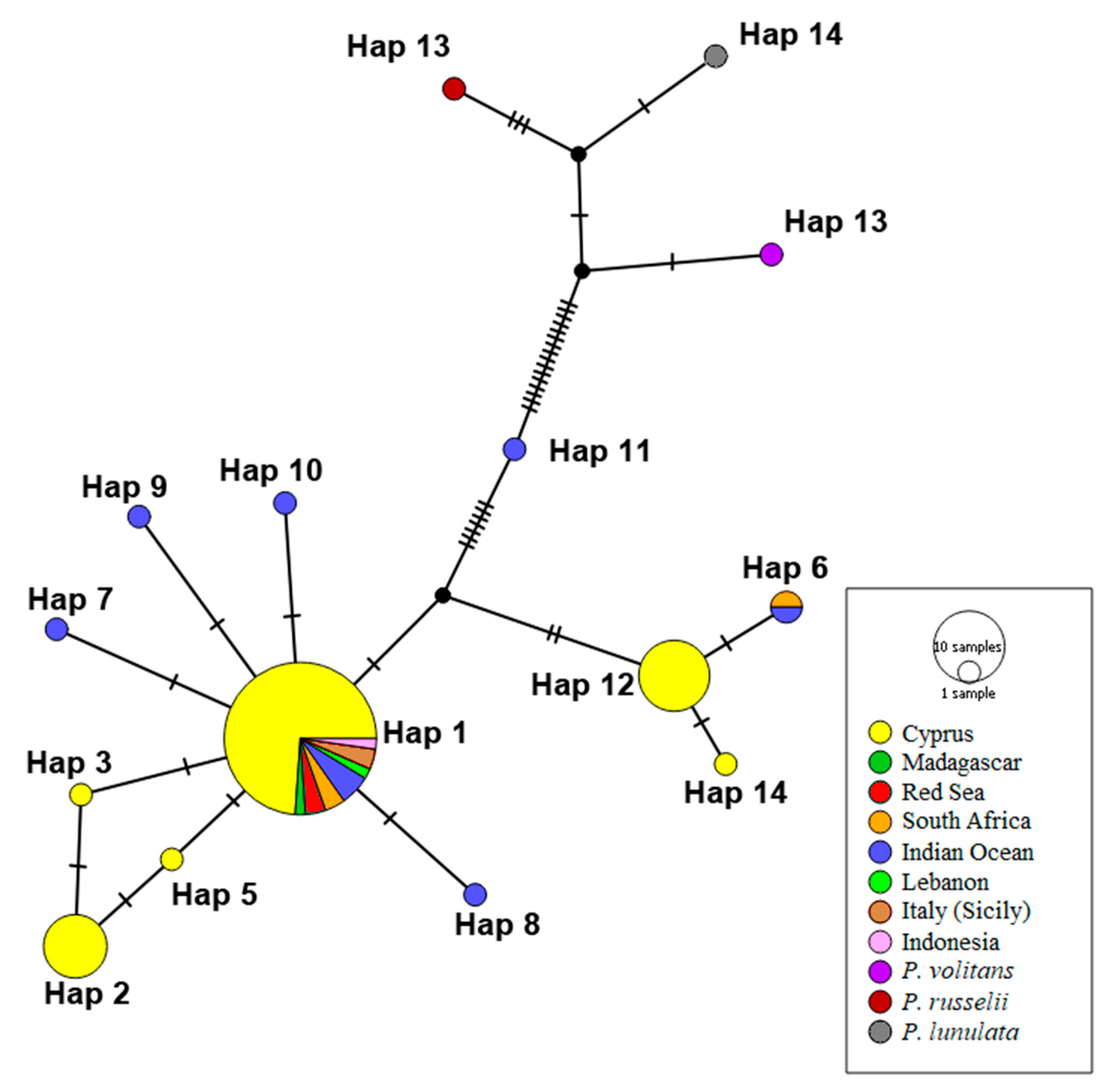
2.4. Geographic Population Structure—Haplotype Networks Construction
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mack, R.N.; Simberloff, D.; Mark Lonsdale, W.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions, causes, epidemiology, global consequences and control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- Bax, N.; Williamson, A.; Aguero, M.; Gonzalez, E.; Geeves, W. Marine invasive alien species: A threat to global biodiversity. Mar. Policy 2003, 27, 313–323. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Wallentinus, I.; Zenetos, A.; Leppäkoski, E.; Çinar, M.E.; Oztürk, B.; Grabowski, M.; Golani, D.; Cardoso, A.C. Impacts of invasive alien marine species on ecosystem services and biodiversity: A pan-European review. Aquat. Invasions 2014, 9, 391–423. [Google Scholar] [CrossRef]
- Lovell, S.J.; Stone, S.F.; Fernandez, L. The economic impacts of aquatic invasive species: A review of the literature. Agric. Resour. Econ. 2006, 35, 195–208. [Google Scholar] [CrossRef]
- Molnar, J.L.; Gamboa, R.L.; Revenga, C.; Spalding, M.D. Assessing the global threat of invasive species to marine biodiversity. Front. Ecol. Environ. 2008, 6, 458–492. [Google Scholar] [CrossRef]
- Vilà, M.; Basnou, C.; Pyšek, P.; Josefsson, M.; Genovesi, P.; Gollasch, S.; Nentwig, W.; Olenin, S.; Roques, A.; Roy, D.; et al. How well do we understand the impacts of alien species on ecosystem services? A pan-European, cross-taxa assessment. Front. Ecol. Environ. 2010, 8, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Azzurro, E.; Bariche, M. Local knowledge and awareness on the incipient lionfish invasion in the eastern Mediterranean Sea. Mar. Freshw. Res. 2017, 68, 1950–1954. [Google Scholar] [CrossRef] [Green Version]
- Galil, B. Eyes wide shut: Managing bio-invasions in Mediterranean marine protected areas. In Management of Marine Protected Areas: A Network Perspective; Goriup, P.D., Ed.; Wiley: New York, NY, USA, 2017; pp. 187–206. [Google Scholar] [CrossRef]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Lasram, F.B.R.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The Biodiversity of the Mediterranean Sea: Estimates, Patterns and Threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef]
- Zenetos, A.; Çinar, M.E.; Crocetta, F.; Golani, D.; Rosso, A.; Servello, G.; Shenkar, N.; Turon, X.; Verlaque, M. Uncertainties and validation of alien species catalogues: The Mediterranean as an example. Estuar. Coast. Shelf. Sci. 2017, 191, 171–187. [Google Scholar] [CrossRef]
- Por, F.D. One hundred years of the Suez Canal—A century of Lessepsian migration: Retrospects and viewpoints. Syst. Zool. 1971, 20, 138–195. [Google Scholar] [CrossRef]
- Galil, B.S.; Boero, F.; Campbell, M.L.; Carlton, J.T.; Cook, E.; Fraschetti, S.; Gollasch, S.; Hewitt, C.L.; Jelmert, A.; Macpherson, E.; et al. ‘Double trouble’: The expansion of the Suez Canal and marine bioinvasions in the Mediterranean Sea. Biol. Invasions 2015, 17, 973–976. [Google Scholar] [CrossRef]
- Galil, B.S. Taking stock: Inventory of alien species in the Mediterranean Sea. Biol. Invasions 2009, 11, 359–372. [Google Scholar] [CrossRef]
- Galil, B.S.; Marchini, A.; Occhipinti-Ambrogi, A. East is east and West is west? Management of marine bioinvasions in the Mediterranean Sea. Estuar. Coast. Shelf. Sci. 2016, 201, 7–16. [Google Scholar] [CrossRef]
- García-Berthou, E. The characteristics of invasive fishes: What has been learned so far? J. Fish. Biol. 2007, 71, 33–55. [Google Scholar] [CrossRef]
- Côté, I.M.; Smith, N.S. The lionfish Pterois sp. invasion: Has the worst-case scenario come to pass? J. Fish. Biol. 2018, 92, 660–689. [Google Scholar] [CrossRef]
- Schultz, E. Pterois volitans and Pterois miles: Two Valid Species. Copeia 1986, 3, 686–690. [Google Scholar] [CrossRef]
- Albins, M.A.; Hixon, M.A. Worst case scenario: Potential long-term effects of invasive predatory lionfish (Pterois volitans) on Atlantic and Caribbean coral-reef communities. Environ. Biol. Fishes 2013, 96, 1151–1157. [Google Scholar] [CrossRef]
- Schofield, P.J. Geographic extent and chronology of the invasion of non-native lionfish (Pterois volitans [Linnaeus 1758] and P. miles [Bennett 1828]) in the western North Atlantic and Caribbean Sea. Aquat. Invasions 2009, 4, 473–479. [Google Scholar] [CrossRef]
- Whitfield, P.E.; Hare, J.A.; David, A.W.; Harter, S.L.; Muñoz, R.C.; Addison, C.M. Abundance estimates of the Indo-Pacific lionfish Pterois volitans/miles complex in the western North Atlantic. Biol. Invasions 2007, 9, 53–64. [Google Scholar] [CrossRef]
- Ferreira, C.E.L.; Luiz, O.J.; Floeter, S.R.; Lucena, M.B.; Barbosa, M.C.; Rocha, C.R.; Rocha, L.A. First record of invasive lionfish (Pterois volitans) for the Brazilian Coast. PLoS ONE 2015, 10, e0123002. [Google Scholar] [CrossRef]
- Hamner, R.M.; Freshwater, D.W.; Whitfield, P.E. Mitochondrial cytochrome b analysis reveals two invasive lionfish species with strong founder effects in the western Atlantic. J. Fish Biol. 2007, 71, 214–222. [Google Scholar] [CrossRef]
- Johnston, M.W.; Purkis, S.J. Spatial analysis of the invasion of lionfish in the western Atlantic and Caribbean. Mar. Pollut. Bull. 2011, 62, 1218–1226. [Google Scholar] [CrossRef]
- Bariche, M.; Kleitou, P.; Kalogirou, S.; Bernardi, G. Genetics reveal the identity and origin of the lionfish invasion in the Mediterranean Sea. Sci. Rep. 2017, 7, 6782. [Google Scholar] [CrossRef] [Green Version]
- Stern, N.; Jimenez, C.; Huseyinoglu, M.F.; Andreou, V.; Hadijioannou, L.; Petrou, A.; Öztürk, B.; Golani, D.; Rothman, S.B. Constructing the genetic population demography of the invasive lionfish Pterois miles in the Levant Basin, Eastern Mediterranean. Mitochondrial DNA Part A 2018, 30, 249–255. [Google Scholar] [CrossRef]
- Selwyn, J.D.; Johnson, J.E.; Downey-Wall, A.M.; Bynum, A.M.; Hamner, R.M.; Hogan, J.D.; Bird, C.E. Simulations indicate that scores of lionfish (Pterois volitans) colonized the Atlantic Ocean. Peer J. 2017, 5, e3996. [Google Scholar] [CrossRef]
- Johnson, J.; Bird, C.E.; Johnston, M.A.; Fogg, A.Q.; Hogan, J.D. Regional genetic structure and genetic founder effects in the invasive lionfish: Comparing the Gulf of Mexico, Caribbean and North Atlantic. Mar. Biol. 2016, 163, 216. [Google Scholar] [CrossRef]
- Wilcox, C.L.; Motomura, H.; Matsunuma, M.; Bowen, B.W. Phylogeography of lionfishes (Pterois) indicate taxonomic over splitting and hybrid origin of the invasive Pterois Volitans. J. Hered. 2017, 109, 162–175. [Google Scholar] [CrossRef]
- Bos, A.R.; Grubich, J.R.; Sanad, A.M. Growth, site fidelity and grouper interactions of the Red Sea lionfish Pterois miles (Scorpaenidae) in its native habitat. Mar. Biol. 2018, 165, 175. [Google Scholar] [CrossRef]
- Bos, A.R.; Sanad, A.M.; Elsayed, K. Gymnothorax spp.(Muraenidae) as natural predators of the lionfish Pterois miles in its native biogeographical range. Environ. Biol. Fishes 2017, 100, 745–748. [Google Scholar] [CrossRef]
- Golani, D.; Sonin, O. New records of the Red Sea fishes, Pterois miles (Scorpaenidae) and Pteragogus pelycus (Labridae) from the eastern Mediterranean Sea. Jpn. J. Ichthyol. 1992, 39, 167–169. [Google Scholar] [CrossRef]
- Bariche, M.; Torres, M.; Azzurro, E. The presence of the invasive Lionfish Pterois miles in the Mediterranean Sea. Mediterr. Mar. Sci. 2013, 14, 292–294. [Google Scholar] [CrossRef]
- Jimenez, C.; Petrou, A.; Andreou, V.; Hadjioannou, L.; Wolf, W.; Koutsoloukas, N.; Abu Alhaija, R. Veni, vidi, vici: The successful establishment of the lionfish Pterois miles in Cyprus (Levantine Sea). Rapp. Comm. Int. Mer Mediterr. 2016, 41, 417. [Google Scholar]
- Kletou, D.; Hall-Spencer, J.M.; Kleitou, P. A lionfish (Pterois miles) invasion has begun in the Mediterranean Sea. Mar. Biodivers. Rec. 2016, 9, 46. [Google Scholar] [CrossRef]
- Baldwin, C.C.; Mounts, J.H.; Smith, D.G.; Weigt, L.A. Genetic identification and color descriptions of early life-history stages of Belizean Phaeoptyx and Astrapogon (Teleostei: Apogonidae) with comments on identification of adult Phaeoptyx. Zootaxa 2009, 26, 1–22. [Google Scholar]
- Handy, S.M.; Deeds, J.R.; Ivanova, N.V.; Hebert, P.D.; Hanner, R.H.; Ormos, A.; Weigt, L.A.; Moore, M.M.; Yancy, H.F. A single-laboratory validated method for the generation of DNA barcodes for the identification of fish for regulatory compliance. J. AOAC Int. 2011, 94, 201–210. [Google Scholar]
- Lee, W.J.; Conroy, J.; Howell, W.H.; Kocher, T.D. Structure and evolution of teleost mitochondrial control regions. J. Mol. Evol. 1995, 41, 54–66. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Yang, Z. Computational Molecular Evolution; Oxford University Press: Oxford, UK, 2006. [Google Scholar]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. Mrbayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J. Tracer 1.5.0; University of Edinburgh: Edinburgh, UK, 2007; Available online: http://beast.Bio.Ed.Ac.uk/Tracer (accessed on 1 June 2019).
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. PopART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Bandelt, H.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Lopez, J.V.; Yuhki, N.; Masuda, R.; Modi, W.; O’Brien, S.J. Numt, a recent transfer and tandem amplification of mitochondrial DNA to the nuclear genome of the domestic cat. J. Molec. Evol. 1994, 39, 174–190. [Google Scholar] [CrossRef]
- Akaike, H. New look at statistical-model identification. IEEE Trans. Automat. Contr. 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Ahrenholz, D.W.; Morris, J.A. Larval duration of the lionfish, Pterois volitans along the Bahamian Archipelago. Environ. Biol. Fishes 2010, 88, 305–309. [Google Scholar] [CrossRef]
- Whitfield, P.E.; Gardner, T.; Vives, S.P.; Gilligan, M.R.; Courtenay, W.R., Jr.; Ray, G.C.; Hare, J.A. Biological invasion of the Indo-Pacific lionfish (Pterois volitans) along the Atlantic coast of North America. Mar. Ecol. Prog. Ser. 2002, 235, 289–297. [Google Scholar] [CrossRef]
- Semmens, B.X.; Buhle, E.R.; Salomon, A.K.; Pattengill-Semmens, C.V. A hotspot of non-native marine fishes: evidence for the aquarium trade as an invasion pathway. Mar. Ecol. Prog. Ser. 2004, 266, 239–244. [Google Scholar] [CrossRef] [Green Version]
- Bariche, M.; Kazanjian, G.; Azzurro, E.A. A lag of 25 years: Evidence from an old capture of Fistularia commersonii Ruppell, 1838 from Lebanon (Mediterranean Sea). J. Appl. Ichthyol. 2014, 30, 535–536. [Google Scholar] [CrossRef]
- Zenetos, A.; Gofas, S.; Verlaque, M.; Cinar, M.E.; García-Raso, J.E.; Bianchi, C.N.; Morri, C.; Azzurro, E.; Bilecenoglu, M.; Froglia, C.; et al. Alien species in the Mediterranean Sea by. A contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part 2. Introduction trends and pathways. Mediterr. Mar. Sci. 2012, 13, 328–352. [Google Scholar] [CrossRef]
- Kitchens, L.L.; Paris, C.B.; Vaz, A.C.; Ditty, J.G.; Cornic, M.; Cowan, J.H.; Rooker, J.R. Occurrence of invasive lionfish (Pterois volitans) larvae in the northern Gulf of Mexico: Characterization of dispersal pathways and spawning areas. Biol. Invasions 2017, 19, 1971–1979. [Google Scholar] [CrossRef]
- Shefer, S.; Abelson, A.; Mokady, O.; Geffen, E.L. Red to Mediterranean Sea bioinvasion: Natural drift through the Suez Canal, or anthropogenic transport? Mol. Ecol. 2004, 13, 2333–2343. [Google Scholar] [CrossRef]
- MacIsaac, H.J.; De Roy, E.M.; Leung, B.; Grgicak-Mannion, A.; Ruiz, G.M. Possible ballast water transfer of Lionfish to the eastern Pacific Ocean. PLoS ONE 2016, 11, e0165584. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Cyprus | 0.26 | ||||||||||
| 2 | Lebanon | 0.16 | n/c | |||||||||
| 3 | Italy (Sicily) | 0.16 | 0.00 | 0.00 | ||||||||
| 4 | Gulf of Aqaba | 0.24 | 0.08 | 0.08 | 0.16 | |||||||
| 5 | Indian ocean | 0.53 | 0.38 | 0.38 | 0.40 | 0.74 | ||||||
| 6 | South Africa | 0.36 | 0.28 | 0.28 | 0.36 | 0.58 | 0.55 | |||||
| 7 | Madagascar | 0.15 | 0.00 | 0.00 | 0.08 | 0.38 | 0.27 | n/c | ||||
| 8 | Indonesia | 0.16 | 0.00 | 0.00 | 0.09 | 0.38 | 0.29 | 0.00 | n/c | |||
| 9 | P. russelii | 4.86 | 4.66 | 4.55 | 4.85 | 5.06 | 4.82 | 4.77 | 4.70 | n/c | ||
| 10 | P. volitans | 4.67 | 4.49 | 4.38 | 4.69 | 4.96 | 4.66 | 4.58 | 4.53 | 0.99 | n/c | |
| 11 | P. lunulata | 4.38 | 4.28 | 4.28 | 4.37 | 4.53 | 4.28 | 4.28 | 4.28 | 0.93 | 0.56 | n/c |
| Group | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Cyprus | 1.06 | ||||||||
| 2 | Lebanon | 0.73 | n/c | |||||||
| 3 | Red Sea | 2.67 | 1.61 | 2.01 | ||||||
| 4 | Rhodes | 0.73 | 0.00 | 1.61 | n/c | |||||
| 5 | Gulf of Aqaba | 2.78 | 1.78 | 2.04 | 1.78 | 2.13 | ||||
| 6 | Indian Ocean | 2.77 | 1.96 | 2.21 | 1.96 | 2.42 | 2.32 | |||
| 7 | Indonesia | 0.96 | 0.00 | 0.77 | 0.00 | 1.03 | 1.73 | n/c | ||
| 8 | USA | 1.54 | 1.04 | 2.05 | 1.04 | 1.97 | 2.88 | 0.98 | n/c | |
| 9 | P. volitans | 8.72 | 9.42 | 14.86 | 9.42 | 14.75 | 14.72 | 10.84 | 10.84 | n/c |
| Source of Variation | df | Sum of Squares | % of Variation | FST | p-Value |
|---|---|---|---|---|---|
| Among populations | 2 | 5.032 | 18.47 | 0.185 | >0.001 |
| Within populations | 102 | 38.549 | 81.53 | ||
| Total | 104 | 43.581 | 100 |
| Group | 1 | 2 | 3 |
|---|---|---|---|
| 1. Mediterranean | + | + | |
| 2. Indian Ocean | 0.23 | − | |
| 3. Red Sea | 0.20 | >0.01 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dimitriou, A.C.; Chartosia, N.; Hall-Spencer, J.M.; Kleitou, P.; Jimenez, C.; Antoniou, C.; Hadjioannou, L.; Kletou, D.; Sfenthourakis, S. Genetic Data Suggest Multiple Introductions of the Lionfish (Pterois miles) into the Mediterranean Sea. Diversity 2019, 11, 149. https://doi.org/10.3390/d11090149
Dimitriou AC, Chartosia N, Hall-Spencer JM, Kleitou P, Jimenez C, Antoniou C, Hadjioannou L, Kletou D, Sfenthourakis S. Genetic Data Suggest Multiple Introductions of the Lionfish (Pterois miles) into the Mediterranean Sea. Diversity. 2019; 11(9):149. https://doi.org/10.3390/d11090149
Chicago/Turabian StyleDimitriou, Andreas C., Niki Chartosia, Jason M. Hall-Spencer, Periklis Kleitou, Carlos Jimenez, Charalampos Antoniou, Louis Hadjioannou, Demetris Kletou, and Spyros Sfenthourakis. 2019. "Genetic Data Suggest Multiple Introductions of the Lionfish (Pterois miles) into the Mediterranean Sea" Diversity 11, no. 9: 149. https://doi.org/10.3390/d11090149
APA StyleDimitriou, A. C., Chartosia, N., Hall-Spencer, J. M., Kleitou, P., Jimenez, C., Antoniou, C., Hadjioannou, L., Kletou, D., & Sfenthourakis, S. (2019). Genetic Data Suggest Multiple Introductions of the Lionfish (Pterois miles) into the Mediterranean Sea. Diversity, 11(9), 149. https://doi.org/10.3390/d11090149