Barcoding Analysis of Paraguayan Squamata



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
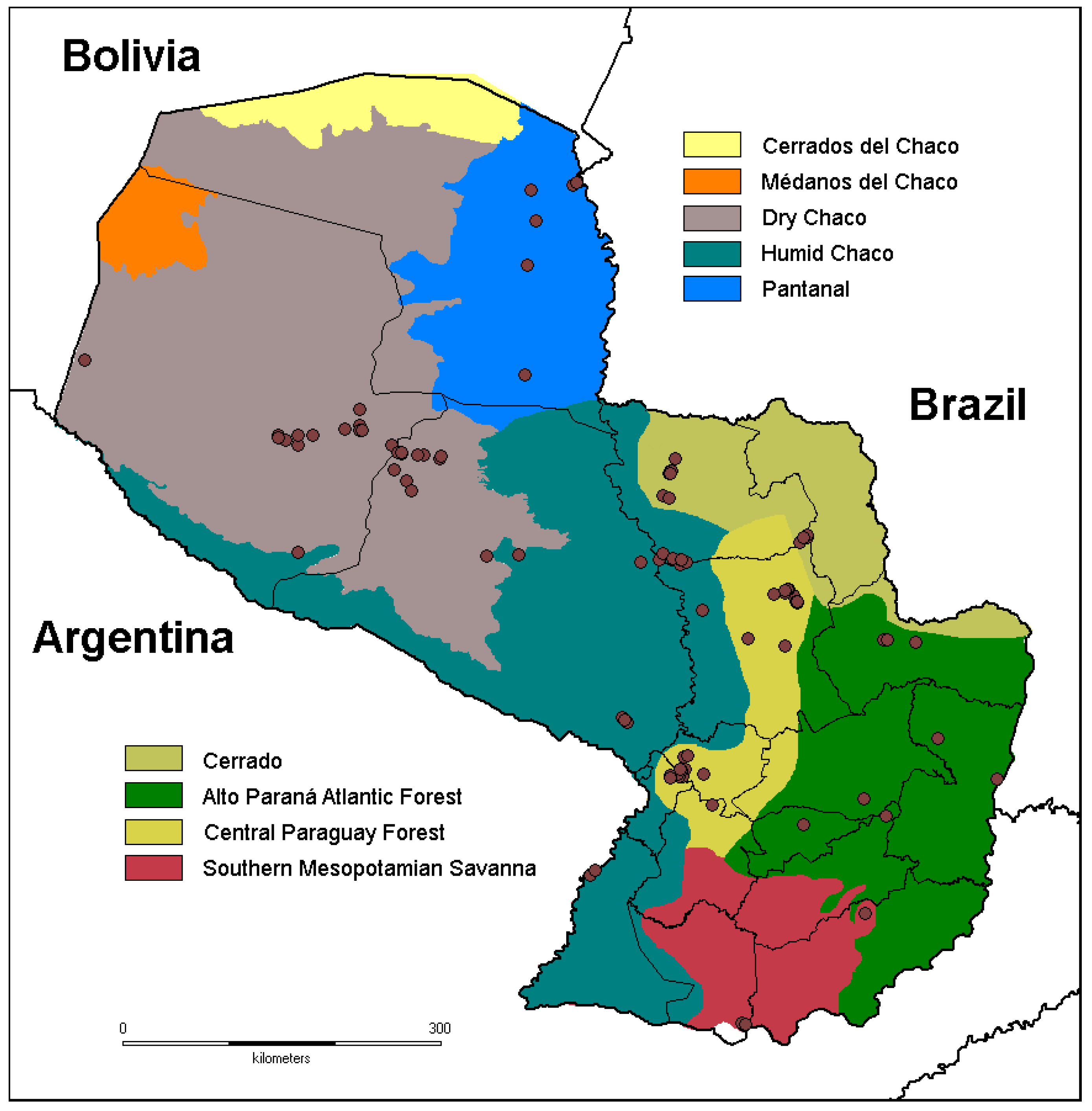
2.1. Study Area
2.2. Data Collection
2.3. Molecular Protocols
2.4. Data Analysis
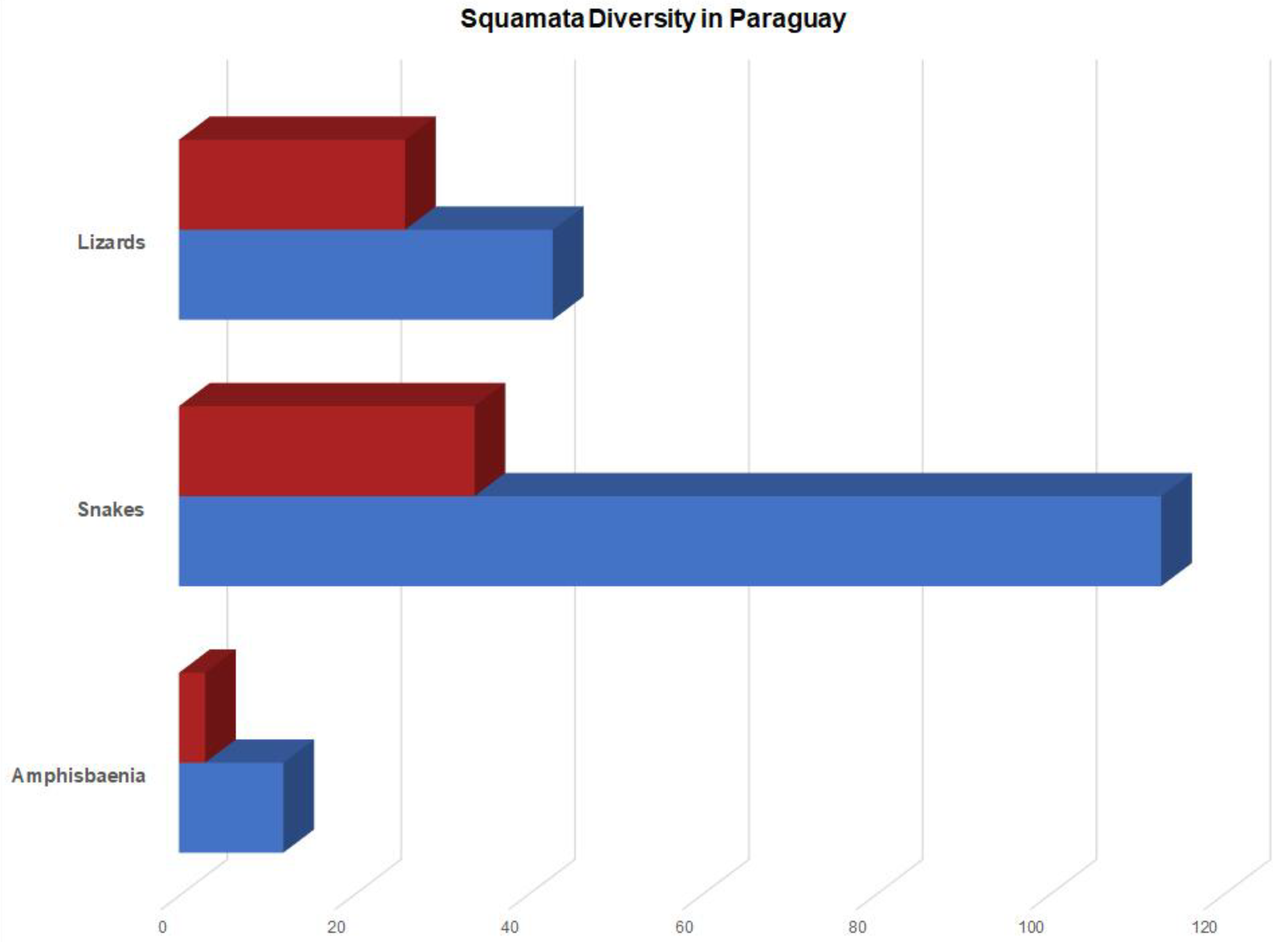
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pullin, A.; Sutherland, W.; Gardner, T.; Kapos, V.; Fa, J.E. Conservation priorities: Identifying need, taking action and evaluating success. In Key Topics in Conservation Biology 2; Macdonald, D.W., Willis, K.J., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 3–22. [Google Scholar] [CrossRef]
- Baumgartner, J.; Esselman, R.; Salzer, D.; Young, J. Conservation Action Planning Handbook; The Nature Conservancy: Arlington, VA, USA, 2007. [Google Scholar]
- Ripple, W.J.; Wolf, C.; Newsome, T.M.; Galetti, M.; Alamgir, M.; Crist, E.; Mahmoud, M.I.; Laurance, W.F. World scientists’ warning to humanity: A second warning. Bioscience 2017, 67, 1026–1028. [Google Scholar] [CrossRef]
- Young, B.E.; Stuart, S.N.; Chanson, J.S.; Cox, N.A.; Boucher, T.M. Disappearing Jewels: The Status of New World Amphibians; NatureServe: Arlington, VA, USA, 2004. [Google Scholar]
- Johnson, C.N.; Balmford, A.; Brook, B.W.; Buettel, J.C.; Galetti, M.; Guangchun, L.; Wilmshurst, J.M. Biodiversity losses and conservation responses in the Anthropocene. Science 2017, 356, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Palumbi, S.R.; Martin, A.; Romano, S.; Mcmillan, W.O.; Stice, L.; Grabowski, G. The Simple Fool’s Guide to PCR; University of Hawaii Press: Honolulu, HI, USA, 1991. [Google Scholar]
- Rudnick, J.A.; Katzner, T.E.; Bragin, E.A.; DeWoody, J.A. Species identification of 548 birds through genetic analysis of naturally shed feathers. Mol. Ecol. Notes 2007, 7, 757–762. [Google Scholar] [CrossRef]
- Yang, L.; Tan, Z.; Wang, D.; Xue, L.; Guan, M.X.; Huang, T.; Li, R. Species identification through mitochondrial rRNA genetic analysis. Sci. Rep. 2014, 4, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sites, J.; Marshall, J.C. Delimiting species: A renaissance issue in systematic biology. Trends. Ecol. Evol. 2003, 18, 462–470. [Google Scholar] [CrossRef]
- Pinzón, J.H.; LaJaunesse, T.C. Species delimitation of common reef corals in the genus Pocillopora using nucleotide sequence phylogenies, population genetics and symbiosis ecology. Mol. Ecol. 2011, 20, 311–325. [Google Scholar] [CrossRef] [PubMed]
- Khodami, S.; Martínez Arbizu, P.; Stöhr, S.; Laakmann, S. Molecular species delimitation of Icelandic brittle stars (Ophiuroidea). Pol. Polar Res. 2014, 35, 243–260. [Google Scholar] [CrossRef]
- Leliaert, F.; Verbruggenc, H.; Vanormelingend, P.; Steena, F.; López-Bautistab, J.M.; Zuccarelloe, G.C.; De Clercka, O. DNA-based species delimitation in algae. Eur. J. Phycol. 2014, 49, 179–196. [Google Scholar] [CrossRef] [Green Version]
- Barcaccia, G.; Lucchini, M.; Cassandro, M. DNA Barcoding as a molecular tool to track down mislabeling and food piracy. Diversity 2016, 8, 2. [Google Scholar] [CrossRef]
- Lane, N. On the origin of bar codes. Nature 2009, 462, 272–274. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.; de Waard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. B 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, R.W.; Crawford, A.J.; Bauer, A.M.; Che, J.; Donnellan, S.C.; Fritz, U.; Haddad, C.F.B.; Nagy, Z.T.; Poyarkov, N.A.; Vences, M.; et al. Cold Code: The global initiative to DNA barcode amphibians and nonavian reptiles. Mol. Ecol. Resour. 2013, 13, 161–167. [Google Scholar] [CrossRef]
- Perl, R.G.B.; Nagy, Z.T.; Sonet, G.; Glaw, F.; Wollenberg, K.C.; Vences, M. DNA barcoding Madagascar’s amphibian fauna. Amphib.-Reptil. 2014, 35, 197–206. [Google Scholar] [CrossRef]
- Shen, Y.Y.; Chen, X.; Murphy, R.W. Assessing DNA barcoding as a tool for species identification and data quality control. PLoS ONE 2013, 8, e57125. [Google Scholar] [CrossRef] [PubMed]
- Meiri, S.; Mace, G.M. New taxonomy and the origin of species. PLoS Biol. 2007, 5, e194. [Google Scholar] [CrossRef]
- Hajibabaei, M.; Singer, G.A.C.; Hebert, P.D.N.; Hickey, D.A. DNA barcoding: How it complements taxonomy, molecular phylogenetics and population genetics. Trends Genet. 2007, 23, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Spichiger, R.; Palese, R.; Chautems, A.; Ramella, L. Origin, affinities and diversity hot spots of the Paraguayan dendrofloras. Candollea 1995, 50, 515–537. [Google Scholar]
- Morrone, J.J. Biogeografía de América Latina y el Caribe; Manuales & Tesis; SEA/UNESCO/CYTED: Zaragoza, España, 2001. [Google Scholar]
- Cacciali, P.; Scott, N. Revisión del Género Ophiodes de Paraguay (Squamata: Anguidae). Bol. Soc. Zool. Urug. 2012, 21, 1–8. [Google Scholar]
- Cabral, H.; Netto, F. Epictia vellardi. Geographic distribution. Herp. Rev. 2016, 47, 83. [Google Scholar]
- Cacciali, P.; Cabral, H. The genus Chironius (Serpentes, Colubridae) in Paraguay: Composition, distribution, and morphology. Bas. App. Herp. 2015, 29, 51–60. [Google Scholar] [CrossRef]
- Cacciali, P.; Smith, P.; Källberg, A.; Pheasey, H.; Atkinson, K. Reptilia, Squamata, Serpentes, Lygophis paucidens Hoge, 1952: First records for Paraguay. Check List 2013, 9, 131–132. [Google Scholar] [CrossRef]
- Smith, P.; Cacciali, P.; Scott, N.; del Castillo, H.; Pheasey, H.; Atkinson, K. First record of the globally-threatened Cerrado endemic snake Philodryas livida (Amaral, 1923) (Serpentes, Dipsadidae) from Paraguay, and the importance of the Reserva Natural Laguna Blanca to its conservation. Cuad. Herpetol. 2014, 28, 169–171. [Google Scholar]
- Cacciali, P.; Espínola, D.; Centrón Viñales, S.; Gauto Espínola, I.; Cabral, H. Squamata, Serpentes, Micrurus silviae Di-Bernardo, Borges-Martins and Silva, 2007: Presence confirmation in Paraguay. Check List 2011, 7, 809–810. [Google Scholar] [CrossRef]
- Carvalho, A.L.G. Three new species of the Tropidurus spinulosus group (Squamata: Tropiduridae) from Eastern Paraguay. Am. Mus. Nov. 2016, 3853, 1–44. [Google Scholar] [CrossRef]
- Cacciali, P.; Scott, N. Key to the Ophiodes (Squamata: Sauria: Diploglossidae) of Paraguay with the description of a new species. Zootaxa 2015, 3980, 42–50. [Google Scholar] [CrossRef]
- Cabral, H.; Cacciali, P. A new species of Phalotris (Serpentes: Dipsadidae) from the Paraguayan Chaco. Herpetologica 2015, 71, 72–77. [Google Scholar] [CrossRef]
- Gamble, T.; Colli, G.R.; Rodrigues, M.T.; Werneck, F.P.; Simons, A.M. Phylogeny and cryptic diversity in geckos (Phyllopezus; Phyllodactylidae; Gekkota) from South America’s open biomes. Mol. Phyl. Evol. 2012, 62, 943–953. [Google Scholar] [CrossRef]
- Werneck, F.P.; Gamble, T.; Colli, G.R.; Rodrigues, M.T.; Sites, J. Deep diversification and long-term persistence in the South American “Dry Diagonal”: Integrating continent-wide phylogeography and distribution modeling of geckos. Evolution 2012, 66, 3014–3034. [Google Scholar] [CrossRef]
- Morando, M.; Medina, C.D.; Ávila, L.J.; Pérez, C.H.F.; Buxton, A.; Sites, J.W. Molecular phylogeny of the New World gecko genus Homonota (Squamata: Phyllodactylidae). Zool. Scr. 2014, 43, 249–260. [Google Scholar] [CrossRef]
- Recoder, R.S.; Werneck, F.; Teixeira, M.; Colli, G.R.; Sites, J.W.; Rodrigues, M.T. Geographic variation and systematic review of the lizard genus Vanzosaura (Squamata, Gymnophthalmidae), with the description of a new species. Zool. J. Lin. Soc. 2014, 171, 206–225. [Google Scholar] [CrossRef]
- Sacchi, C.T.; Whitney, A.M.; Mayer, L.W.; Morey, R.; Steigerwalt, A.; Boras, A.; Weyant, R.S.; Popovic, T. Sequencing of 16S rRNA gene: A rapid tool for identification of Bacillus anthracis. Emerg. Infect. Dis. 2002, 8, 1117–1123. [Google Scholar] [CrossRef]
- Jansen, M.; Schultze, A. Molecular, morphology and bioacoustic data suggest Bolivian distribution of a large species of the Leptodactylus pentadactylus group (Amphibia: Anura: Leptodactylidae). Zootaxa 2012, 3307, 35–47. [Google Scholar] [CrossRef]
- Scherz, M.D.; Vences, M.; Borrell, J.; Ball, L.; Nomenjanahary, D.H.; Parker, D.; Rakotondratsima, M.; Razafimandimby, E.; Starnes, T.; Rabearivony, J.; et al. A new frog species of the subgenus Asperomantis (Anura, Mantellidae, Gephyromantis) from the Bealanana District of northern Madagascar. Zoosyst. Evol. 2017, 93, 451–466. [Google Scholar] [CrossRef]
- Batista, A.; Hertz, A.; Köhler, G.; Mebert, K.; Veselý, M. Morphological variation and phylogeography of frogs related to Pristimantis caryophyllaceus (Anura: Terra-rana: Craugastoridae) in Panama. Salamandra 2014, 50, 155–171. [Google Scholar]
- Köhler, G.; Townsend, J.H.; Petersen, C.B. A taxonomic revision of the Norops tropidonotus complex (Squamata, Dactyloidae), with the resurrection of N. spilorhipis (Álvarez del Toro and Smith, 1956) and the description of two new species. Mesoam. Herpetol. 2016, 3, 8–41. [Google Scholar]
- Cacciali, P.; Martínez, N.; Köhler, G. Revision of the phylogeny and chorology of the tribe Iphisini with the revalidation of Colobosaura kraepelini Werner, 1910 (Reptilia, Squamata, Gymnophthalmidae). ZooKeys 2017, 669, 89–105. [Google Scholar] [CrossRef]
- Cacciali, P.; Morando, M.; Medina, C.D.; Köhler, G.; Motte, M.; Avila, L.J. Taxonomic analysis of Paraguayan samples of Homonota fasciata Duméril & Bibron (1836) with the revalidation of Homonota horrida Burmeister (1861) (Reptilia: Squamata: Phyllodactylidae) and the description of a new species. PeerJ 2017, 5, e3523. [Google Scholar] [CrossRef]
- Cacciali, P.; Köhler, G. Diversity of Tropidurus (Squamata: Tropiduridae) in Paraguay—An integrative taxonomic approach based on morphological and molecular genetic evidence. Zootaxa 2018, 4375, 511–536. [Google Scholar] [CrossRef]
- Cacciali, P.; Morando, M.; Avila, L.J.; Köhler, G. Description of a new species of Homonota (Reptilia, Squamata, Phyllodactylidae) from the central region of northern Paraguay. Zoosyst. Evol. 2018, 94, 147–161. [Google Scholar] [CrossRef]
- Cacciali, P.; Lotzkat, S.; Gamble, T.; Köhler, G. Cryptic diversity in the Neotropical gecko genus Phyllopezus Peters, 1878 (Reptilia: Squamata: Phyllodactylidae): A new species from Paraguay. Int. J. Zool 2018. [Google Scholar] [CrossRef]
- Cacciali, P. Colecta y Preparación de Anfibios y Reptiles: Manual para Colecta Científica; Editorial Académica Española: Saarbrücken, Germany, 2013. [Google Scholar]
- Scrocchi, G.; Kretzschmar, S. Guía de Métodos de Captura y Preparación de Anfibios y Reptiles para Estudios Científicos y Manejo de Colecciones Herpetológicas; Miscelánea N° 102; Fundación Miguel Lillo: San Miguel de Tucumán, Argentina, 1996. [Google Scholar]
- Simmons, J.E. Herpetological Collecting and Collections Management; Society for the Study of Amphibians and Reptiles: Salt Lake, UT, USA, 2002. [Google Scholar]
- Cacciali, P.; Scott, N.; Aquino, A.L.; Fitzgerald, L.A.; Smith, P. The Reptiles of Paraguay: Literature, distribution, and an annotated taxonomic checklist. Spec. Publ. Mus. Southwest. Biol. 2016, 11, 1–373. [Google Scholar]
- Ivanova, N.V.; Dewaard, J.R.; Hebert, P.D. An inexpensive, automation-friendly protocol for recovering high-quality DNA. Mol. Ecol. Notes 2006, 6, 998–1002. [Google Scholar] [CrossRef]
- Lotzkat, S.; Hertz, A.; Bienentreu, J.F.; Köhler, G. Distribution and variation of the giant alpha anoles (Squamata: Dactyloidae) of the genus Dactyloa in the highlands of western Panama, with the description of a new species formerly referred to as D. microtus. Zootaxa 2013, 3626, 1–54. [Google Scholar] [CrossRef]
- Vilgalys, R. Taxonomic misidentification in public DNA databases. New Phytol. 2003, 160, 4–5. [Google Scholar] [CrossRef]
- Zhang, G. Specimens versus sequences. Science 2009, 323, 1672. [Google Scholar] [CrossRef]
- Park, K.S.; Ki, C.S.; Kang, C.I.; Kim, Y.J.; Chung, D.R.; Peck, K.R.; Song, J.H.; Lee, N.Y. Evaluation of the GenBank, EzTaxon, and BIBI services for molecular identification of clinical blood culture isolates that were unidentifiable or misidentified by conventional methods. J. Clin. Microbiol. 2012, 50, 1792–1795. [Google Scholar] [CrossRef]
- Pellegrino, K.C.M.; Rodrigues, M.T.; Yonenaga-Yassuda, Y.; Sites, J.W. A molecular perspective on the evolution of microteiid lizards (Squamata, Gymnophthalmidae), and a new classification for the family. Biol. J. Lin. Soc. 2001, 74, 315–338. [Google Scholar] [CrossRef] [Green Version]
- Stucky, B.J. SeqTrace: A graphical tool for rapidly processing DNA sequencing chromatograms. J. Biomol. Tech. 2012, 23, 90–93. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2017, bbx108. [Google Scholar] [CrossRef]
- Katoh, K.; Toh, H. Recent developments in the MAFFT multiple sequence alignment program. Brief. Bioinform. 2008, 9, 286–298. [Google Scholar] [CrossRef] [Green Version]
- Yachdav, G.; Wilzbach, S.; Rauscher, B.; Sheridan, R.; Sillitoe, I.; Procter, J.; Lewis, S.E.; Rost, B.; Goldberg, T. MSAViewer: Interactive JavaScript visualization of multiple sequence alignments. Bioinformatics 2016, 32, 3501–3503. [Google Scholar] [CrossRef]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2016, 34, 772–773. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Schwarz, G.E. Estimating the dimension of a model. Ann. Stat. 1978, 6, 461–464. [Google Scholar] [CrossRef]
- Brewer, M.J.; Butler, A.; Cooksley, S. The relative performance of AIC, AICc and BIC in the presence of unobserved heterogeneity. Met. Ecol. Evol. 2016, 7, 679–692. [Google Scholar] [CrossRef]
- Yang, Z.; Landry, J.F.; Hebert, P.D.N. A DNA Barcode Library for North American Pyraustinae (Lepidoptera: Pyraloidea: Crambidae). PLoS ONE 2016, 11, e0161449. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating Maximum Likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for Maximum Likelihood analysis. Nucleic Acid Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef]
- Anisimova, M.; Gil, M.; Dufayard, J.F.; Dessimoz, C.; Gascuel, O. Survey of branch support methods demonstrates accuracy, power, and robustness of fast Likelihood-based approximation schemes. Syst. Biol. 2011, 60, 685–699. [Google Scholar] [CrossRef]
- Minh, B.Q.; Thi Nguyen, M.A.; von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Miller, H.C. Cloacal and buccal swabs are a reliable source of DNA for microsatellite genotyping of reptiles. Conserv. Genet. 2006, 7, 1001–1003. [Google Scholar] [CrossRef]
- Pyron, R.A.; Burbrink, F.T.; Wiens, J.J. A phylogeny and revised classification of Squamata, including 4161 species of lizards and snakes. BMC Evol. Biol. 2013, 13, 93. [Google Scholar] [CrossRef]
- Bogaardt, C.; Carvalho, L.; Hill, V.; O’Toole, A.; Rambaut, A. FigTree, Version 1.4.3, Molecular Evolution, Phylogenetics and Epidemiology, United Kingdom. 2018. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 14 November 2018).
- Madison, W.P.; Madison, D.R. Mesquite: A Modular System for Evolutionary Analysis, Version 3.2, Mesquite Project, United States. 2017. Available online: http://mesquiteproject.org (accessed on 15 June 2018).
- Chapple, D.G.; Ritchie, P.A. A retrospective approach to testing DNA barcoding method. PLoS ONE 2013, 8, e77882. [Google Scholar] [CrossRef]
- Padial, J.M.; Miralles, A.; De la Riva, I.; Vences, M. The integrative future of taxonomy. Front. Zool. 2010, 7, 16. [Google Scholar] [CrossRef]
- Mott, T.; Vieites, D.R. Molecular phylogenetics reveals extreme morphological homoplasy in Brazilian worm lizards challenging current taxonomy. Mol. Phylogenet. Evol. 2009, 51, 190–200. [Google Scholar] [CrossRef]
- Grazziotin, F.G.; Zaher, H.; Murphy, R.W.; Scrocchi, G.; Benavides, M.A.; Zhang, Y.P.; Bonatto, S.L. Molecular phylogeny of the New World Dipsadidae (Serpentes: Colubroidea): A reappraisal. Cladistics 2012, 28, 437–459. [Google Scholar] [CrossRef]
- Da Silva, N.J.; Sites, J.W. Revision of the Micrurus frontalis complex (Serpentes: Elapidae). Herpetol. Monogr. 1999, 13, 142–194. [Google Scholar] [CrossRef]
- Cartes, J.L. Brief history of conservation in the Interior Atlantic Forest. In The Atlantic Forest of South America; Galindo-Leal, C., Gusmão Câmara, I., Eds.; Island Press: London, UK, 2003; pp. 269–287. [Google Scholar]
- Huang, C.; Kim, S.; Song, K.; Townshend, J.; Davis, P.; Altstatt, A.; Rodas, O.; Yanosky, A.A.; Clay, R.; Tucker, C.J.; et al. Assessment of Paraguay’s forest cover change using Landsat observations. Glob. Planet. Chang. 2009, 67, 1–12. [Google Scholar] [CrossRef]
- Yanosky, A.A. Paraguay’s challenge of conserving natural habitats and biodiversity with global markets demanding for products. In Conservation Biology: Voices from the Tropics; Sodhi, N.S., Gibson, L., Raven, P.H., Eds.; Wiley-Blackwell: Oxford, UK, 2013; pp. 113–119. [Google Scholar]
- Cacciali, P.; Cabral, H.; Yanosky, A.A. Conservation implications of protected areas’ coverage for Paraguay’s reptiles. Parks 2015, 21, 101–119. [Google Scholar] [CrossRef]
- Motte, M.; Núñez, K.; Cacciali, P.; Brusquetti, F.; Scott, N.; Aquino, A.L. Categorización del estado de conservación de los anfibios y reptiles de Paraguay. Cuad. Herpetol. 2009, 23, 5–18. [Google Scholar]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cacciali, P.; Buongermini, E.; Köhler, G. Barcoding Analysis of Paraguayan Squamata. Diversity 2019, 11, 152. https://doi.org/10.3390/d11090152
Cacciali P, Buongermini E, Köhler G. Barcoding Analysis of Paraguayan Squamata. Diversity. 2019; 11(9):152. https://doi.org/10.3390/d11090152
Chicago/Turabian StyleCacciali, Pier, Emilio Buongermini, and Gunther Köhler. 2019. "Barcoding Analysis of Paraguayan Squamata" Diversity 11, no. 9: 152. https://doi.org/10.3390/d11090152
APA StyleCacciali, P., Buongermini, E., & Köhler, G. (2019). Barcoding Analysis of Paraguayan Squamata. Diversity, 11(9), 152. https://doi.org/10.3390/d11090152