Inventory and Historical Changes in the Marine Flora of Tomioka Peninsula (Amakusa Island), Japan

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Research Area
2.2. Study Sites
2.3. Sampling Timing and Strategy
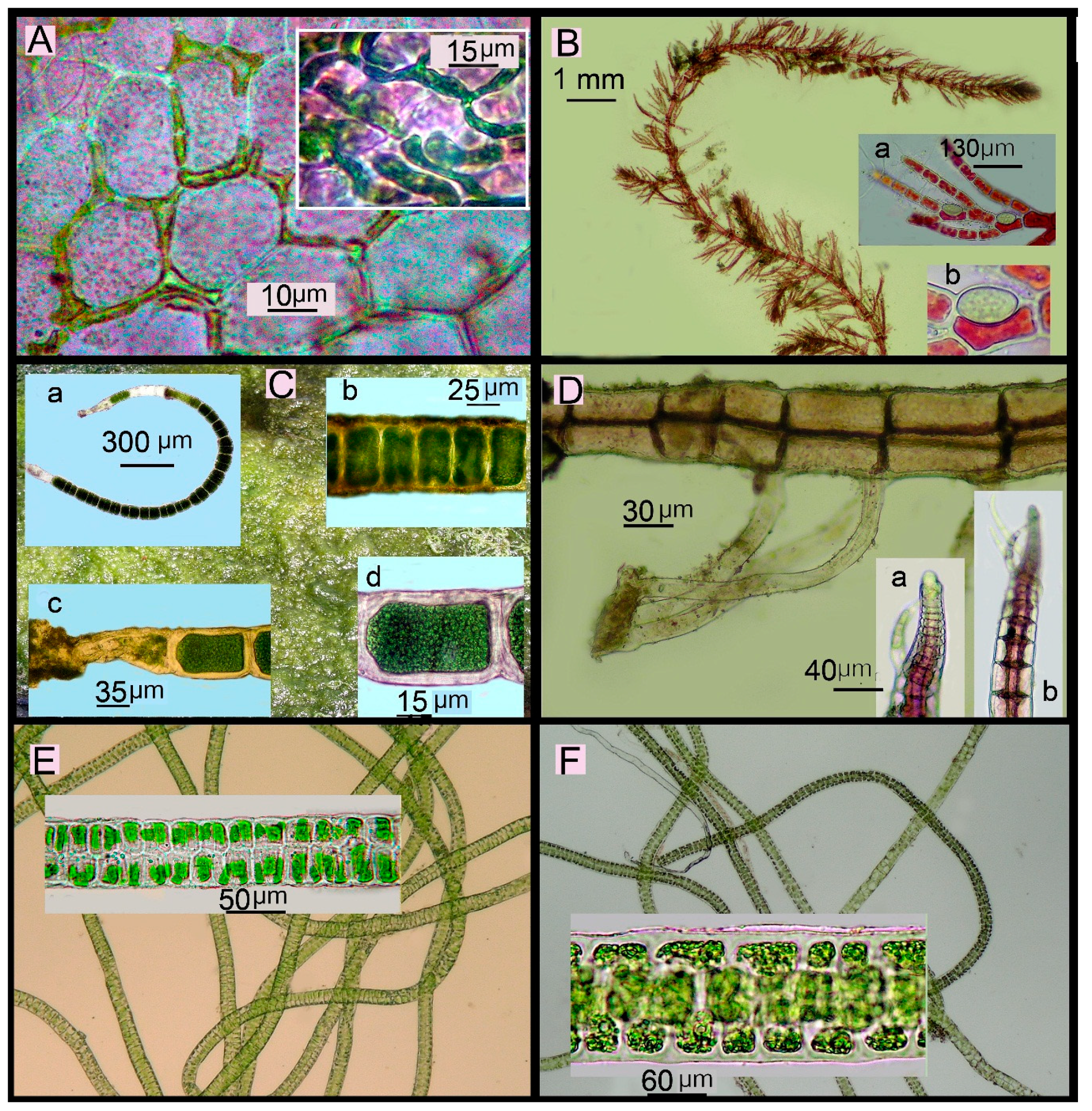
2.4. Collection, Identification and Conservation of Marine Plants
- (1)
- In each tidal zone, algal turf communities (with thalli less than 5 cm in height), crustose coralline algae (CCA) and large upright-growing algae (with thalli more than 5 cm in height) in an area of ~100 m2 were visually identified. The selected communities were photographed many times at a right angle. In the communities of algal turf and CCA, samples were collected from three randomly selected square plots, each plot measuring ~100 cm2. In communities of upright-growing algae, samples were also collected from three squares, each measuring from 0.5 m2 to 1 m2. Algae were also separately collected from areas outside the selected square plots.
- (2)
- Algae collected from different communities were stored in separate plastic bags and then placed in the refrigerator prior to further processing. Freshly collected material was identified using monographic publications, floristic studies, and systematic articles described by Titlyanova et al. [10]. The systematics and nomenclature followed Guiry & Guiry [11]. The previously known and newly recorded species for Amakusa-Shimoshima Island were verified using the article by Segawa & Yoshida [3] and for Japan by AlgaeBase [11]. Hierarchical classification of the Phylum Rhodophyta (Rh) was carried out as Saunders & Hommersand [12]. The classification system of the Phyla Chlorophyta (Ch) and Ochrophyta followed Tsuda [13].
- (3)
- The collections of both macrophytes and their epiphytes were preserved as dried herbarial specimens and deposited in the herbarium at the A.V. Zhirmunsky Institute of Marine Biology, National Scientific Center of Marine Biology, Far Eastern Branch, Russian Academy of Science, Vladivostok 690041, Russian Federation.
2.5. Terms of the Comparative Analysis, when Studying Historical Changes in the Flora
3. Results
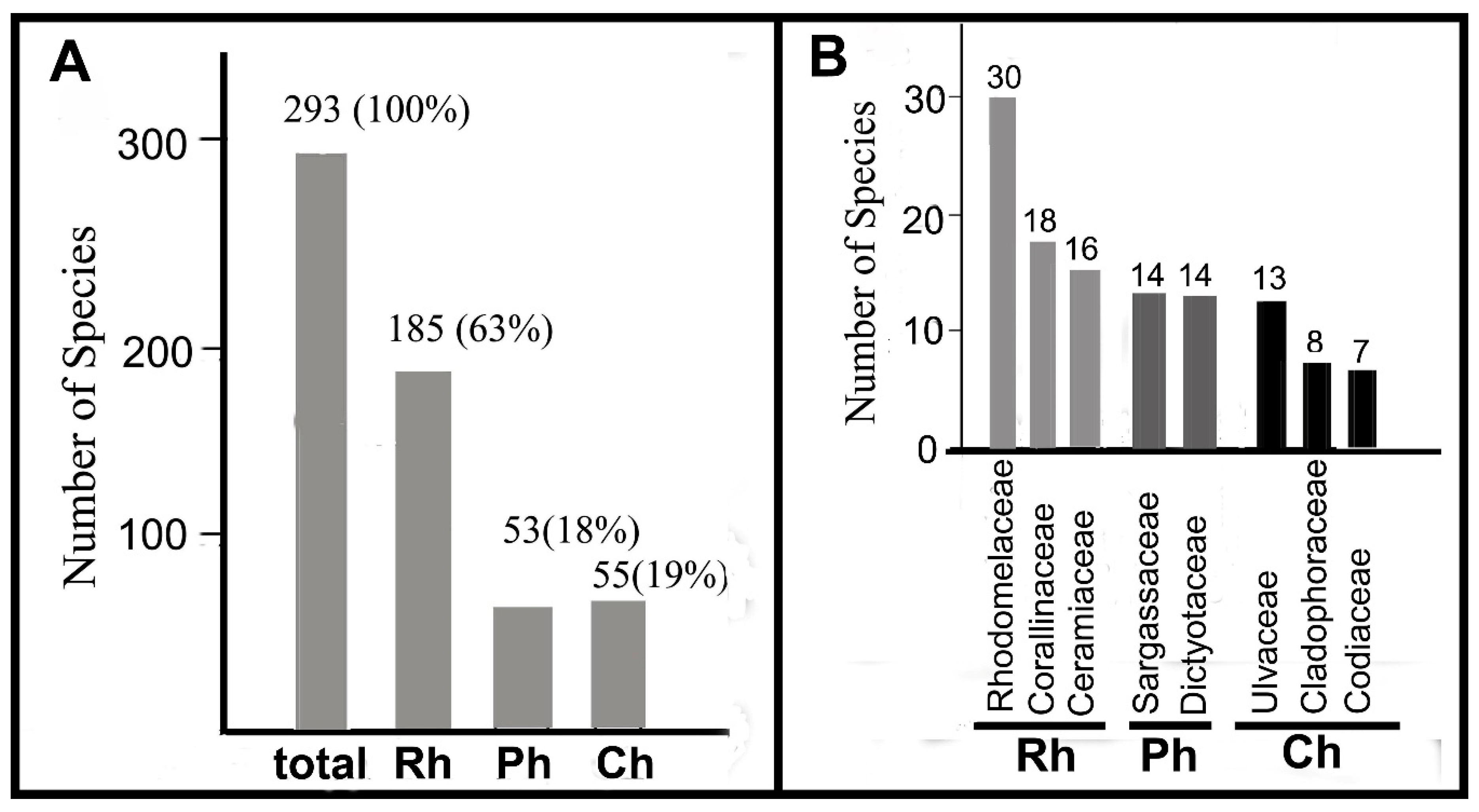
3.1. Species and Life Form Diversity
3.2. Newly Recorded Species for Amakusa-Shimoshima Island
3.3. Newly Recorded Species for Japan
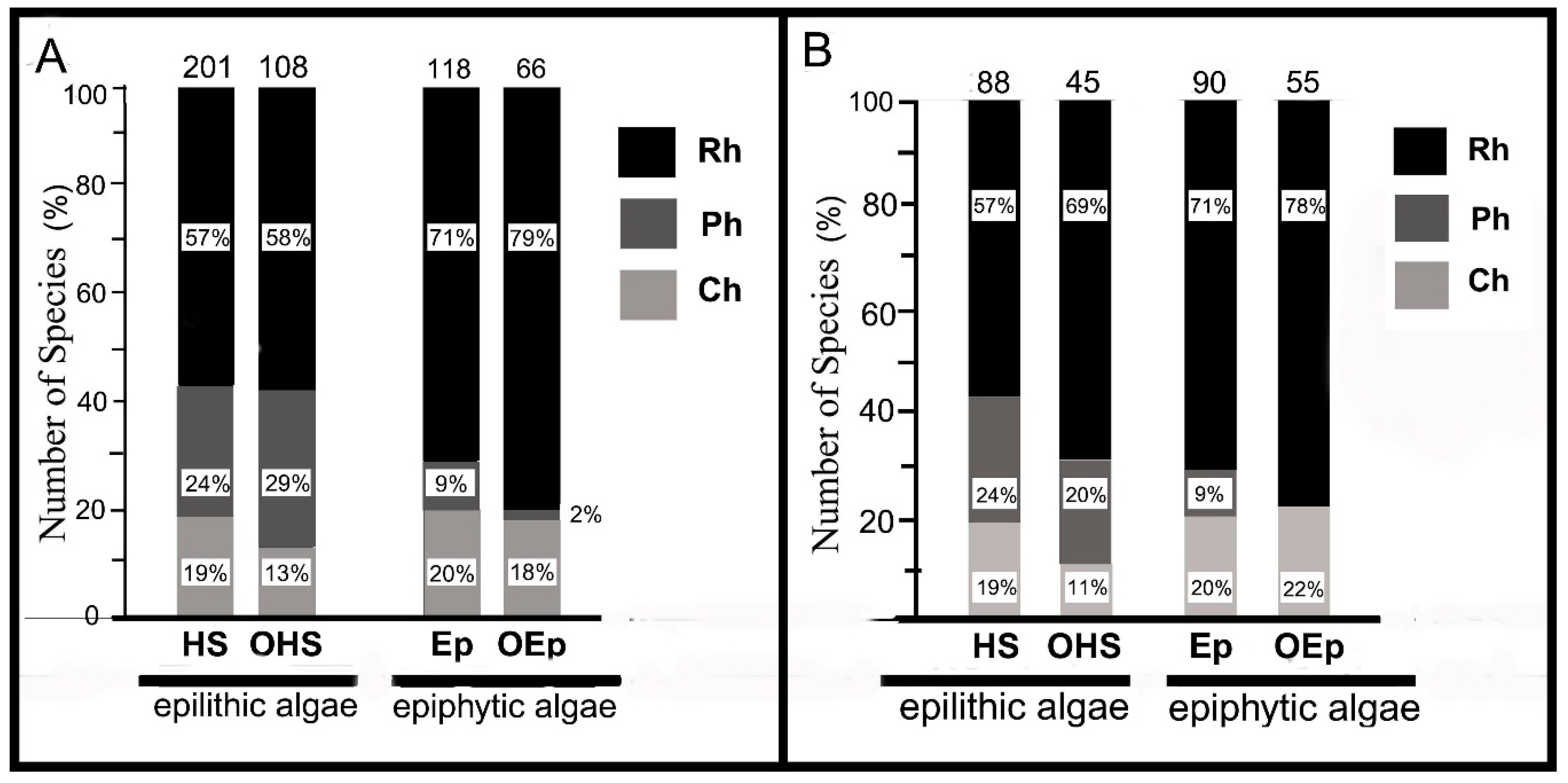
3.4. Distribution of Algal Species and their Communities in Tidal Zones
4. Discussion
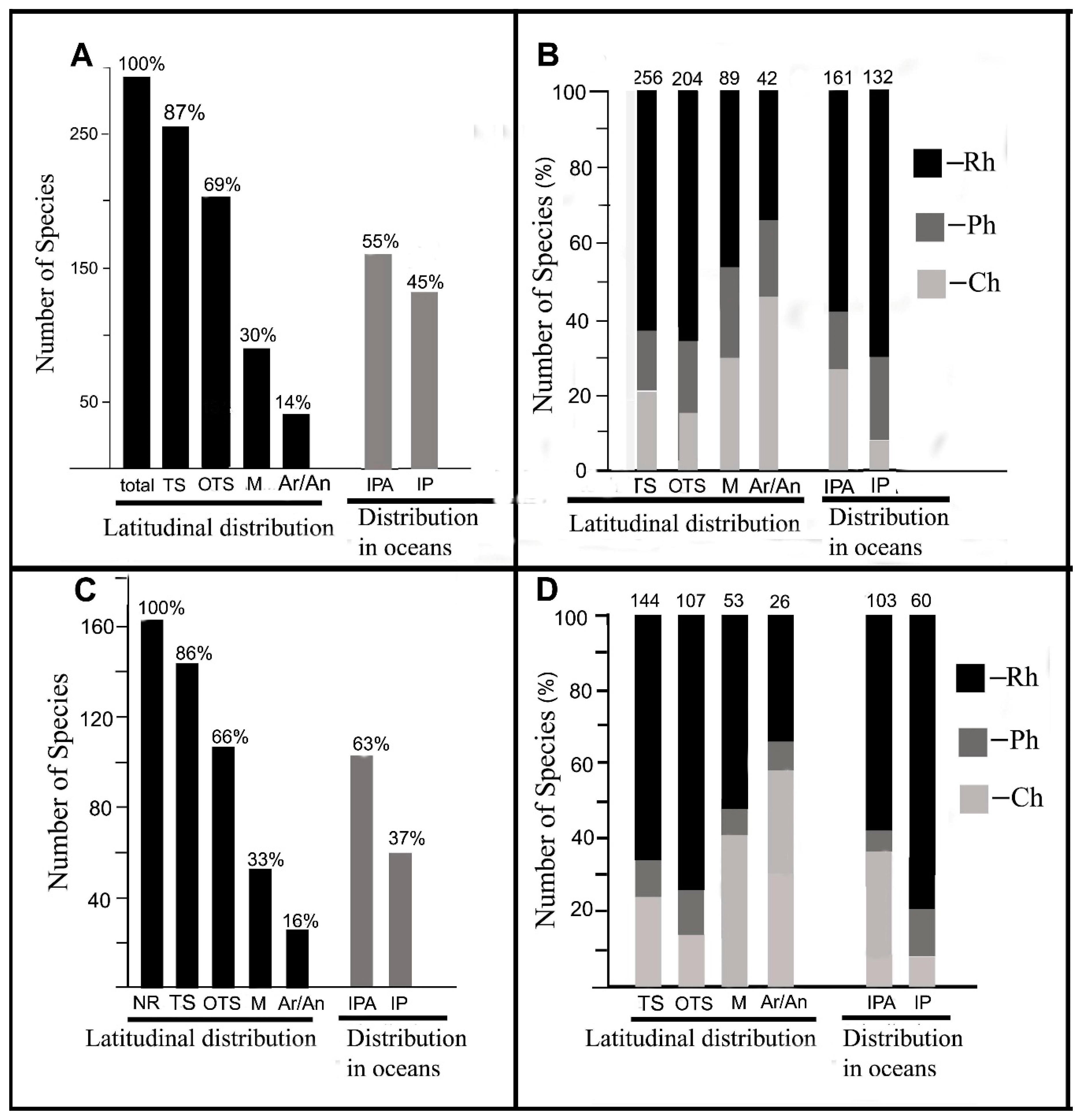
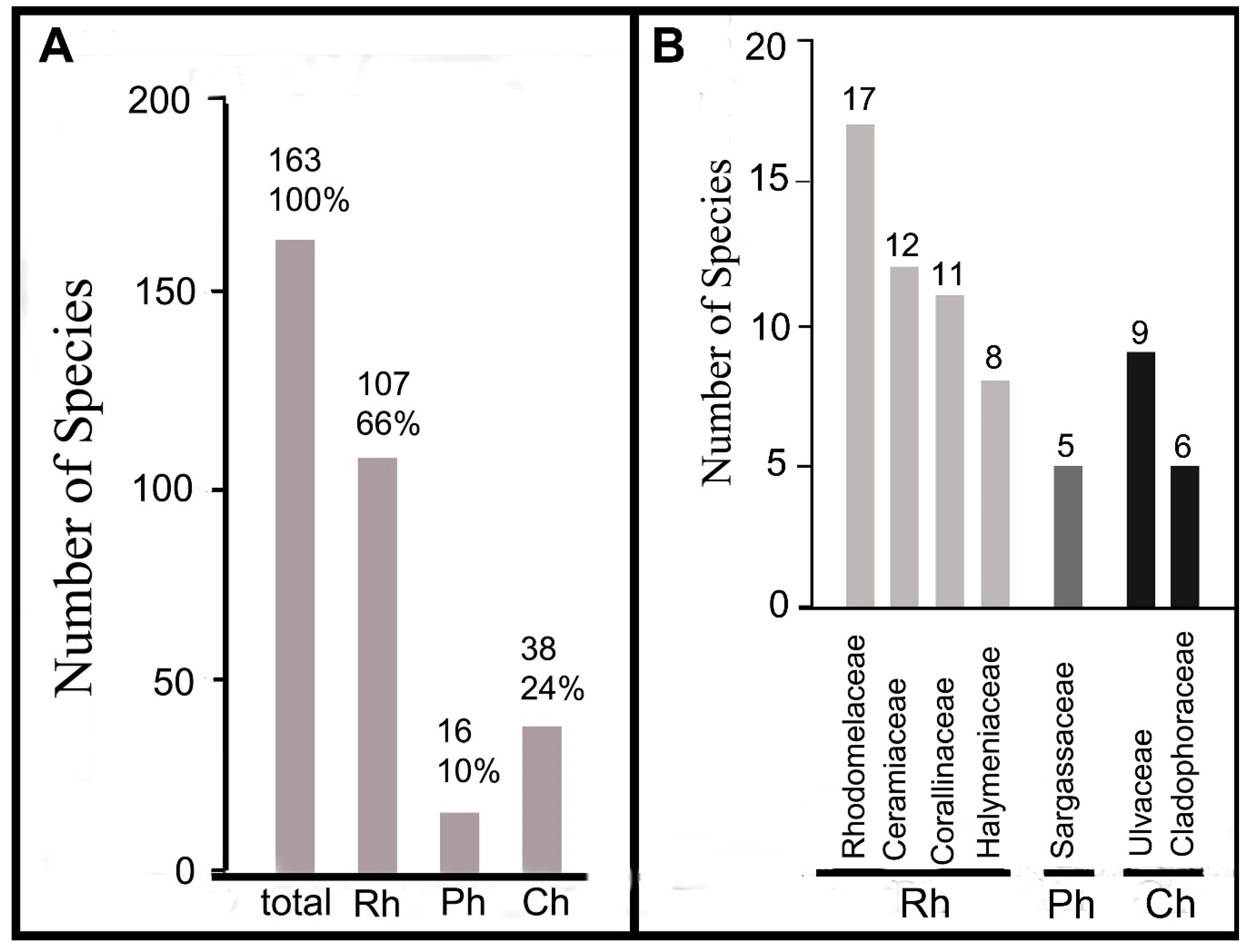
4.1. Recent Seaweed Flora
4.2. Historical Changes in the Marine Flora
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Climate-Japan. Available online: https://www.climatestotravel.com/climate/japan (accessed on 29 August 2019).
- Japan Sea temperatures. Available online: https://www.seatemperature.org/asia/japan/ (accessed on 29 August 2019).
- Segawa, S.; Yoshida, T. Decapoda crustacean. In Fauna and Flora of the Sea around the Amakusa Marine Biological Laboratory, Part II; Tomioka, R., Ed.; Kumamoto Prefecture: Amakusa, Japan, 1961; p. 24. [Google Scholar]
- Nandakumar, K.; Tanaka, M.; Kikuchi, T. Interspecific competition among fouling organisms in Tomioka Bay, Japan. Mar. Ecol. 1993, 94, 43–50. [Google Scholar] [CrossRef]
- Tioho, H.; Tokeshi, M.; Nojima, S. Experimental analysis of recruitment in a scleractinian coral at high latitude. Mar. Ecol. 2001, 213, 79–86. [Google Scholar] [CrossRef]
- Nishihara, G.N.; Noro, T.; Terada, R. Effect of temperature and light on the photosynthetic performance of two edible seaweeds: Meristotheca coacta Okamura and Meristotheca papulosa J. Agardh (Solieriaceae, Rhodophyta). Aquac. Sci. 2012, 60, 377–388. [Google Scholar]
- Terada, R.; Tanaka, T.; Uchimura, M. Morphology and distribution of Caulerpa lentillifera J. Agardh (Chrolophyceae) in Japanese waters, including the first record from Southern Kyushu and Northern Ryukyu Islands. J. Jpn. Bot. 2012, 60, 377–388. [Google Scholar]
- Nakano, Y. Global environmental change and coral bleaching. Coral Reefs Jpn. 2004, 42–48. [Google Scholar]
- Perestenko, L.P. Red Algae of the Far-Eastern Seas of Russia; Olga Publishing House: St Petersburg, Russia, 1994; p. 330. [Google Scholar]
- Titlyanova, T.V.; Titlyanov, E.A.; Kalita, T.L. Marine algal flora of Hainan Island: A comprehensive synthesis. Coast. Ecosyst. 2014, 1, 28–53. [Google Scholar]
- AlgaeBase. World-Wide Electronic Publication. Available online: http://www.algaebase.org (accessed on 29 August 2019).
- Saunders, G.W.; Hommersand, M.H. Assessing red algal supraordinal diversity and taxonomy in the context of contemporary systematic data. Am. J. Bot. 2004, 91, 1494–1507. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, R.T. Checklist and Bibliography of the Marine Benthic Algae from the Mariana Islands (Guam and CNMI); University of Guam: Guam, USA, 2003; Volume 107, pp. 1–37. [Google Scholar]
- Titlyanov, E.A.; Titlyanova, T.V.; Belous, O.S. Checklist of the marine flora of Nha Trang Bay (Vietnam, South China Sea) and decadal changes in the species diversity composition between 1953 and 2010. Bot. Mar. 2015, 58, 367–377. [Google Scholar] [CrossRef]
- Vinogradova, K.L. Green algae. In Algae of the Far-Eastern Seas of USSR; Perestenko, L.P., Ed.; Nauka: St. Petersburg, Russia, 1975; p. 147. [Google Scholar]
- Titlyanova, T.V.; Titlyanov, E.A.; Xia, B.; Bartsch, I. New records of benthic marine green algae (Chlorophyta) for the island of Hainan (China). Nova Hedwigia 2012, 94, 441–470. [Google Scholar] [CrossRef]
- Littler, D.M.; Littler, M.M. Caribbean Reef Plants; OffShore Graphics: Washington, DC, USA, 2000; p. 542. [Google Scholar]
- Titlyanova, T.V.; Titlyanov, E.A.; Li, X.B. The significance of new records of benthic red algae (Rhodophyta) for Hainan Island (and China) between 1990 and 2016. Diversity 2017, 9, 24. [Google Scholar] [CrossRef]
- Briggs, J.C. Marine Zoogeography; McGraw-Hill: New York, NY, USA, 1974; p. 475. [Google Scholar]
- Santelices, B.; Bolton, J.J.; Meneses, I. Chapter Six Marine Algal Communities. In Marine Macroecology; Whitman, J.D., Roy, K., Eds.; Chicago University Press: Chicago, IL, USA, 2009; pp. 153–192. [Google Scholar]
- Lüning, K. Seaweeds. Their Environment, Biogeography and Ecophysiology; John Wiley & Sons: Hoboken, NJ, USA, 1990; p. 527. [Google Scholar]
- Pham, H.H. Marine Algae of South Vietnam; Trung Tam Hoc Lieu Saigon: Saigon, Vietnam, 1969; p. 558. [Google Scholar]
- Lewis, J.E.; Norris, J.N. A history and annotated account of the benthic marine algae of Taiwan. Smithson. Contrib. Mar. Sci. 1987, 29, 1–38. [Google Scholar] [CrossRef]
- Perestenko, L.P. Algae of the Peter Great Bay (the Sea of Japan); Nauka: St. Petersburg, Russia, 1980; p. 232. [Google Scholar]
- Nguyen, T.V.; Le, N.H.; Lin, S.M.; Steen, F.; de Clerck, O. Checklist of the marine macroalgae of Vietnam. Bot. Mar. 2013, 56, 207–227. [Google Scholar] [CrossRef]
- Belous, O.S.; Titlyanova, T.V.; Titlyanov, E.A. Marine Plants of the Trinity Bay and Adjacent Water Areas; Dalnauka: Vladivostok, Russia, 2013; p. 264. [Google Scholar]
- Titlyanov, E.A.; Titlyanova, T.V.; Li, X.B.; Kalita, T.L.; Huang, H. Recent (2008–2012) seaweed flora of Hainan Island, South China Sea. Mar. Biol. Res. 2015, 11, 540–550. [Google Scholar] [CrossRef]
- Hayashida, F. Synecological studies of a brown alga, Ecklonia cava Kjellmann, forming aquatic forest. I. Benthic marine algal vegetation at survey area. J. Mar. Sci. Tech-Japan. 1983, 16, 207–212. [Google Scholar]
- Hayashida, F. Synecological studies of a brown alga, Ecklonia cava Kjellmann, forming aquatic forest. II. On growth of Ecklonia cava. J. Mar. Sci. Tech-Jpn. 1984, 18, 275–280. [Google Scholar]
- Maegawa, M.; Yokohama, Y.; Aruga, Y. Critical light conditions for young Ecklonia cava and Eisenia bicyclis with reference to photosynthesis. Hydrobiologia 1987, 447–455. [Google Scholar] [CrossRef]
- Titlyanov, E.A.; Titlyanova, T.V.; Belous, O.S.; Kalita, T.L. Inventory change (1990s–2010s) in the marine flora of Sanya Bay (Hainan Island, China). J. Mar. Biol. Assoc. UK 2015, 95, 461–470. [Google Scholar] [CrossRef]
- Titlyanov, E.A.; Titlyanova, T.V.; Kalita, T.L.; Tokeshi, M. Decadal changes in the algal assemblages of tropical-subtropical Yonaguni Island in the western Pacific. Coast. Ecosyst. 2016, 3, 16–37. [Google Scholar]
- Haraguchi, H.; Sekida, S. Recent changes in the distribution of Sargassum species in Kochi, Japan. Kuroshio Sci. 2008, 2, 41–46. [Google Scholar]
- Nagai, S.; Yoshida, G.; Tarutani, K. Change in species composition and distribution of algae in the coastal waters of western Japan. In Global Warming Impacts: Case Studies on the Economy, Human Health, and on Urban and Natural Environments; Casalegno, S., Ed.; BoD–Books on Demand: Norderstedt, Germany, 2011; pp. 2–31. [Google Scholar]
- Fujita, D. Current status and problems of isoyake in Japan. Bull Fish Res. Agen 2010, 32, 33–42. [Google Scholar]
- Hiraoka, M.; Ura, Y.; Haraguchi, H. Relationship between seaweed beds and seawater temperature in the Tosa Bay. Aquabiology 2005, 27, 485–493. [Google Scholar]
- Aratake, H.; Shimizu, H.; Watanabe, K.; Yoshida, G. Long-term change in Sargassum-bed distribution along the coast of Kushima-city, southern part of Miyazaki Prefecture, Japan. Bull. Miyazaki Prefectural Fish. Exp. Stn. 2007, 11, 1–13. [Google Scholar]
- Lima, F.P.; Ribeiro, P.A.; Queiroz, N.; Hawkins, S.J.; Santos, A.M. Do distributional shifts of northern and southern species of algae match the warming pattern? Glob. Chang. Biol. 2007, 13, 2592–2604. [Google Scholar] [CrossRef]
- Hawkins, S.J.; Moore, P.J.; Burrows, M.T.; Poloczanska, E.; Mieszkowska, N.; Herbert, R.J.H.; Jenkins, S.R.; Thompson, R.C.; Genner, M.J.; Southward, A.J. Complex interactions in a rapidly changing world: Responses of rocky shore communities to recent climate change. Clim. Res. 2008, 37, 123–133. [Google Scholar] [CrossRef]
- Schiel, D.; Steinbeck, J.R.; Foster, M.S. Ten years of induced ocean warming causes comprehensive. Ecology 2004, 85, 1833–1839. [Google Scholar] [CrossRef]
- Araujo, R.M.; Assis, J.; Aquillar, R.; Airoldi, L.; Barbara, I.; Bartsch, I.; Bekkby, T.; Christie, H.; Davoult, D.; Derrien-Courtel, S.; et al. Status, trends and drivers of kelp forests in Europe: An expert assessment. Biodivers. Conserv. 2016, 25, 1319–1348. [Google Scholar] [CrossRef]
- Harley, C.D.G.; Anderson, K.M.; Demes, K.W.; Jorve, J.P.; Kordas, R.L.; Coyle, T.A. Effects of climate change on global seaweed communities. J. Phycol. 2012, 48, 1064–1078. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Titlyanov, E.A.; Titlyanova, T.V.; Tokeshi, M.; Li, X. Inventory and Historical Changes in the Marine Flora of Tomioka Peninsula (Amakusa Island), Japan. Diversity 2019, 11, 158. https://doi.org/10.3390/d11090158
Titlyanov EA, Titlyanova TV, Tokeshi M, Li X. Inventory and Historical Changes in the Marine Flora of Tomioka Peninsula (Amakusa Island), Japan. Diversity. 2019; 11(9):158. https://doi.org/10.3390/d11090158
Chicago/Turabian StyleTitlyanov, Eduard A., Tamara V. Titlyanova, Mutsunori Tokeshi, and Xiubao Li. 2019. "Inventory and Historical Changes in the Marine Flora of Tomioka Peninsula (Amakusa Island), Japan" Diversity 11, no. 9: 158. https://doi.org/10.3390/d11090158
APA StyleTitlyanov, E. A., Titlyanova, T. V., Tokeshi, M., & Li, X. (2019). Inventory and Historical Changes in the Marine Flora of Tomioka Peninsula (Amakusa Island), Japan. Diversity, 11(9), 158. https://doi.org/10.3390/d11090158