Thalassic Rotifers from the United States: Descriptions of Two New Species and Notes on the Effect of Salinity and Ecosystem on Biodiversity †



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Taxonomic Analysis
2.3. Data Analysis
2.4. Compilation of Checklist of U.S. Thalassic Rotifer Taxa
3. Results
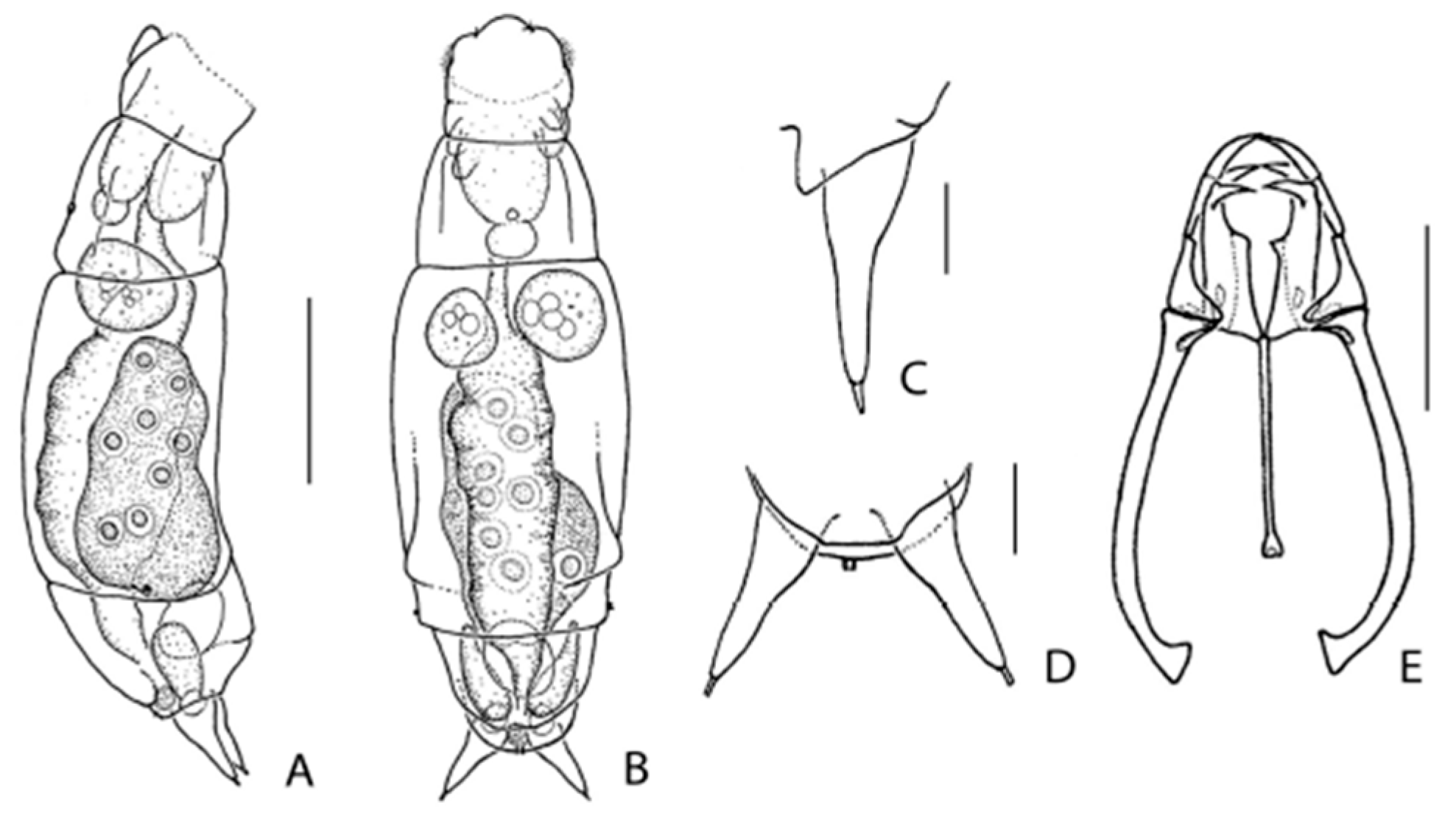
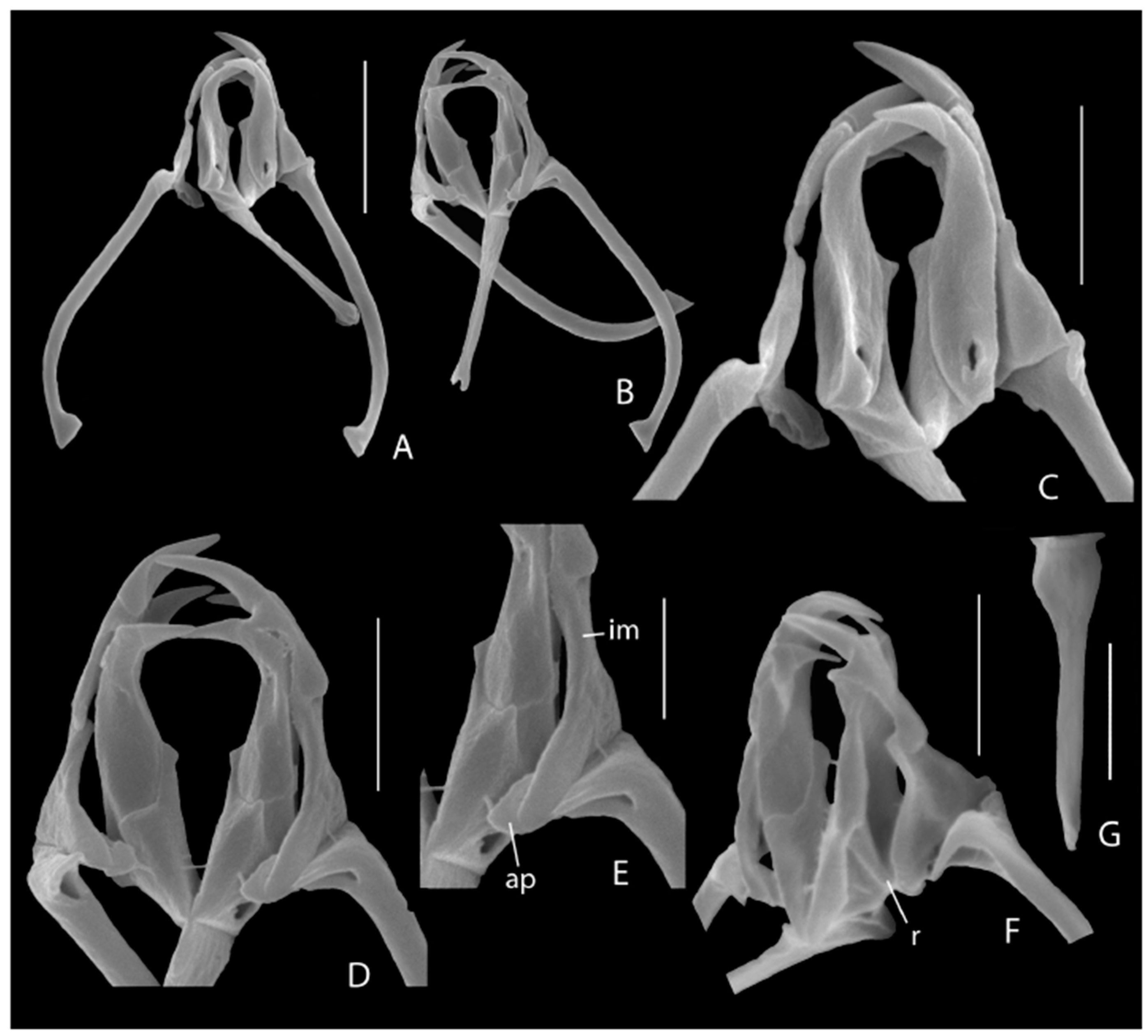
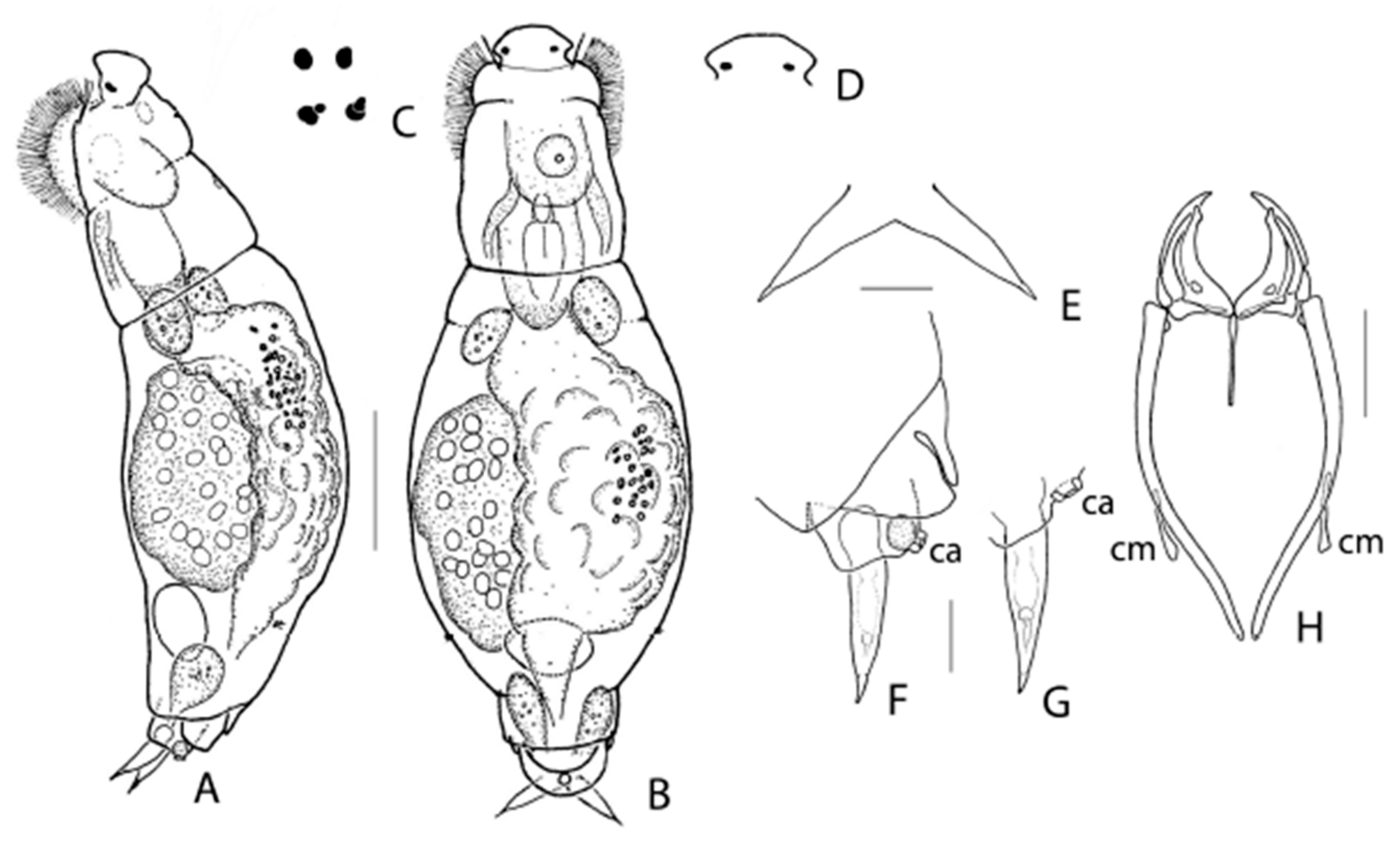
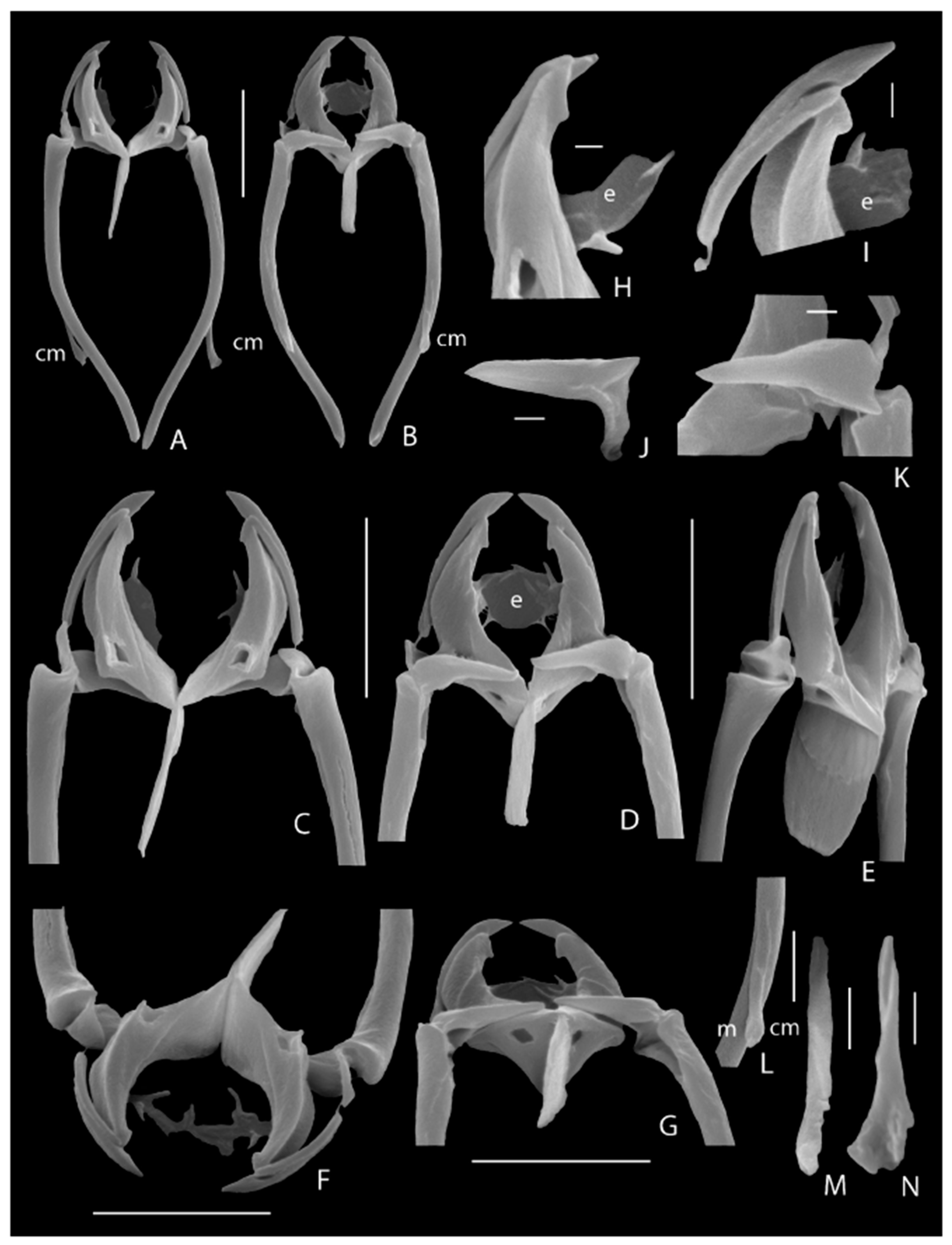
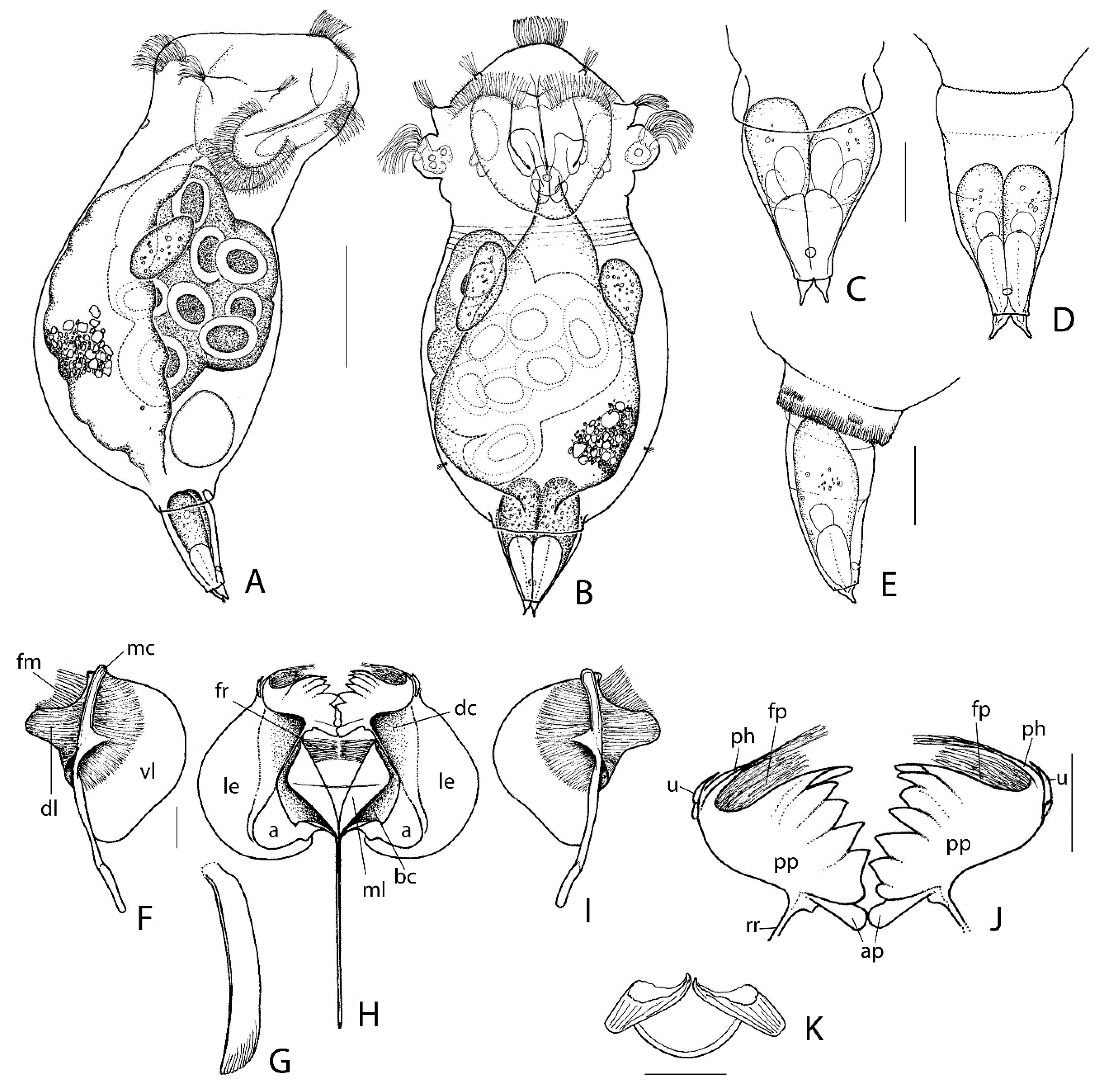
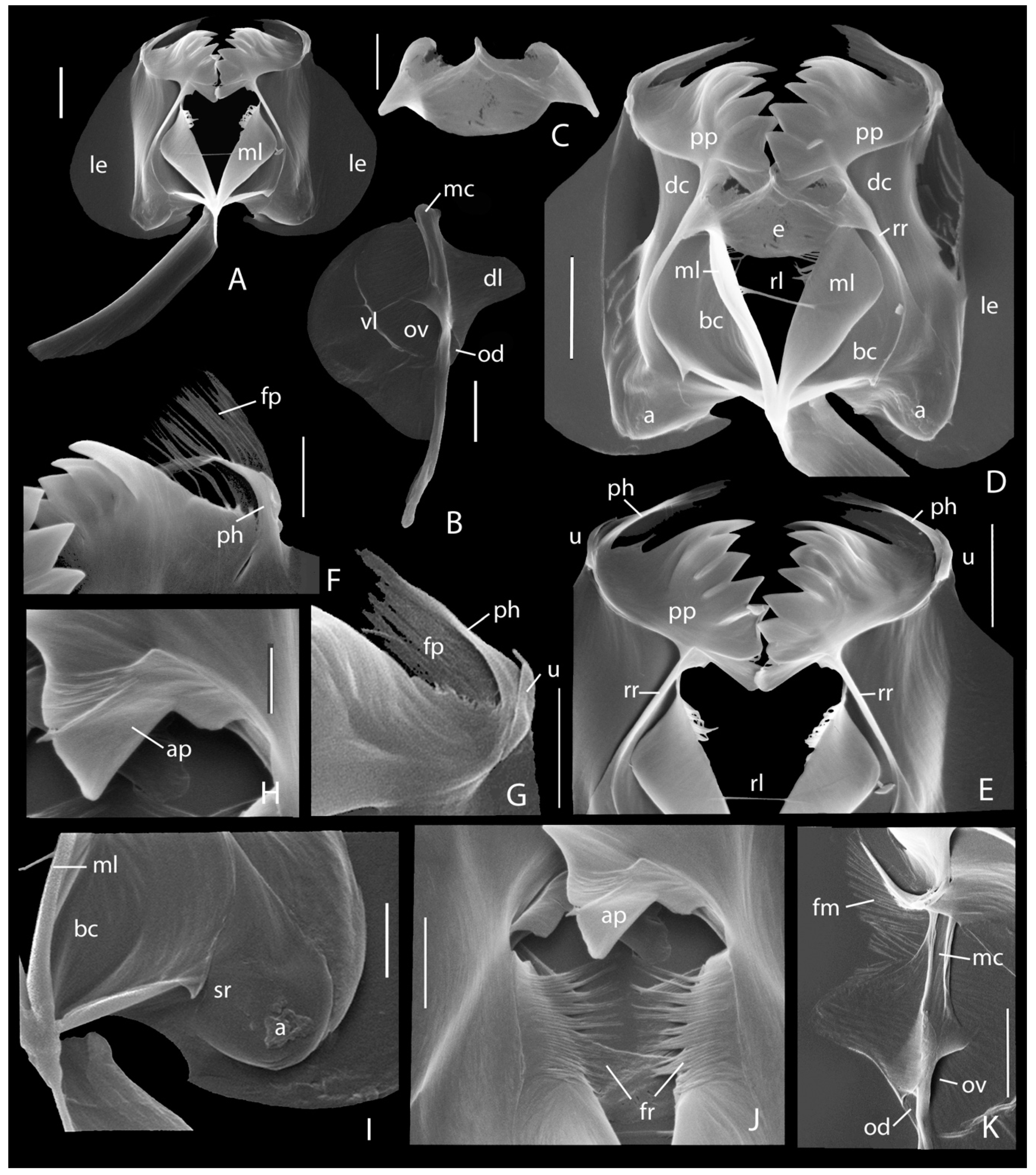
3.1. Taxonomic Analysis
3.2. Species Composition
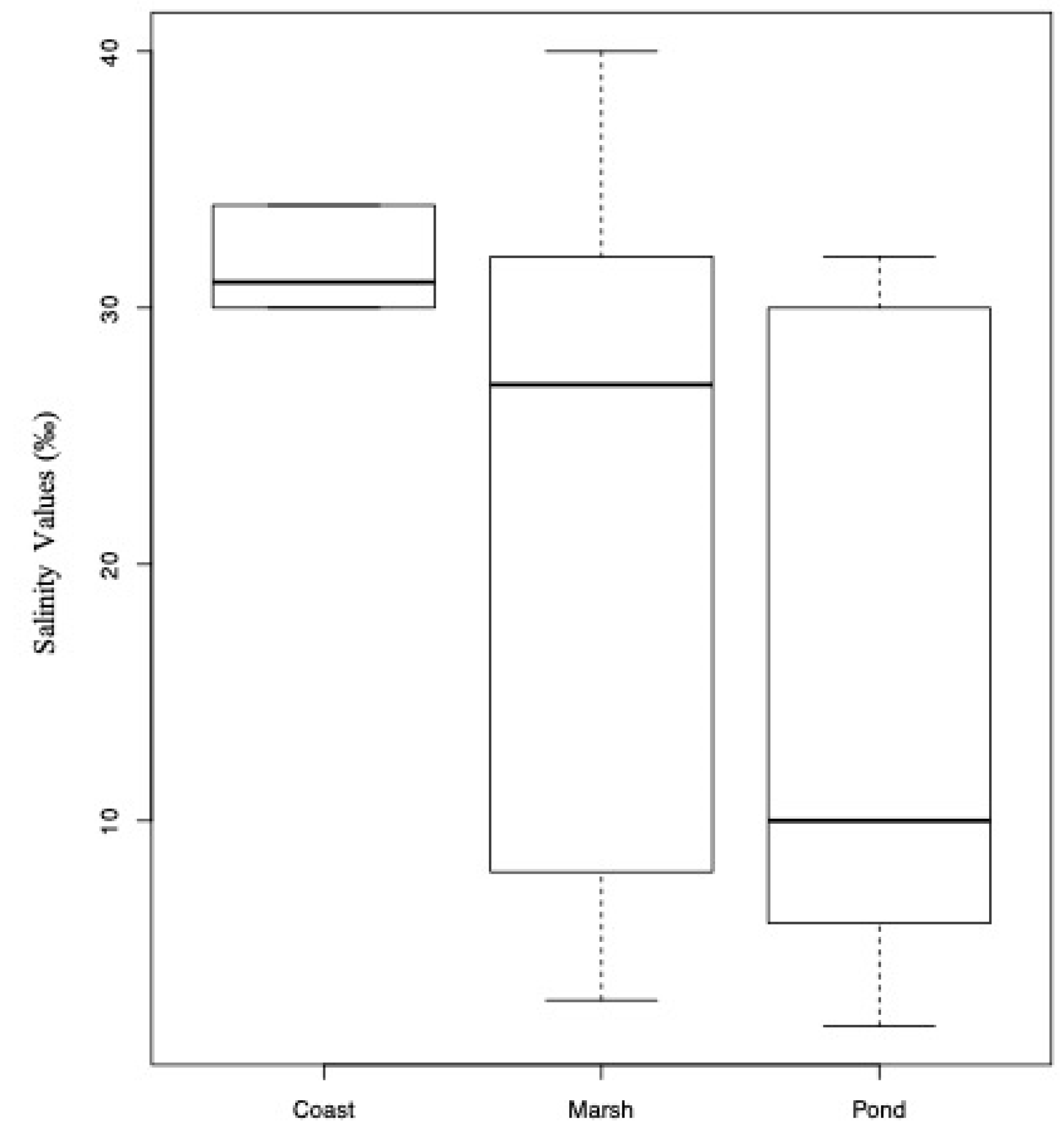
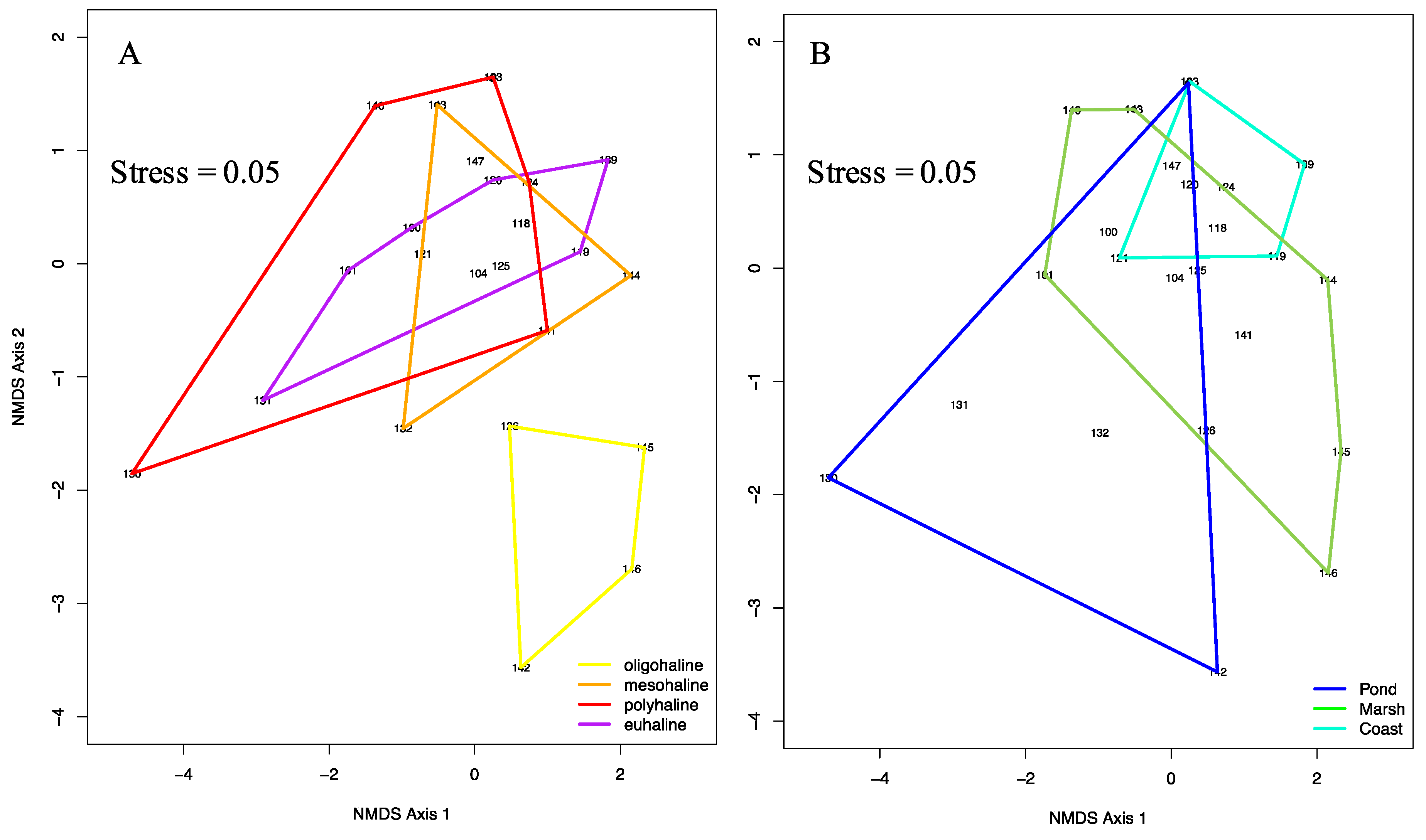
3.3. Community Ecology
3.4. Thalassic Rotifer Taxa from the U.S.
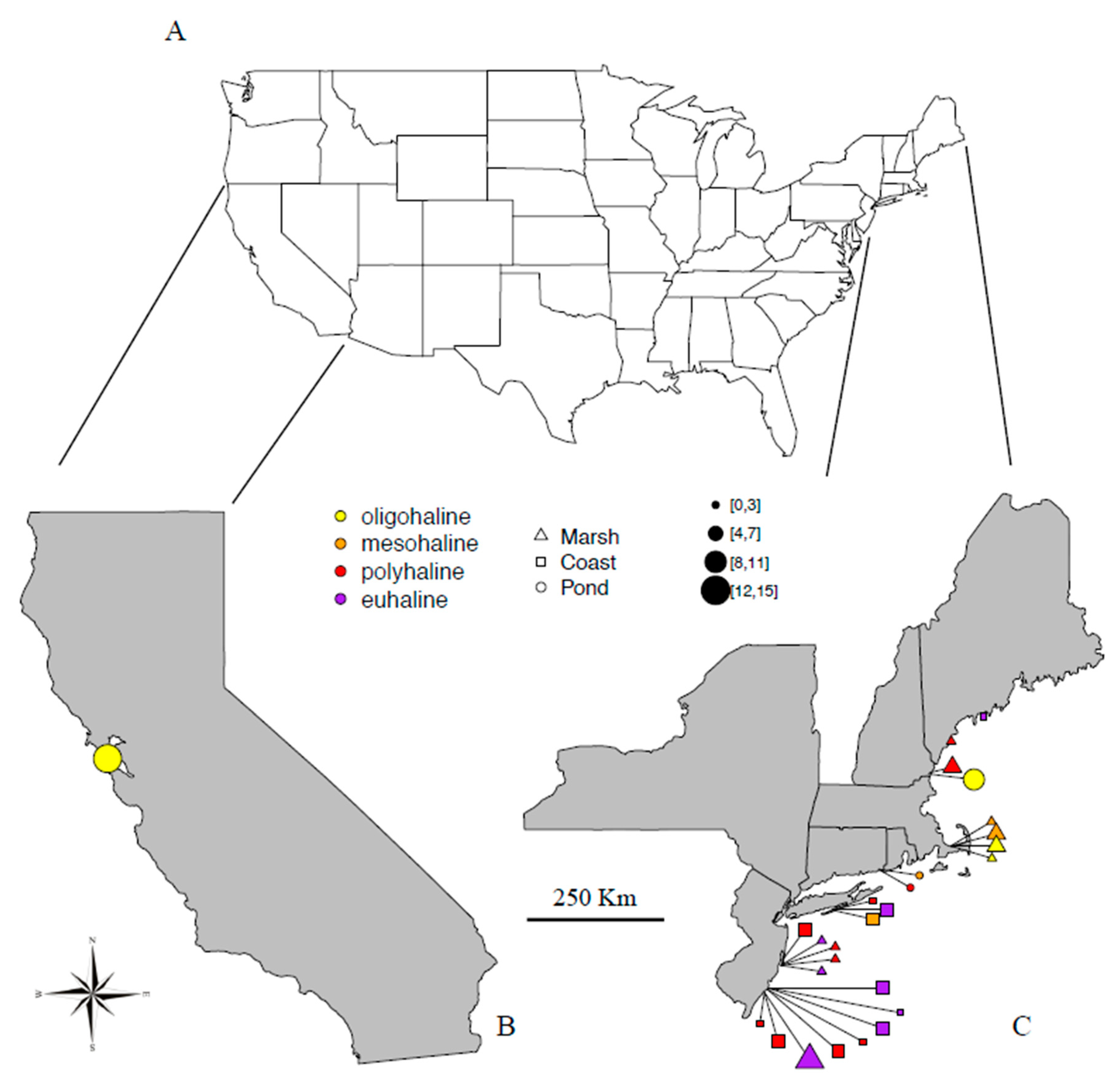
3.5. Distribution of U.S. Thalassic Rotifers
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Segers, H. Annotated checklist of the rotifers (Phylum Rotifera), with notes on nomenclature, taxonomy and distribution. Zootaxa 2007, 1546, 104. [Google Scholar] [CrossRef]
- Segers, H. Global diversity of rotifers (Rotifera) in freshwater. Hydrobiologia 2008, 595, 49–59. [Google Scholar] [CrossRef]
- Jersabek, C.D.; Leitner, M.F. The Rotifer World Catalog. World Wide Web Electronic Publication, 2013. Available online: http://www.rotifera.hausdernatur.at/Species/Index/222 (accessed on 15 June 2019).
- Fontaneto, D.; De Smet, W.H. Rotifera. In Handbook of Zoology: Gastrotricha, Cycloneuralia and Gnathifera. Volume 3: Gastrotricha and Gnathifera; Schmidt-Rhaesa, A., Ed.; Walter de Gruyter: Berlin, Germany, 2015; pp. 217–300. [Google Scholar]
- Wallace, R.L.; Snell, T.W. Chapter 8—Rotifera. In Ecology and Classification of North American Freshwater Invertebrates, 3rd ed.; Thorp, J.H., Covich, A.P., Eds.; Academic Press: San Diego, CA, USA, 2010; pp. 173–235. [Google Scholar]
- Fontaneto, D.; De Smet, W.H.; Ricci, C. Rotifers in saltwater environments, re-evaluation of an inconspicuous taxon. J. Mar. Biol. Assoc. U. K. 2006, 86, 623–656. [Google Scholar] [CrossRef]
- Fontaneto, D.; De Smet, W.H.; Melone, G. Identification key to the genera of marine rotifers worldwide. Meiofauna Mar. 2008, 16, 75–99. [Google Scholar]
- Appeltans, W.; Ahyong, S.T.; Anderson, G.; Angel, M.V.; Artois, T.; Bailly, N.; Bamber, R.; Barber, A.; Bartsch, I.; Berta, A.; et al. The Magnitude of Global Marine Species Diversity. Curr. Biol. 2012, 22, 2189–2202. [Google Scholar] [CrossRef]
- Curini-Galletti, M.; Artois, T.; Delogu, V.; De Smet, W.H.; Fontaneto, D.; Jondelius, U.; Leasi, F.; Martínez, A.; Meyer-Wachsmuth, I.; Nilsson, K.S.; et al. Patterns of diversity in soft-bodied meiofauna: Dispersal ability and body size matter. PLoS ONE 2012, 7, e33801. [Google Scholar] [CrossRef]
- De Smet, W.H.; Melone, G.; Fontaneto, D.; Leasi, F. Marine Rotifera. In Fauna D’Italia Volume 50; Calderini: Bologna, Italy, 2015; p. 252. [Google Scholar]
- Fontaneto, D.; Segers, H.; Melone, G. Marine rotifers from the Northern Adriatic Sea, with description of Lecane insulaconae sp. nov. (Rotifera: Monogononta: Lecanidae). J. Mar. Biol. Assoc. U. K. 2008, 88, 253–258. [Google Scholar] [CrossRef]
- Fontaneto, D.; Barbosa, A.M.; Segers, H.; Pautasso, M. The ‘rotiferologist’ effect and other global correlates of species richness in monogonont rotifers. Ecography 2012, 35, 174–182. [Google Scholar] [CrossRef]
- Remane, A. Rotatoria. In Die Tierwelt der Nord- und Ostsee, 16(7.e); Akademische Verlagsgesellschaft M.B.H.: Leipzig, Germany, 1929; p. 156. [Google Scholar]
- Dumont, H.J. Biogeography of Rotifers BT—Biology of Rotifers; Pejler, B., Starkweather, R., Nogrady, T., Eds.; Springer: Dordrecht, The Netherlands, 1983; pp. 19–30. [Google Scholar]
- Fautin, D.; Dalton, P.; Incze, L.S.; Leong, J.-A.C.; Pautzke, C.; Rosenberg, A.; Sandifer, P.; Sedberry, G.; Tunnell, J.W., Jr.; Abbott, I.; et al. An Overview of Marine Biodiversity in United States Waters. PLoS ONE 2010, 5, e11914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, W.R.; Perry, E.S. The coastal marine Tardigrada of the Americas. Zootaxa 2016, 4126, 375–396. [Google Scholar] [CrossRef]
- Leasi, F.; Sevigny, J.L.; Laflamme, E.M.; Artois, T.; Curini-Galletti, M.; de Jesus Navarrete, A.; Di Domenico, M.; Goetz, F.; Hall, J.A.; Hochberg, R.; et al. Biodiversity estimates and ecological interpretations of meiofaunal communities are biased by the taxonomic approach. Commun. Biol. 2018, 1, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, J.C. Synchaeta bicornis: A new rotifer from the brackish waters of Lake Pontchartrain, Louisiana. Trans. Am. Microsc. Soc. 1904, 25, 121–126. [Google Scholar] [CrossRef]
- Harring, H.K. A list of the Rotatoria of Washington and vicinity, with descriptions of a new genus and ten new species. Proc. U. S. Natl. Mus. 1913, 46, 387–405. [Google Scholar] [CrossRef] [Green Version]
- Harring, H.K.; Myers, F.J. The rotifer fauna of Wisconsin. Trans. Wis. Acad. Sci. Arts Lett. 1922, 20, 553–662. [Google Scholar]
- Harring, H.K.; Myers, F.J. The rotifer fauna of Wisconsin—II. A revision of the notommatid rotifers, exclusive of the Dicranophoridae. Trans. Wis. Acad. Sci. Arts Lett. 1924, 50, 415–549. [Google Scholar]
- Harring, H.K.; Myers, F.J. The rotifer fauna of Wisconsin—III. A revision of the genera Lecane and Monostyla. Trans. Wis. Acad. Sci. Arts Lett. 1926, 50, 315–423. [Google Scholar]
- Harring, H.K.; Myers, F.J. The rotifer fauna of Wisconsin—IV. The Dicranophoridae. Trans. Wis. Acad. Sci. Arts Lett. 1928, 23, 666–808. [Google Scholar]
- Myers, F.J. The distribution of Rotifera on Mount Desert Island. Am. Mus. Novit. 1931, 494, 12. [Google Scholar]
- Myers, F.J. The distribution of Rotifera on Mount Desert Island. Part VII. New Testudinellidae of the genus Testudinella and new species of Brachionidae of the genus Trichotria. Am. Mus. Novit. 1934, 761, 8. [Google Scholar]
- Myers, F.J. Three new brackish water and one new marine species of Rotatoria. Trans. Am. Microsc. Soc. 1936, 55, 428–432. [Google Scholar] [CrossRef]
- Edmondson, W.T. Rotatoria from Penikese Island, Massachusetts, with a description of Ptygura agassizi n. sp. Biol. Bull. 1948, 94, 263–266. [Google Scholar] [CrossRef] [PubMed]
- Björklund, B.G. Taxonomic and ecological studies of species of Notholca (Rotatoria) found in sea- and brackish water, with description of a new species. Sarsia 1972, 51, 25–66. [Google Scholar] [CrossRef]
- Ahlstrom, E. Rotatoria of Florida. Trans. Am. Microsc. Soc. 1934, 53, 251–266. [Google Scholar] [CrossRef]
- Ahlstrom, E. A revision of the rotatorian genera Brachionus and Platyias with descriptions of one new species and two new varieties. Bull. Am. Mus. Nat. Hist. 1940, 77, 148–184. [Google Scholar]
- Koste, W. Das Rädertier-Porträt. Einige auffalende Synchaeta-Arten aus Küstengewässern. Mikrokosmos 1981, 70, 169–176. [Google Scholar]
- Turner, P. Distribution of rotifers in a Floridian saltwater beach, with a note on rotifer dispersal. Hydrobiologia 1993, 255, 435–439. [Google Scholar] [CrossRef]
- Turner, P.N. Some Interstitial Rotifera from a Florida, U.S.A., Beach. Trans. Am. Microsc. Soc. 1990, 109, 417–421. [Google Scholar] [CrossRef]
- Harring, H.K. The Rotatoria of the Canadian Arctic Expedition, 1913–1918. Rep. Can. Arct. Exp. 1921, 8 Pt E, 3–23. [Google Scholar]
- Weber, E.F. Rotateurs (Voyage du Dr. W. Volz.). Zool. Jahrb. 1907, 24, 207–226. [Google Scholar]
- Hauer, J. Rotatorien aus dem “Zwischengebiet Wallacea”. 1. Teil. I. Das Genus Pedalia Barrois. Int. Rev. Der Gesamten Hydrobiol. Und Hydrogr. 1941, 41, 177–203. [Google Scholar] [CrossRef]
- Jersabek, C.D. Freshwater Rotifera (Monogononta) from Hawai’i, a preliminary checklist. Bishop Mus. Occas. Pap. 2003, 74, 46–72. [Google Scholar]
- Leasi, F.; Rouse, G.W.; Sørensen, M.V. A new species of Paraseison (Rotifera: Seisonacea) from the coast of California, USA. J. Mar. Biol. Assoc. U. K. 2012, 92, 959–965. [Google Scholar] [CrossRef]
- Heinbokel, J.; Coats, D.; Henderson, K.; Tyler, M. Reproduction rates and secondary production of three species of the rotifer genus Synchaeta in the estuarine Potomac River. J. Plankton Res. 1988, 10, 659–674. [Google Scholar] [CrossRef]
- Dolan, J.; Gallegos, C. Trophic coupling of rotifers, microflagellates, and bacteria during fall months in the Rhode River Estuary. Mar. Ecol. Prog. Ser. 1991, 77, 147–156. [Google Scholar] [CrossRef]
- Park, G.S.; Marshall, H.G. The trophic contributions of rotifers in tidal freshwater and estuarine habitats. Estuar. Coast. Shelf Sci. 2000, 51, 729–742. [Google Scholar] [CrossRef]
- Ekman, S. Zoogeography of the Sea; Sidgwick & Jackson Ltd.: London, UK, 1953; p. 417. [Google Scholar]
- Leasi, F.; Tang, C.Q.; De Smet, W.H.; Fontaneto, D. Cryptic diversity with wide salinity tolerance in the putative euryhaline Testudinella clypeata (Rotifera, Monogononta). Zool. J. Linn. Soc. 2013, 168, 17–28. [Google Scholar] [CrossRef]
- Anonymous. The Venice System for the Classification of Marine Waters According to Salinity. Limnol. Oceanogr. 1958, 3, 346–347. [Google Scholar]
- Willems, W.; Curini-Galletti, M.; Ferrero, T.; Fontaneto, D.; Heiner, I.; Huys, R.; Ivanenko, V.N.; Kristensen, R.M.; Kånneby, T.; MacNaughton, M.; et al. Meiofauna of the Koster-area, results from a workshop at the Sven Lovén Centre for Marine Sciences (Tjärnö, Sweden). Meiofauna Mar. 2009, 17, 1–34. [Google Scholar]
- Koste, W. Rotatoria. Die Rädertiere Mitteleuropas. Ein Bestimmungswerk, Begründet Von Max Voigt. Ueberordnung Monogononta, 2nd ed.; Tafelband, 234 Taf; Gebr. Borntraeger: Berlin, Germany; Stuttgart, Germany, 1978; p. 673. [Google Scholar]
- Nogrady, T.; Pourriot, R.; Segers, H. Rotifera. Volume 3. The Notommatidae and the Scaridiidae. In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World; Nogrady, T., Ed.; SPB Academic Publishing bv: The Hague, The Netherlands, 1995; p. 248. [Google Scholar]
- Segers, H. Rotifera, Volumr 2: The Lecanidae (Monogononta). In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World; Nogrady, T., Ed.; SPB Academic Publishing bv: The Hague, The Netherlands, 1995; p. 226. [Google Scholar]
- De Smet, W.H. Rotifera. Volume 4. The Proalidae (Monogononta). In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World; Nogrady, T., Ed.; SPB Academic Publishing bv: The Hague, The Netherlands, 1996; p. 102. [Google Scholar]
- De Smet, W.H. The Dicranophoridae and the Ituridae (Monogononta). In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World; Nogrady, T., Ed.; SPB Academic Publishing bv: The Hague, The Netherlands, 1997; p. 325. [Google Scholar]
- Nogrady, T.; Segers, H. Rotifera. Volume 6. The Asplanchnidae, Gastropodidae, Lindiidae, Microcodinidae, Synchaetidae, Trochosphaeridae and Filinia. In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World; Nogrady, T., Ed.; Backhuys Publishers: Leiden, The Netherlands, 2002; p. 264. [Google Scholar]
- Segers, H.; Fisher, W.; Fontaneto, D.; Michaloudi, E.; Wallace, R.L.; Jersabek, C.D. Towards a list of available names in zoology, partim Phylum Rotifera. Zootaxa 2012, 3179, 61–68. [Google Scholar] [CrossRef]
- Jersabek, C.D.; De Smet, W.H.; Hinz, C.; Fontaneto, D.; Michaloudi, E.; Wallace, R.L.; Segers, H. List of Available Names in Zoology, Candidate Part Phylum Rotifera, Species-Group Names Established before 1 January 2000. Final Version. 1 January 2018. Available online: https://www.iczn.org/assets/a42ca743cc/LAN_CandidatePart-SpeciesRotifera-2018-04-11.pdf (accessed on 13 January 2020).
- De Smet, W.H. Preparation of rotifer trophi for light and scanning electron microscopy. Hydrobiologia 1998, 387, 117–121. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Package ‘Vegan’. R Package Version 2.0-8. 2013, Volume 254. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 13 January 2020).
- Hervé, M. RVAideMemoire: Testing and Plotting Procedures for Biostatistics. R Package Version 0.9-72. 2019. Available online: https://cran.r-project.org/web/packages/RVAideMemoire/RVAideMemoire.pdf. (accessed on 13 January 2020).
- Faith, D.P.; Minchin, P.R.; Belbin, L. Compositional dissimilarity as a robust measure of ecological distance. Vegetatio 1987, 69, 57–68. [Google Scholar] [CrossRef]
- Chambers, J.M.; Freeny, A.E.; Heiberger, R.M. Analysis of variance; designed experiments. In Statistical Models in S; Routledge: New York, NY, USA, 2017; pp. 145–193. [Google Scholar]
- Ambler, J.; Cloern, J.; Hutchinson, A. Seasonal cycles of zooplankton from San Francisco Bay. Hydrobiologia 1985, 129, 177–197. [Google Scholar] [CrossRef]
- Battaglene, S.; Talbot, B. Mass production of rotifers. Austasia Aquac. Mag. 1989, 3, 6–7. [Google Scholar]
- Chester, A.J. Microzooplankton in the Surface Waters of the Strait of Juan de Fuca. NOAA Technical Report ERL-403-PM. 1978, p. 26. Available online: http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.514.763&rep=rep1&type=pdf (accessed on 13 January 2020).
- Dolan, J.R.; Gallegos, C.C. Trophic role of planktonic rotifers in the Rhode River Estuary, spring-summer 1991. Mar. Ecol. Prog. Ser. 1992, 85, 187–199. [Google Scholar] [CrossRef]
- Fielder, D.S.; Purser, G.J.; Battaglene, S.C. Effect of rapid changes in temperature and salinity on availability of the rotifers Brachionus rotundiformis and Brachionus plicatilis. Aquaculture 2000, 189, 85–99. [Google Scholar] [CrossRef]
- Fish, C.J.; Johnson, M.W. The Biology of the Zooplankton Population in the Bay of Fundy and Gulf of Maine with Special Reference to Production and Distribution. J. Biol. Board Can. 1937, 3, 189–322. [Google Scholar] [CrossRef]
- Fradkin, S.C. Rotifer Distributions in the Coastal Waters of the Northeast Pacific Ocean BT—Rotifera IX; Sanoamuang, L., Segers, H., Shiel, R.J., Gulati, R.D., Eds.; Springer: Dordrecht, The Netherlands, 2001; pp. 173–177. [Google Scholar]
- Guilini, K.; Levin, L.A.; Vanreusel, A. Cold seep and oxygen minimum zone associated sources of margin heterogeneity affect benthic assemblages, diversity and nutrition at the Cascadian margin (NE Pacific Ocean). Prog. Oceanogr. 2012, 96, 77–92. [Google Scholar] [CrossRef]
- Holland, J.S.; Maciolek, N.J.; Kalke, R.D.; Mullins, L.; Oppenheimer, C.H. A benthos and plankton study of the Corpus Christi, Copano and Aransas Bay systems. In Report on Data Collected during the Period July 1973–April 1974; University of Texas Marine Science Institute: Port Aransas, TX, USA, 1974; p. 121. [Google Scholar]
- Hopkins, T.L. The Plankton of the St. Andrew Bay System, Florida; Collections Galveston Bay Bibliography: Galveston, TX, USA, 1966; pp. 12–64. [Google Scholar]
- Hopkins, T.L. Zooplankton distribution in surface waters of Tampa Bay, Florida. Bull. Mar. Sci. 1977, 27, 467–478. [Google Scholar]
- Lee, W.Y.; Macko, S.A.; Ciereszko, L.S. Toxic effects of cembranolides derived from octocorals on the rotifer Brachionus plicatilis Müller and the amphipod Parhyale hawaiensis (Dana). J. Exp. Mar. Biol. Ecol. 1981, 54, 91–96. [Google Scholar] [CrossRef]
- Martin, J.H. Phytoplankton-zooplankton relationships In Arragansett Bay 1. Limnol. Oceanogr. 1965, 10, 185–191. [Google Scholar] [CrossRef]
- Myers, F.J. The rotifer fauna of Wisconsin. V. The genera Euchlanis and Monommata. Trans. Wis. Acad. Sci. 1930, 25, 353–413. [Google Scholar]
- Sellner, K.G.; Lacouture, R.V.; Cibik, S.J.; Brindley, A.; Brownlee, S.G. Importance of a winter dinoflagellate-microflagellate bloom in the Patuxent River estuary. Estuar. Coast. Shelf Sci. 1991, 32, 27–42. [Google Scholar] [CrossRef]
- Sommer, S.; Gutzmann, E.; Ahlrichs, W.; Pfannkuche, O. Rotifers colonising sediments with shallow gas hydrates. Naturwissenschaften 2003, 90, 273–276. [Google Scholar] [CrossRef] [PubMed]
- Sommer, S.; Gutzmann, E.; Pfannkuche, O. Sediments hosting gas hydrate: Oases for metazoan meiofauna. Mar. Ecol. Prog. Ser. 2007, 337, 27–37. [Google Scholar] [CrossRef]
- Suchar, V.A.; Chigbu, P. The effects of algae species and densities on the population growth of the marine rotifer, Colurella dicentra. J. Exp. Mar. Biol. Ecol. 2006, 337, 96–102. [Google Scholar] [CrossRef]
- Tunnell, J.W., Jr.; Alvarado, S.A. Checklist of Species within Corpus Christi Bay National Estuary Program Study Area: References, Habitats, Distribution, and Abundance; Corpus Christi Bay National Estuary Program CCBNEP-06D; Texas National Resource Conservation Commission: Austin, TX, USA, 1996; p. 298. [Google Scholar]
- Winder, M.; Jassby, A.D. Shifts in zooplankton community structure: Implications for food web processes in the upper San Francisco Estuary. Estuar. Coasts 2011, 34, 675–690. [Google Scholar] [CrossRef] [Green Version]
- Jersabek, C.D.; Segers, H.; Dingmann, B.J. The Frank J. In Myers Rotifera Collection; The Whole Collection in Digital Images; The Academy of Natural Sciences of Philadelphia, Special Publication 20: Philadelphia, PA, USA, 2003. [Google Scholar]
- De Smet, W.H. Rotifera from the Mediterranean Sea, with description of ten new species. Zootaxa 2015, 4028, 151–196. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, M.V.; Kristensen, R.M. Marine Rotifera from Ikka Fjord, SW Greenland. With the description of a new species from the rare mineral ikaite. Meddelelser Om Grønland Bioscience 2000, 51, 46. [Google Scholar]
- Althaus, B. Neue Sandbodenrotatorien aus dem Schwarzen Meer. Wiss. Z. Univ. Halle Math. Nat. 1957, 6, 445–458. [Google Scholar]
- Jersabek, C.D. The ‘Frank J. Myers Rotifera collection’ at the Academy of Natural Sciences of Philadelphia BT—Rotifera X: Rotifer Research: Trends, New Tools and Recent Advances. In Proceedings of the International Rotifer Symposium, Illmitz, Austria, 7–13 June 2003; Herzig, A., Gulati, R.D., Jersabek, C.D., May, L., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 137–140. [Google Scholar]
- De Ridder, M.; Segers, H. Monogonont Rotifera Recorded in the World Literature (Except Africa) from 1960 to 1992; KBIN: Bruxelles, Bélgica, 1997; Volume 88, pp. 3–481. [Google Scholar]
- Litton, J.R. The marine interstitial and planktonic rotifer fauna of Puerto Rico. Proc. Indiana Acad. Sci. 1982, 92, 470. [Google Scholar]
- Wilke, T.; Ahlrichs, W.H.; Bininda-Emonds, O.R.P. A weighted taxonomic matrix key for species of the rotifer genus Synchaeta (Rotifera, Monogononta, Synchaetidae). Zookeys 2019, 871, 1–40. [Google Scholar] [CrossRef] [Green Version]
- Hollowday, E.D. Family Synchaetidae Hudson and Gosse, 1886. Rotifera 6: Asplanchnidae, Gastropodidae, Lindiidae, Microcodidae, Synchaetidae, Trochosphaeridae and Filinia. In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World; Nogrady, T., Ed.; Backhuys Publishers: Leiden, The Netherlands, 2002; pp. 87–211. [Google Scholar]
- De Smet, W.H. Cotylegaleatidae, a new family of Ploima (Rotifera: Monogononta), for Cotylegaleata perplexa gen. et sp. nov., from freshwater benthos of Belgium. Zootaxa 2007, 1425, 35–43. [Google Scholar] [CrossRef]
- Peters, F. Untersuchungen über Anatomie und Zellkonstanz von Synchaeta (S. grimpei Remane, S. baltica Ehr., S. tavina Hood und S. triophthalma Laut). Ein Beitrag der Artunterschiede bei konstantzelligen Tieren. Zeitschrift Für Wissenschaftliche Zoologie 1931, 139, 1–119. [Google Scholar]
- Remane, A. Rotatoria (4. Lieferung). HG Bronn’s Klassen und Ordnungen desTier-Reichs. In Vierter Band Vermes. II; Abteilung Aschelminthen. I. B. Rotatorien, Gastrotrichen und Kinorhynchen; Akademische Verlagsgesellschaft: Leipzig, Germany, 1933; pp. 452–576. [Google Scholar]
- Spalding, M.D.; Fox, H.E.; Allen, G.R.; Davidson, N.; Ferdana, Z.A.; Finlayson, M.; Halpern, B.S.; Jorge, M.A.; Lombana, A.; Lourie, S.A. Marine ecoregions of the world: A bioregionalization of coastal and shelf areas. BioScience 2007, 57, 573–583. [Google Scholar] [CrossRef] [Green Version]
- Friedrich, C.; De Smet, W.H. The rotifer fauna of arctic sea ice from the Barents Sea, Laptev Sea and Greenland Sea. Hydrobiologia 2000, 432, 73–90. [Google Scholar] [CrossRef]
- Devetter, M.; Fontaneto, D.; Jersabek, C.D.; Mark Welch, D.B.; May, L.; Walsh, E.J. Preface: Evolving rotifers, evolving science. Hydrobiologia 2017, 796, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Obertegger, U.; Flaim, G. Taxonomic and functional diversity of rotifers, what do they tell us about community assembly? Hydrobiologia 2018, 823, 79–91. [Google Scholar] [CrossRef]
- Mills, S.; Alcántara-Rodríguez, J.A.; Ciros-Pérez, J.; Gómez, A.; Hagiwara, A.; Galindo, K.H.; Jersabek, C.D.; Malekzadeh-Viayeh, R.; Leasi, F.; Lee, J.-S.; et al. Fifteen species in one: Deciphering the Brachionus plicatilis species complex (Rotifera, Monogononta) through DNA taxonomy. Hydrobiologia 2016, 796, 39–58. [Google Scholar] [CrossRef] [Green Version]
- Martínez, A.; Di Domenico, M.; Leasi, F.; Curini-Galletti, M.; Todaro, M.A.; Zotto, M.D.; Gobert, S.; Artois, T.; Norenburg, J.; Jörger, K.M.; et al. Patterns of diversity and endemism of soft-bodied meiofauna in an oceanic island, Lanzarote, Canary Islands. Mar. Biodivers. 2019, 49, 2033–2055. [Google Scholar] [CrossRef] [Green Version]
- Rougier, C.; Pourriot, R.; Lam-Hoai, T. The genus Synchaeta (rotifers) in a north-western Mediterranean coastal lagoon (Etang de Thau, France): Taxonomical and ecological remarks. Hydrobiologia 2000, 436, 105–117. [Google Scholar] [CrossRef]
- Fonseca, V.G.; Carvalho, G.R.; Nichols, B.; Quince, C.; Johnson, H.F.; Neill, S.P.; Lambshead, J.D.; Thomas, W.K.; Power, D.M.; Creer, S. Metagenetic analysis of patterns of distribution and diversity of marine meiobenthic eukaryotes. Glob. Ecol. Biogeogr. 2014, 23, 1293–1302. [Google Scholar] [CrossRef] [Green Version]
- Carugati, L.; Corinaldesi, C.; Dell’Anno, A.; Danovaro, R. Metagenetic tools for the census of marine meiofaunal biodiversity: An overview. Mar. Genom. 2015, 24, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Gómez, A.; Carmona, M.J.; Serra, M. Ecological factors affecting gene flow in the Brachionus plicatilis complex (Rotifera). Oecologia 1997, 111, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Khlebovich, V.V.; Aladin, N.V. The salinity factor in animal life. Her. Russ. Acad. Sci. 2010, 80, 299–304. [Google Scholar] [CrossRef]
- McNamara, J.C.; Faria, S.C. Evolution of osmoregulatory patterns and gill ion transport mechanisms in the decapod Crustacea: A review. J. Comp. Physiol. B 2012, 182, 997–1014. [Google Scholar] [CrossRef]
- Leasi, F.; Andrade, S.C.D.S.; Norenburg, J. At least some meiofaunal species are not everywhere. Indication of geographic, ecological and geological barriers affecting the dispersion of species of Ototyphlonemertes (Nemertea, Hoplonemertea). Mol. Ecol. 2016, 25, 1381–1397. [Google Scholar] [CrossRef]
- Leasi, F.; Gaynus, C.; Mahardini, A.; Moore, T.N.; Norenburg, J.L.; Barber, P.H. Spatial and ecologic distribution of neglected microinvertebrate communities across endangered ecosystems: Meiofauna in Bali (Indonesia). Mar. Ecol. 2016, 37, 970–987. [Google Scholar] [CrossRef]
- Lefeuvre, J.-C.; Laffaille, P.; Feunteun, E.; Bouchard, V.; Radureau, A. Biodiversity in salt marshes: From patrimonial value to ecosystem functioning. The case study of the Mont-Saint-Michel bay. C. R. Biol. 2003, 326, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Visser, J.M.; Midway, S.; Baltz, D.M.; Sasser, C.E. Chapter 15—Ecosystem Structure of Tidal Saline Marshes; Perillo, G.M.E., Wolanski, E., Cahoon, D.R., Hopkinson, C.S.B.T.-C.W., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 519–538. [Google Scholar]
- Akter, R.; Asik, T.Z.; Sakib, M.; Akter, M.; Sakib, M.N.; Azad, A.; Alauddin, A.S.M.; Maruf, M.; Haque, A.; Rahman, M. The dominant climate change event for salinity intrusion in the GBM Delta. Climate 2019, 7, 69. [Google Scholar] [CrossRef] [Green Version]
- Zeppilli, D.; Sarrazin, J.; Leduc, D.; Arbizu, P.M.; Fontaneto, D.; Fontanier, C.; Gooday, A.J.; Kristensen, R.M.; Ivanenko, V.N.; Sørensen, M.V. Is the meiofauna a good indicator for climate change and anthropogenic impacts? Mar. Biodivers. 2015, 45, 505–535. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site ID | State * | Site | Salinity ‰ | Salinity Group | Ecosystem | Lat N | Long W | Venice Classification | Date of Sampling mm/dd/2012 |
|---|---|---|---|---|---|---|---|---|---|
| 100 | NJ | Barnegat Bay | 34 | Marine | Marsh | 39.7917 | 74.113 | euhaline | 03/30 |
| 101 | NJ | Barnegat Bay | 40 | Marine | Marsh | 39.7904 | 74.1150 | euhaline | 03/30 |
| 102 | NJ | Barnegat Bay | 30 | Marine | Marsh | 39.7905 | 74.1171 | polyhaline | 03/30 |
| 103 | NJ | Barnegat Bay | 30 | Marine | Marsh | 39.7909 | 74.1171 | polyhaline | 03/30 |
| 104 | NJ | Barnegat Bay | 24 | Marine | Marsh | 39.8718 | 74.0822 | polyhaline | 03/30 |
| 118 | NJ | Brigantine | 34 | Marine | Coast | 39.4192 | 74.3796 | euhaline | 06/05 |
| 119 | NJ | Brigantine | 34 | Marine | Coast | 39.4191 | 74.3800 | euhaline | 06/05 |
| 120 | NJ | Brigantine | 34 | Marine | Coast | 39.4191 | 74.3800 | euhaline | 06/05 |
| 121 | NJ | Brigantine | 30 | Marine | Coast | 39.3947 | 74.4030 | polyhaline | 06/05 |
| 122 | NJ | Brigantine | 30 | Marine | Coast | 39.3949 | 74.4026 | polyhaline | 06/05 |
| 123 | NJ | Calvert | 30 | Marine | Coast | 39.3459 | 74.4765 | polyhaline | 06/05 |
| 124 | NJ | Calvert | 30 | Marine | Coast | 39.3460 | 74.4761 | polyhaline | 06/05 |
| 125 | NJ | Calvert | 34 | Marine | Marsh | 39.3526 | 74.4790 | euhaline | 06/05 |
| 126 | CA | San Francisco | 4 | Brackish | Pond | 37.7803 | 122.5139 | oligohaline | 07/09 |
| 130 | NY | Fire Island | 30 | Marine | Pond | 40.6927 | 72.9882 | polyhaline | 07/31 |
| 131 | NY | Fire Island | 32 | Marine | Pond | 40.6939 | 72.9868 | euhaline | 07/ 31 |
| 132 | NY | Fire Island | 8 | Brackish | Pond | 40.6915 | 72.9863 | mesohaline | 07/ 31 |
| 139 | ME | Boothbay | 32 | Marine | Coast | 43.8422 | 69.6545 | euhaline | 09/28 |
| 140 | ME | Biddeford | 20 | Brackish | Marsh | 43.4395 | 70.3654 | polyhaline | 09/28 |
| 141 | NH | Hampton | 30 | Marine | Marsh | 42.9295 | 70.8405 | polyhaline | 09/28 |
| 142 | NH | Hampton | 2 | Brackish | Pond | 42.9078 | 70.8667 | oligohaline | 09/28 |
| 143 | MA | Cape Cod Barnstable | 6 | Brackish | Marsh | 41.7338 | 70.3794 | mesohaline | 09/30 |
| 144 | MA | Cape Cod Barnstable | 10 | Brackish | Marsh | 41.7333 | 70.3794 | mesohaline | 09/30 |
| 145 | MA | Cape Cod Barnstable | 3 | Brackish | Marsh | 41.7332 | 70.3794 | oligohaline | 09/30 |
| 146 | MA | Cape Cod Barnstable | 4 | Brackish | Marsh | 41.7333 | 70.3794 | oligohaline | 09/30 |
| 147 | CT | Mystic | 10 | Brackish | Pond | 41.3563 | 71.9644 | mesohaline | 09/30 |
| 148 | CT | Mystic | 30 | Marine | Pond | 41.3562 | 71.9648 | polyhaline | 09/30 |
| BDELLOIDEA |
| Bdelloidea indet.: 131, 142, 144, 145, 146, 147 |
| Philodinidae |
| * Philodina citrina Ehrenberg, 1830: 126 |
| * Rotaria rotatoria (Pallas, 1766): 126, 132 |
| MONOGONONTA |
| Brachionidae |
| Keratella americana Carlin, 1943: 125 |
| Notholca marina Focke, 1961: 126, 145, 146 |
| * Plationus patulus (Müller, 1786): 142 |
| Dicranophoridae |
| Encentrum algente Harring, 1921: 141 |
| * E. graingeri Chengalath, 1985: 145 |
| E. marinum (Dujardin, 1841): 100, 101, 104, 121 |
| * E. melonei sp. nov.: 121, 124, 125, 139 |
| E. rousseleti (Lie-Pettersen, 1905): 125, 144 |
| E. villosum Harring and Myers, 1928: 101, 121, 125 |
| * E. cf. villosum Harring and Myers, 1928: 142 |
| Gastropodidae |
| * Gastropus hyptopus (Ehrenberg, 1838): 142 |
| Lecanidae |
| Lecane closterocerca (Schmarda, 1859): 126 |
| L. grandis (Murray, 1913): 130, 131 |
| L. hastata (Murray, 1913): 131 |
| L. luna (Müller, 1776): 126 |
| L. punctata (Murray, 1913): 132 |
| * L. quadridentata (Ehrenberg, 1830): 126 |
| Lepadellidae |
| Colurella adriatica Ehrenberg, 1831: 104, 118, 119, 120, 125, 126, 141 |
| C. colurus (Ehrenberg, 1830): 104, 118, 125, 132 |
| C. colurus compressa Lucks, 1912: 126, 132 |
| C. dicentra (Gosse, 1887): 100, 104, 120, 121, 124, 125, 126, 141, 147 |
| C. obtusa (Gosse, 1886): 125, 126 |
| * C. uncinata (Müller, 1773): 139 |
| * C. unicauda Eriksen, 1968: 120, 122, 123 |
| * Lepadella patella persimilis De Ridder, 1961: 126 |
| Lindiidae |
| Lindia tecusa Harring and Myers, 1922: 141, 144, 145 |
| Mytilinidae |
| * Lophocharis oxysternon (Gosse, 1851): 126 |
| * Mytilina brevispina (Ehrenberg, 1830): 126 |
| Notommatidae |
| * Cephalodella sp. [undescribed species]: 131, 132 |
| * Monommata sp.: 142 |
| Pleurotrocha atlantica Myers, 1936: 131 |
| Proalidae |
| Proales halophila Remane, 1929: 104 |
| P. reinhardti (Ehrenberg, 1834): 104, 118, 119, 124, 125, 139, 144 |
| P. similis Beauchamp, 1907: 121, 131 |
| Synchaetidae |
| * Synchaeta grossa sp. nov.: 100, 102, 103, 104, 121, 125, 140, 143 |
| Testudinellidae |
| Testudinella clypeata (Müller, 1786): 102, 103, 118, 120, 121, 122, 123, 124, 125, 143, 146, 148 |
| Trichocercidae |
| * Trichocerca brachyura (Gosse, 1851): 126, 142 |
| * T. longiseta (Schrank, 1802): 142 |
| * T. tigris (Müller, 1786): 142 |
| * Trichocerca sp. [undescribed species]: 126, 142 |
| Trochosphaeridae |
| Filinia sp.: 142 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leasi, F.; De Smet, W.H. Thalassic Rotifers from the United States: Descriptions of Two New Species and Notes on the Effect of Salinity and Ecosystem on Biodiversity. Diversity 2020, 12, 28. https://doi.org/10.3390/d12010028
Leasi F, De Smet WH. Thalassic Rotifers from the United States: Descriptions of Two New Species and Notes on the Effect of Salinity and Ecosystem on Biodiversity. Diversity. 2020; 12(1):28. https://doi.org/10.3390/d12010028
Chicago/Turabian StyleLeasi, Francesca, and Willem H. De Smet. 2020. "Thalassic Rotifers from the United States: Descriptions of Two New Species and Notes on the Effect of Salinity and Ecosystem on Biodiversity" Diversity 12, no. 1: 28. https://doi.org/10.3390/d12010028
APA StyleLeasi, F., & De Smet, W. H. (2020). Thalassic Rotifers from the United States: Descriptions of Two New Species and Notes on the Effect of Salinity and Ecosystem on Biodiversity. Diversity, 12(1), 28. https://doi.org/10.3390/d12010028