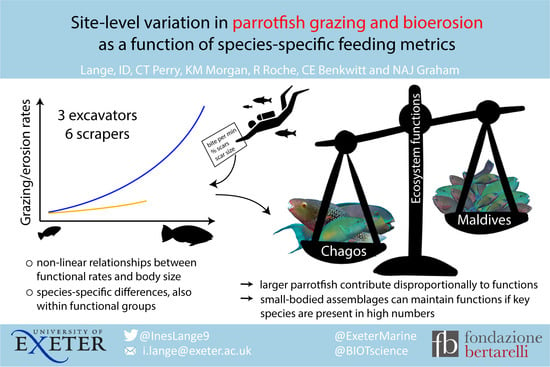
Site-Level Variation in Parrotfish Grazing and Bioerosion as a Function of Species-Specific Feeding Metrics

 , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
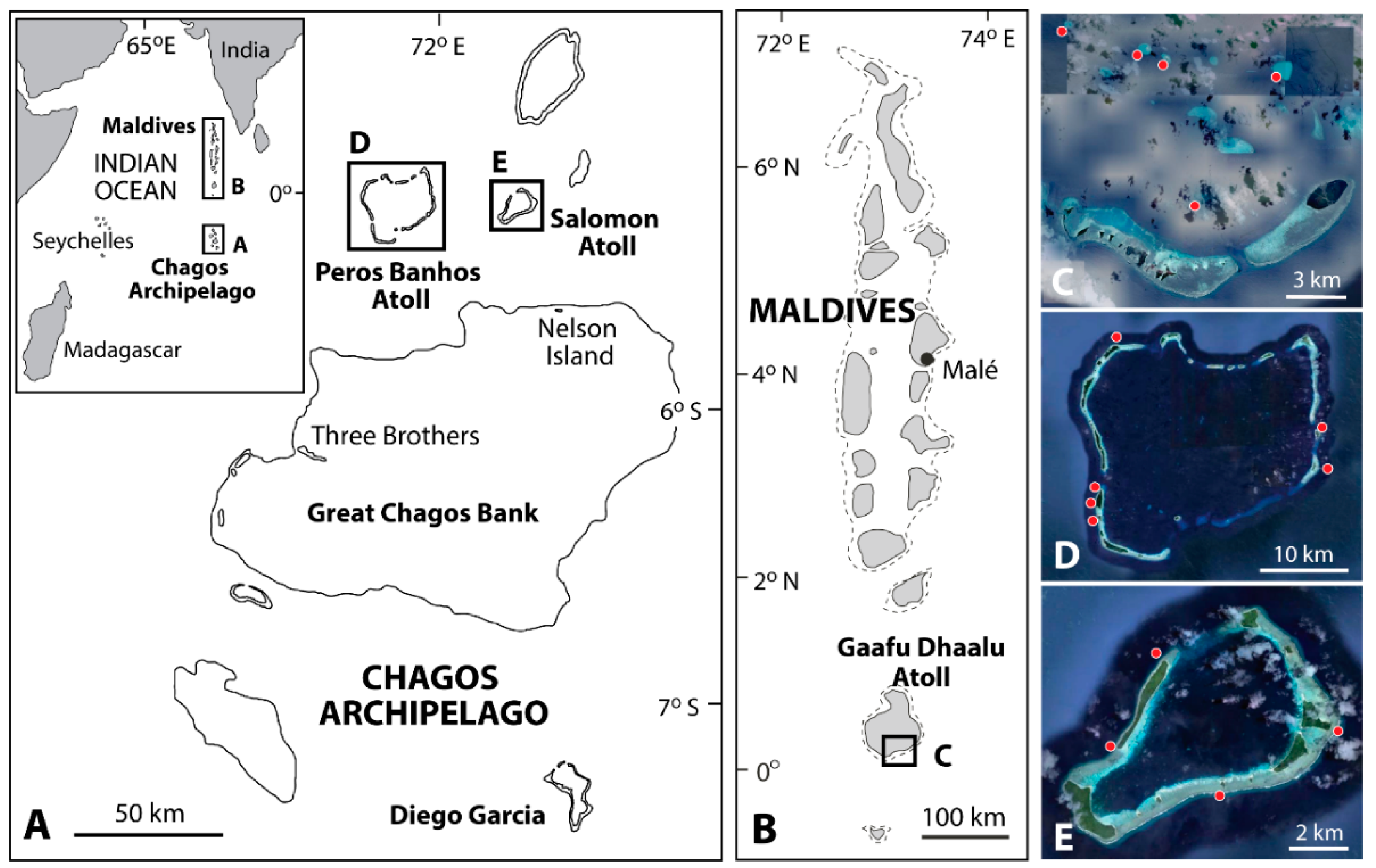
2.1. Study Sites and Parrotfish Species
2.2. Feeding metrics
2.3. Individual Grazing and Bioerosion Rates
2.4. Community Grazing and Bioerosion Levels
2.5. Statistical Analysis
3. Results
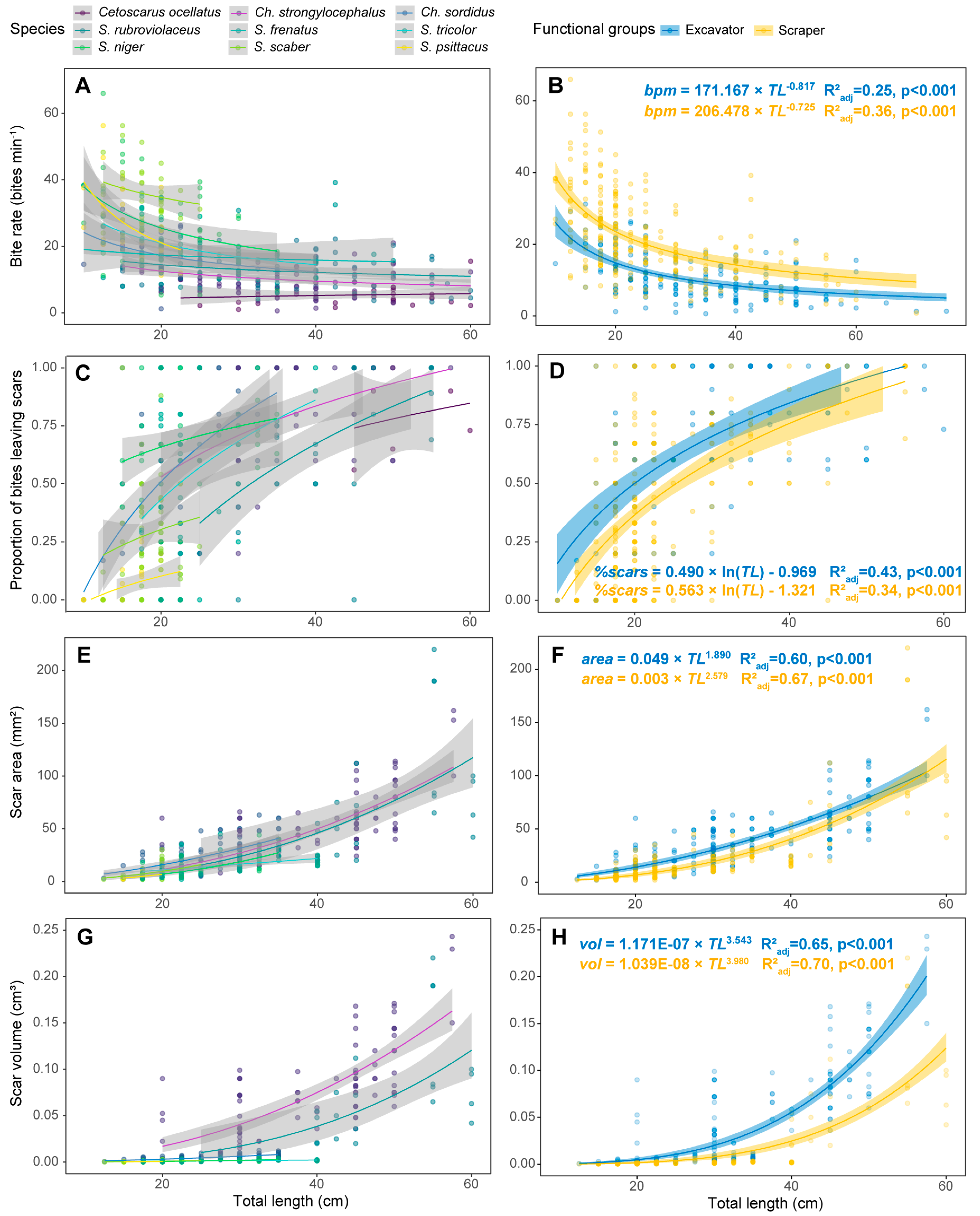
3.1. Feeding metrics
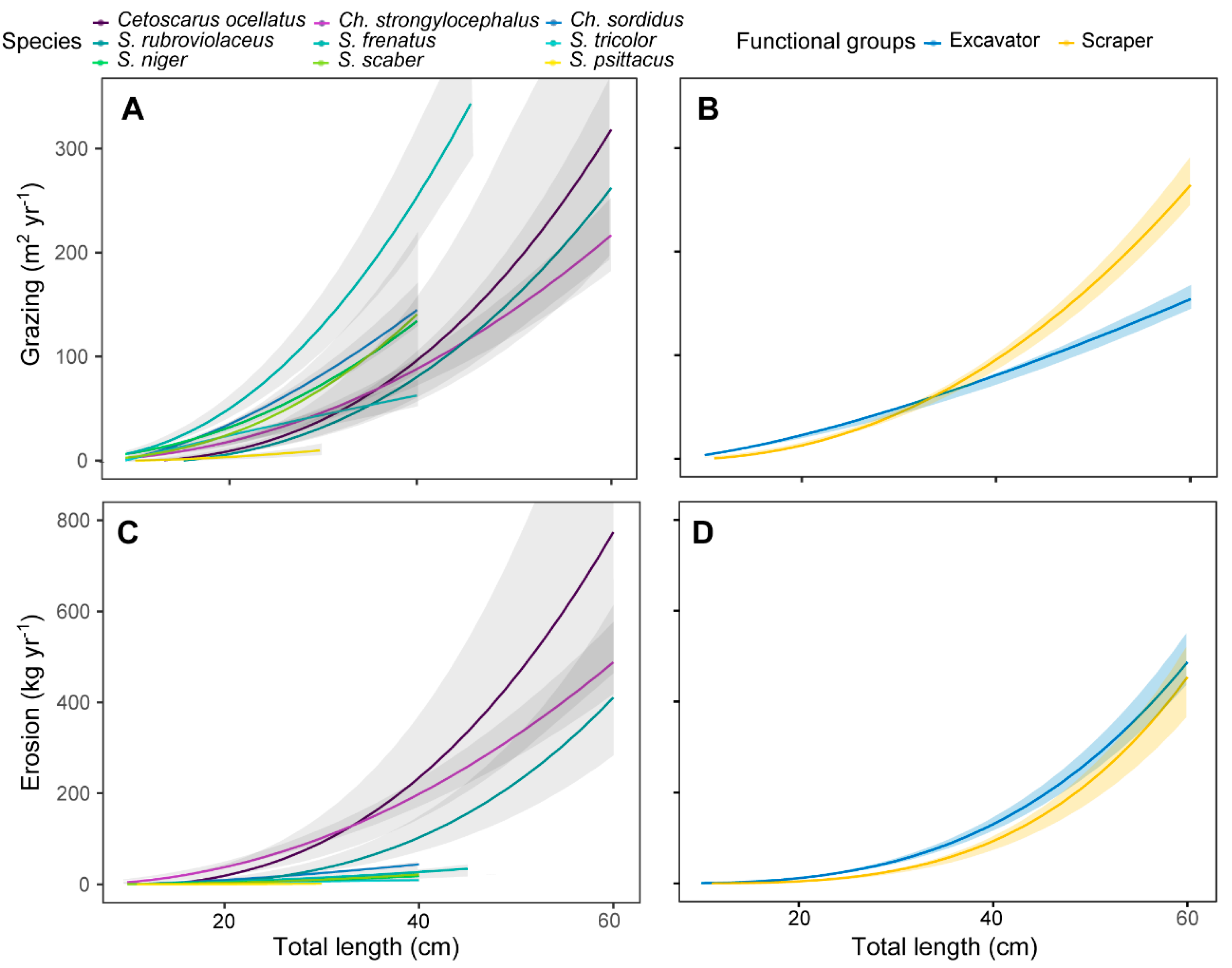
3.2. Individual Grazing and Bioerosion Rates
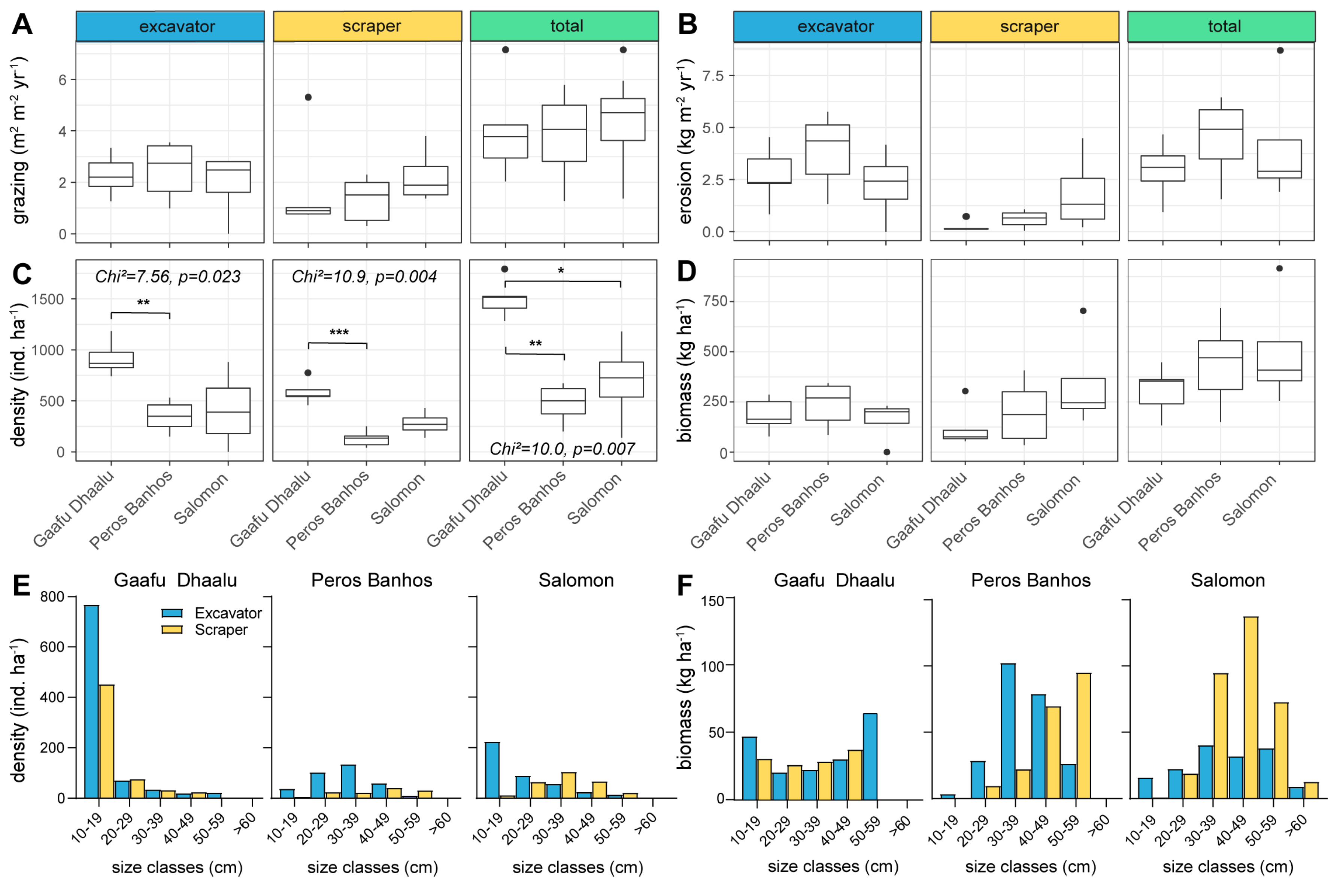
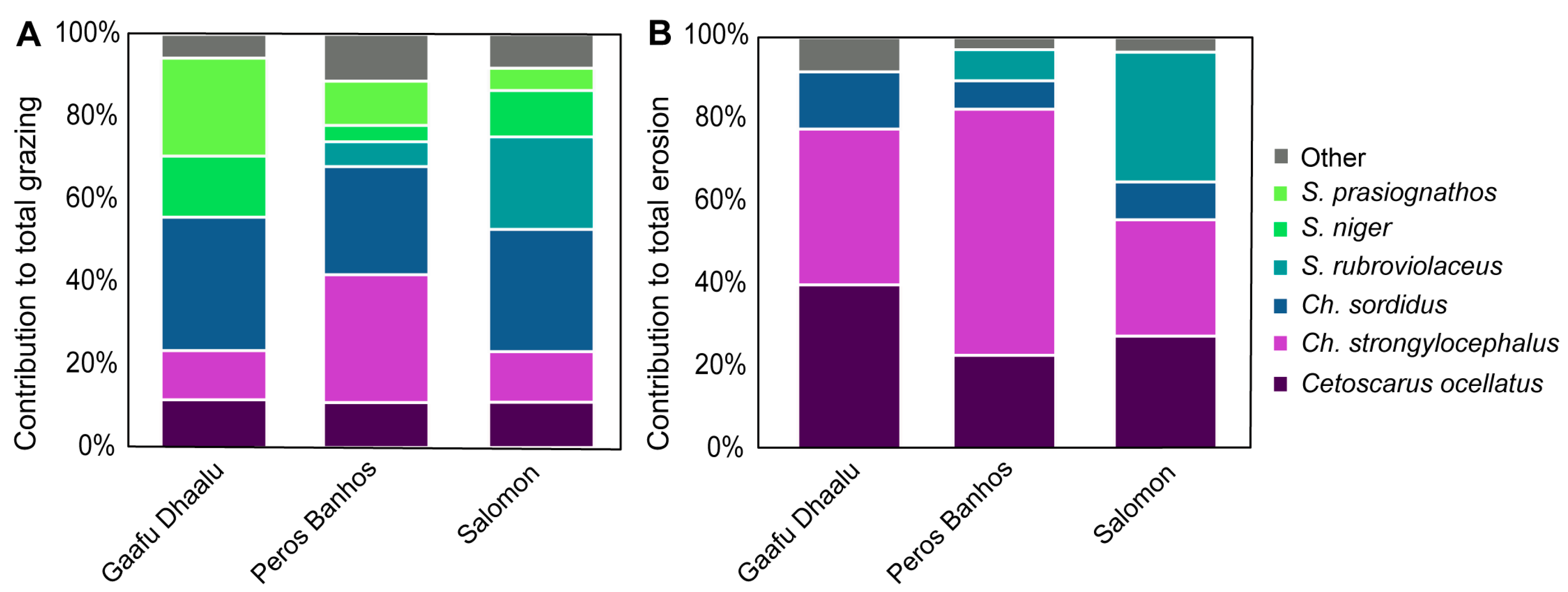
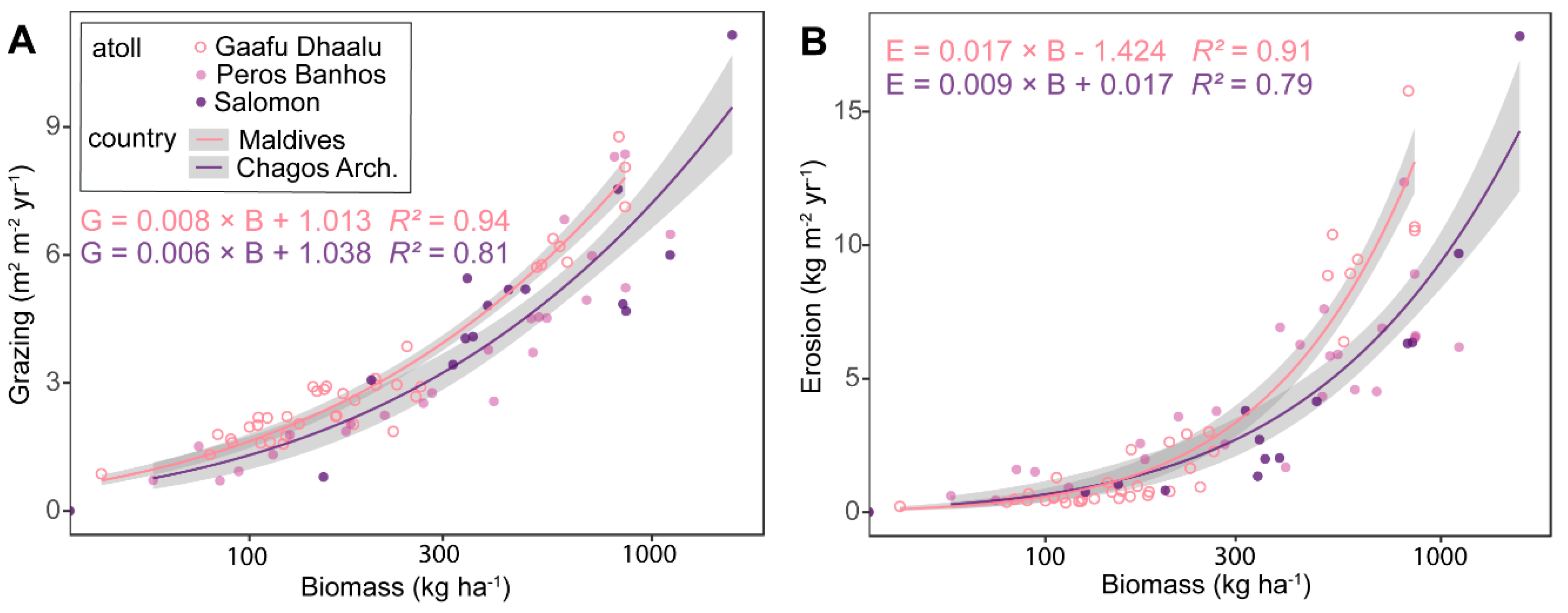
3.3. Community Grazing and Bioerosion Levels
4. Discussion
4.1. Individual Grazing and Bioerosion Rates
4.2. Community Grazing and Bioerosion Levels
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data
References
- Clements, K.D.; German, D.P.; Piché, J.; Tribollet, A.; Choat, J.H. Integrating ecological roles and trophic diversification on coral reefs: Multiple lines of evidence identify parrotfishes as microphages. Biol. J. Linn. Soc. 2017, 120, 729–751. [Google Scholar] [CrossRef]
- Clements, K.D.; Choat, J.H. Nutritional ecology of parrotfishes (Scarinae, Labridae). In Biology of Parrotfishes; Hoey, A.S., Bonaldo, R.M., Eds.; Taylor and Francis: Boca Raton, FL, USA, 2018; pp. 42–68. [Google Scholar]
- Bonaldo, R.M.; Hoey, A.S.; Bellwood, D.R. The ecosystem roles of parrotfishes on tropical reefs. Oceanogr. Mar. Biol. Annu. Rev. 2014, 52, 81–132. [Google Scholar]
- Bellwood, D.R.; Choat, J.H. A functional analysis of grazing in parrotfishes (family scaridae)—The ecological implications. Environ. Biol. Fish 1990, 28, 189–214. [Google Scholar] [CrossRef]
- Bonaldo, R.M.; Bellwood, D.R. Size-dependent variation in the functional role of the parrotfish Scarus rivulatus on the Great Barrier Reef, Australia. Mar. Ecol. Prog. Ser. 2008, 360, 237–244. [Google Scholar] [CrossRef]
- Bruggemann, J.H.; Kuyper, M.W.M.; Breeman, A.M. Comparative analysis of foraging and habitat use by the sympatric Caribbean parrotfish Scarus vetula and Sparisoma viride (Scaridae). Mar. Ecol. Prog. Ser. 1994, 112, 51–66. [Google Scholar] [CrossRef]
- Bellwood, D.R.; Streit, R.P.; Brandl, S.J.; Tebbett, S.B. The meaning of the term ‘function’in ecology: A coral reef perspective. Funct. Ecol. 2019, 33, 948–961. [Google Scholar]
- Williams, G.J.; Graham, N.A. Rethinking coral reef functional futures. Funct. Ecol. 2019, 33, 942–947. [Google Scholar] [CrossRef]
- Hoey, A.S.; Bellwood, D.R. Cross-shelf variation in the role of parrotfishes on the Great Barrier Reef. Coral Reefs 2008, 27, 37–47. [Google Scholar] [CrossRef]
- Robinson, J.P.; McDevitt-Irwin, J.M.; Dajka, J.C.; Hadj-Hammou, J.; Howlett, S.; Graba-Landry, A.; Hoey, A.S.; Nash, K.L.; Wilson, S.K.; Graham, N.A. Habitat and fishing control grazing potential on coral reefs. Funct. Ecol. 2020, 34, 240–251. [Google Scholar] [CrossRef]
- Bellwood, D.R.; Hoey, A.S.; Hughes, T.P. Human activity selectively impacts the ecosystem roles of parrotfishes on coral reefs. Proc. R. Soc. B Biol. Sci. 2012, 279, 1621–1629. [Google Scholar] [CrossRef] [Green Version]
- Graham, N.A.; Wilson, S.K.; Carr, P.; Hoey, A.S.; Jennings, S.; MacNeil, M.A. Seabirds enhance coral reef productivity and functioning in the absence of invasive rats. Nature 2018, 559, 250–253. [Google Scholar] [CrossRef] [Green Version]
- Perry, C.T.; Morgan, K.M.; Lange, I.D.; Yarlett, R.T. Bleaching-driven reef community shifts drive pulses of increased reef sediment generation. R. Soc. Open Sci. 2020, 7, 192153. [Google Scholar] [CrossRef] [Green Version]
- Steneck, R.S.; Mumby, P.J.; MacDonald, C.; Rasher, D.B.; Stoyle, G. Attenuating effects of ecosystem management on coral reefs. Sci. Adv. 2018, 4, eaao5493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, I.; Perry, C.; Alvarez-Filip, L. Carbonate budgets as indicators of functional reef “health”: A critical review of data underpinning census-based methods and current knowledge gaps. Ecol. Indic. 2020, 110, 105857. [Google Scholar] [CrossRef]
- Lokrantz, J.; Nyström, M.; Thyresson, M.; Johansson, C. The non-linear relationship between body size and function in parrotfishes. Coral Reefs 2008, 27, 967–974. [Google Scholar] [CrossRef]
- Ong, L.; Holland, K.N. Bioerosion of coral reefs by two Hawaiian parrotfishes: Species, size differences and fishery implications. Mar. Biol. 2010, 157, 1313–1323. [Google Scholar] [CrossRef]
- Afeworki, Y.; Bruggemann, J.H.; Videler, J.J. Limited flexibility in resource use in a coral reef grazer foraging on seasonally changing algal communities. Coral Reefs 2011, 30, 109–122. [Google Scholar] [CrossRef] [Green Version]
- Morgan, K.M.; Kench, P.S. Parrotfish erosion underpins reef growth, sand talus development and island building in the Maldives. Sediment. Geol. 2016, 341, 50–57. [Google Scholar] [CrossRef]
- Yarlett, R.T.; Perry, C.T.; Wilson, R.W.; Philpot, K.E. Constraining species-size class variability in rates of parrotfish bioerosion on Maldivian coral reefs: Implications for regional-scale bioerosion estimates. Mar. Ecol. Prog. Ser. 2018, 590, 155–169. [Google Scholar] [CrossRef]
- Hoey, A.S. Feeding in parrotfishes: The influence of species, body size, and temperature. In Biology of Parrotfishes; Hoey, A.S., Bonaldo, R.M., Eds.; Taylor and Francis: Boca Raton, FL, USA, 2018; pp. 119–133. [Google Scholar]
- Ferreira, D.E.L.; Peret, A.C.; Coutinho, R. Seasonal grazing rates and food processing by tropical herbivorous fishes. J. Fish Biol. 1998, 53, 222–235. [Google Scholar] [CrossRef]
- Afeworki, Y.; Zekeria, Z.A.; Videler, J.J.; Bruggemann, J.H. Food intake by the parrotfish Scarus ferrugineus varies seasonally and is determined by temperature, size and territoriality. Mar. Ecol. Prog. Ser. 2013, 489, 213–224. [Google Scholar] [CrossRef]
- Bejarano, S.; Jouffray, J.; Chollett, I.; Allen, R.; Roff, G.; Marshell, A.; Steneck, R.; Ferse, S.C.A.; Mumby, P.J. The shape of success in a turbulent world: Wave exposure filtering of coral reef herbivory. Funct. Ecol. 2017, 31, 1312–1324. [Google Scholar] [CrossRef]
- Karkarey, R.; Rathod, P.; Arthur, R.; Yadav, S.; Theo, A.; Alcoverro, T. Wave exposure reduces herbivory in post-disturbed reefs by filtering species composition, abundance and behaviour of key fish herbivores. Sci. Rep. 2020, 10, 9854. [Google Scholar] [CrossRef]
- Gordon, S.E.; Goatley, C.H.; Bellwood, D.R. Low-quality sediments deter grazing by the parrotfish Scarus rivulatus on inner-shelf reefs. Coral Reefs 2016, 35, 285–291. [Google Scholar] [CrossRef]
- Moustaka, M.; Langlois, T.J.; McLean, D.; Bond, T.; Fisher, R.; Fearns, P.; Dorji, P.; Evans, R.D. The effects of suspended sediment on coral reef fish assemblages and feeding guilds of north-west Australia. Coral Reefs 2018, 659–673. [Google Scholar] [CrossRef]
- Lange, I.D.; Perry, C.T. Bleaching impacts on carbonate production in the Chagos Archipelago: Influence of functional coral groups on carbonate budget trajectories. Coral Reefs 2019, 38, 619–624. [Google Scholar] [CrossRef] [Green Version]
- Taylor, B.M.; Benkwitt, C.E.; Choat, H.; Clements, K.D.; Graham, N.A.; Meekan, M.G. Synchronous biological feedbacks in parrotfishes associated with pantropical coral bleaching. Glob. Chang. Biol. 2020, 26, 1285–1294. [Google Scholar] [CrossRef]
- Bellwood, D.R. Direct estimate of bioerosion by two parrotfish species, Chlorurus gibbus and C. sordidus, on the Great Barrier Reef, Australia. Mar. Biol. 1995, 121, 419–429. [Google Scholar] [CrossRef]
- Siqueira, A.C.; Bellwood, D.R.; Cowman, P.F. The evolution of traits and functions in herbivorous coral reef fishes through space and time. Proc. R. Soc. B Biol. Sci. 2019, 286, 20182672. [Google Scholar] [CrossRef] [Green Version]
- 32. Sonia, B.; Yimnang, G.; Tebaua, S.; Mumby, P.J. Ecological risk and the exploitation of herbivorous reef fish across Micronesia. Mar. Ecol. Prog. Ser. 2013, 482, 197–215. [Google Scholar]
- Morgan, K.; Kench, P. Skeletal extension and calcification of reef-building corals in the central Indian Ocean. Mar. Environ. Res. 2012, 81, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Samoilys, M.; Roche, R.; Koldewey, H.; Turner, J. Patterns in reef fish assemblages: Insights from the Chagos Archipelago. PLoS ONE 2018, 13, e0191448. [Google Scholar] [CrossRef] [PubMed]
- Froese, R.; Thorson, J.T.; Reyes Jr, R. A Bayesian approach for estimating length-weight relationships in fishes. J. Appl. Ichthyol. 2014, 30, 78–85. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Faraway, J.J. Linear models with R; Texts in Statistical Science; Chatfield, C., Tanner, M., Zidek, J., Eds.; Taylor and Francis e-library, CRC press: Boca Raton, FL, USA, 2006; Volume 63. [Google Scholar]
- Anderson, G. Error propagation by the Monte Carlo method in geochemical calculations. Geochim. Et Cosmochim. Acta 1976, 40, 1533–1538. [Google Scholar] [CrossRef]
- Ripley, B.; Venables, B.; Bates, D.M.; Hornik, K.; Gebhardt, A.; Firth, D.; Ripley, M.B. Package ‘MASS’. Available online: https://cran.r-project.org/web/packages/MASS/MASS (accessed on 6 September 2020).
- Field, A.; Miles, J.; Field, Z. Discovering Statistics Using R; Sage publications: New York, NY, USA, 2012. [Google Scholar]
- Bruggemann, J.H.; vanKessel, A.M.; van Rooij, J.M.; Breeman, A.M. Bioerosion and sediment ingestion by the Caribbean parrotfish Scarus vetula and Sparisoma viride: Implications of fish size, feeding mode and habitat use. Mar. Ecol. Prog. Ser. 1996, 134, 59–71. [Google Scholar] [CrossRef]
- Adam, T.C.; Duran, A.; Fuchs, C.E.; Roycroft, M.V.; Rojas, M.C.; Ruttenberg, B.I.; Burkepile, D.E. Comparative analysis of foraging behavior and bite mechanics reveals complex functional diversity among Caribbean parrotfishes. Mar. Ecol. Prog. Ser. 2018, 597, 207–220. [Google Scholar] [CrossRef]
- Alwany, M.A.; Thaler, E.; Stachowitsch, M. Parrotfish bioerosion on Egyptian Red Sea reefs. J. Exp. Mar. Biol. Ecol. 2009, 371, 170–176. [Google Scholar] [CrossRef]
- Nicholson, G.M.; Clements, K.D. Resolving resource partitioning in parrotfishes (Scarini) using microhistology of feeding substrata. Coral Reefs 2020. [Google Scholar] [CrossRef]
- Bellwood, D.R. A phylogenetic study of the parrotfish family Scaridae (Pisces: Labroidea), with a revision of genera. Rec. Aust. Mus.Suppl. 1994, 20, 1–86. [Google Scholar] [CrossRef]
- Heenan, A.; Hoey, A.S.; Williams, G.J.; Williams, I.D. Natural bounds on herbivorous coral reef fishes. Proc. R. Soc. B Biol. Sci. 2016, 283, 20161716. [Google Scholar] [CrossRef]
- Adam, T.C.; Schmitt, R.J.; Holbrook, S.J.; Brooks, A.J.; Edmunds, P.J.; Carpenter, R.C.; Bernardi, G. Herbivory, connectivity, and ecosystem resilience: Response of a coral reef to a large-scale perturbation. PLoS ONE 2011, 6, e23717. [Google Scholar] [CrossRef] [PubMed]
- Gilmour, J.P.; Smith, L.D.; Heyward, A.J.; Baird, A.H.; Pratchett, M.S. Recovery of an isolated coral reef system following severe disturbance. Science 2013, 340, 69–71. [Google Scholar] [CrossRef] [PubMed]
- Taylor, B.M.; Trip, E.D.; Choat, J.H. Dynamic demography: Investigations of life-history variation in the parrotfishes. In Biology of Parrotfishes; Hoey, A.S., Bonaldo, R.M., Eds.; Taylor and Francis: Boca Raton, FL, USA, 2018; pp. 69–98. [Google Scholar]
- Madin, E.M.; Gaines, S.D.; Warner, R.R. Field evidence for pervasive indirect effects of fishing on prey foraging behavior. Ecology 2010, 91, 3563–3571. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.; Abdulla, A.; Bertz, N.; Mawyer, A. King tuna: Indian Ocean trade, offshore fishing, and coral reef resilience in the Maldives archipelago. ICES J. Mar. Sci. 2019, 77, 398–407. [Google Scholar] [CrossRef]
- Taylor, B.M.; Lindfield, S.J.; Choat, J.H. Hierarchical and scale-dependent effects of fishing pressure and environment on the structure and size distribution of parrotfish communities. Ecography 2015, 38, 520–530. [Google Scholar] [CrossRef]
- Kuempel, C.D.; Altieri, A.H. The emergent role of small-bodied herbivores in pre-empting phase shifts on degraded coral reefs. Sci. Rep.-UK 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Shantz, A.A.; Ladd, M.C.; Burkepile, D.E. Overfishing and the ecological impacts of extirpating large parrotfish from Caribbean coral reefs. Ecol. Monogr. 2020, 90, e01403. [Google Scholar] [CrossRef]
- Hoey, A.S.; Feary, D.A.; Burt, J.A.; Vaughan, G.; Pratchett, M.S.; Berumen, M.L. Regional variation in the structure and function of parrotfishes on Arabian reefs. Mar. Pollut. Bull. 2016, 105, 524–531. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grazing (m2 ind.−1 yr−1) | 10–19 cm | 20–29 cm | 30–39 cm | 40–49 cm | 50–59 cm | ≥60 cm | a | b |
|---|---|---|---|---|---|---|---|---|
| Cetoscarus ocellatus | 6.9 | 21.6 | 69.4 | 151.7 | 274.1 | 441.5 | 0.0010 | 3.130 |
| Chlorurus strongylocephalus | 7.7 | 29.2 | 64.9 | 114.9 | 179.2 | 245.5 | 0.0209 | 2.256 |
| Chlorurus sordidus | 13.8 | 54.8 | 111.5 | 0.0433 | 2.209 | |||
| Scarus rubroviolaceus | 4.6 | 15.3 | 52.5 | 114.9 | 205.5 | 318.7 | 0.0011 | 3.028 |
| Scarus frenatus | 20.0 | 81.9 | 188.3 | 341.1 | 0.0318 | 2.439 | ||
| Scarus tricolor | 11.6 | 33.2 | 53.1 | 0.1836 | 1.591 | |||
| Scarus niger | 15.6 | 48.4 | 100.6 | 0.0367 | 2.200 | |||
| Scarus scaber | 10.1 | 42.1 | 100.5 | 0.0116 | 2.541 | |||
| Scarus psittacus | 1.0 | 6.0 | 0.0004 | 2.986 | ||||
| Bioerosion (kg ind.−1 yr−1) | 10–19 cm | 20–29 cm | 30–39 cm | 40–49 cm | 50–59 cm | ≥60 cm | a | b |
| Cetoscarus ocellatus | 15.7 | 48.8 | 157.2 | 343.5 | 620.7 | 999.6 | 0.0024 | 3.111 |
| Chlorurus strongylocephalus | 17.4 | 65.8 | 146.2 | 258.7 | 403.5 | 552.9 | 0.0475 | 2.577 |
| Chlorurus sordidus | 4.2 | 16.7 | 33.9 | 0.0131 | 2.211 | |||
| Scarus rubroviolaceus | 3.6 | 15.5 | 62.9 | 156.0 | 308.2 | 519.4 | 0.0003 | 3.488 |
| Scarus frenatus | 3.0 | 12.4 | 28.5 | 51.6 | 0.0058 | 2.386 | ||
| Scarus tricolor | 1.8 | 5.0 | 8.1 | 0.0294 | 1.576 | |||
| Scarus niger | 2.1 | 6.6 | 13.8 | 0.0059 | 2.183 | |||
| Scarus scaber | 1.6 | 6.5 | 15.5 | 0.0019 | 2.517 | |||
| Scarus psittacus | 0.2 | 1.0 | 0.0001 | 2.927 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lange, I.D.; Perry, C.T.; Morgan, K.M.; Roche, R.; Benkwitt, C.E.; Graham, N.A. Site-Level Variation in Parrotfish Grazing and Bioerosion as a Function of Species-Specific Feeding Metrics. Diversity 2020, 12, 379. https://doi.org/10.3390/d12100379
Lange ID, Perry CT, Morgan KM, Roche R, Benkwitt CE, Graham NA. Site-Level Variation in Parrotfish Grazing and Bioerosion as a Function of Species-Specific Feeding Metrics. Diversity. 2020; 12(10):379. https://doi.org/10.3390/d12100379
Chicago/Turabian StyleLange, Ines D, Chris T Perry, Kyle M Morgan, Ronan Roche, Cassandra E Benkwitt, and Nicholas AJ Graham. 2020. "Site-Level Variation in Parrotfish Grazing and Bioerosion as a Function of Species-Specific Feeding Metrics" Diversity 12, no. 10: 379. https://doi.org/10.3390/d12100379
APA StyleLange, I. D., Perry, C. T., Morgan, K. M., Roche, R., Benkwitt, C. E., & Graham, N. A. (2020). Site-Level Variation in Parrotfish Grazing and Bioerosion as a Function of Species-Specific Feeding Metrics. Diversity, 12(10), 379. https://doi.org/10.3390/d12100379