An Assessment of the Influence of Host Species, Age, and Thallus Part on Kelp-Associated Diatoms


Abstract
:
1. Introduction
2. Materials and Methods
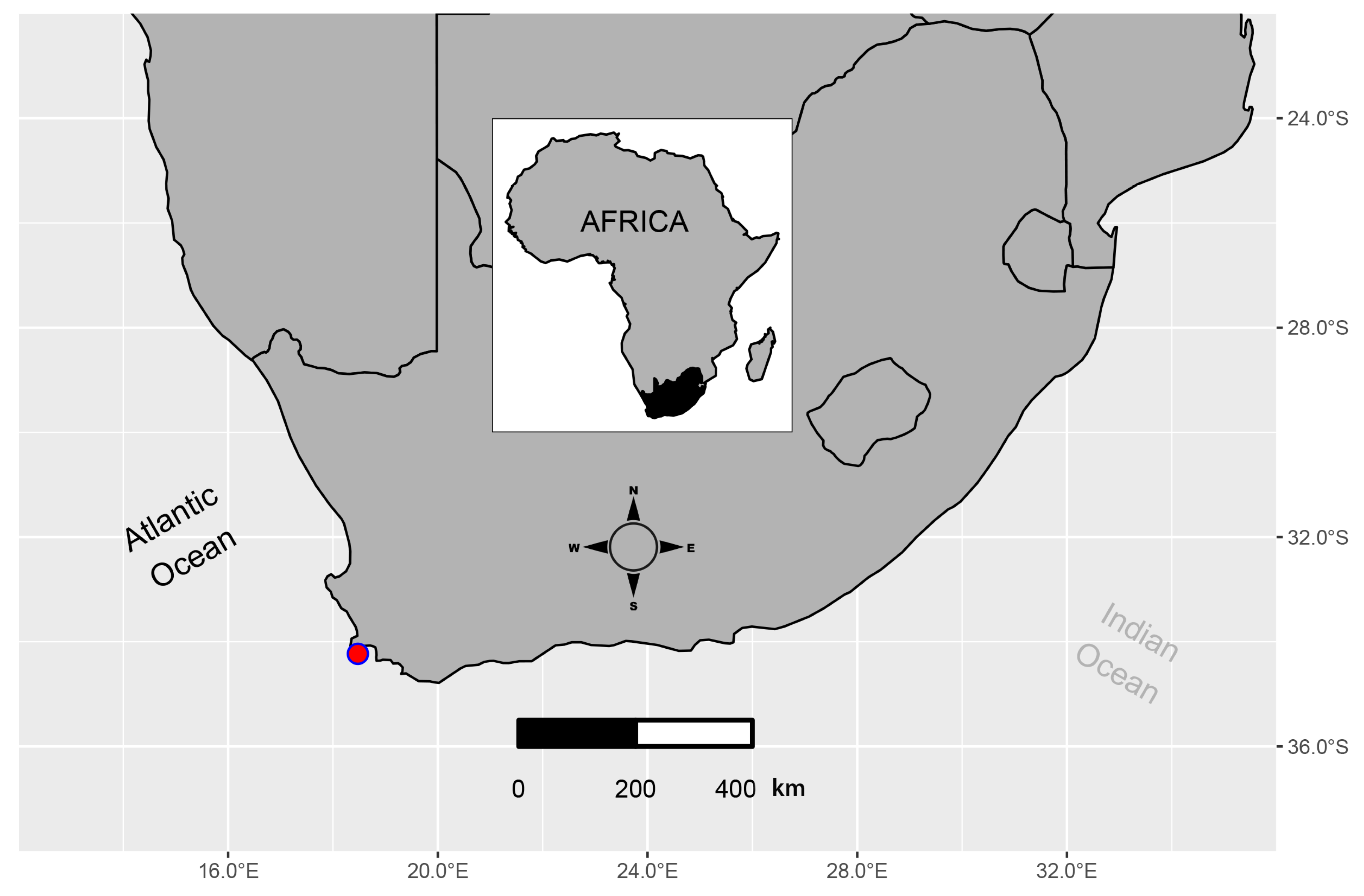
2.1. Study Site
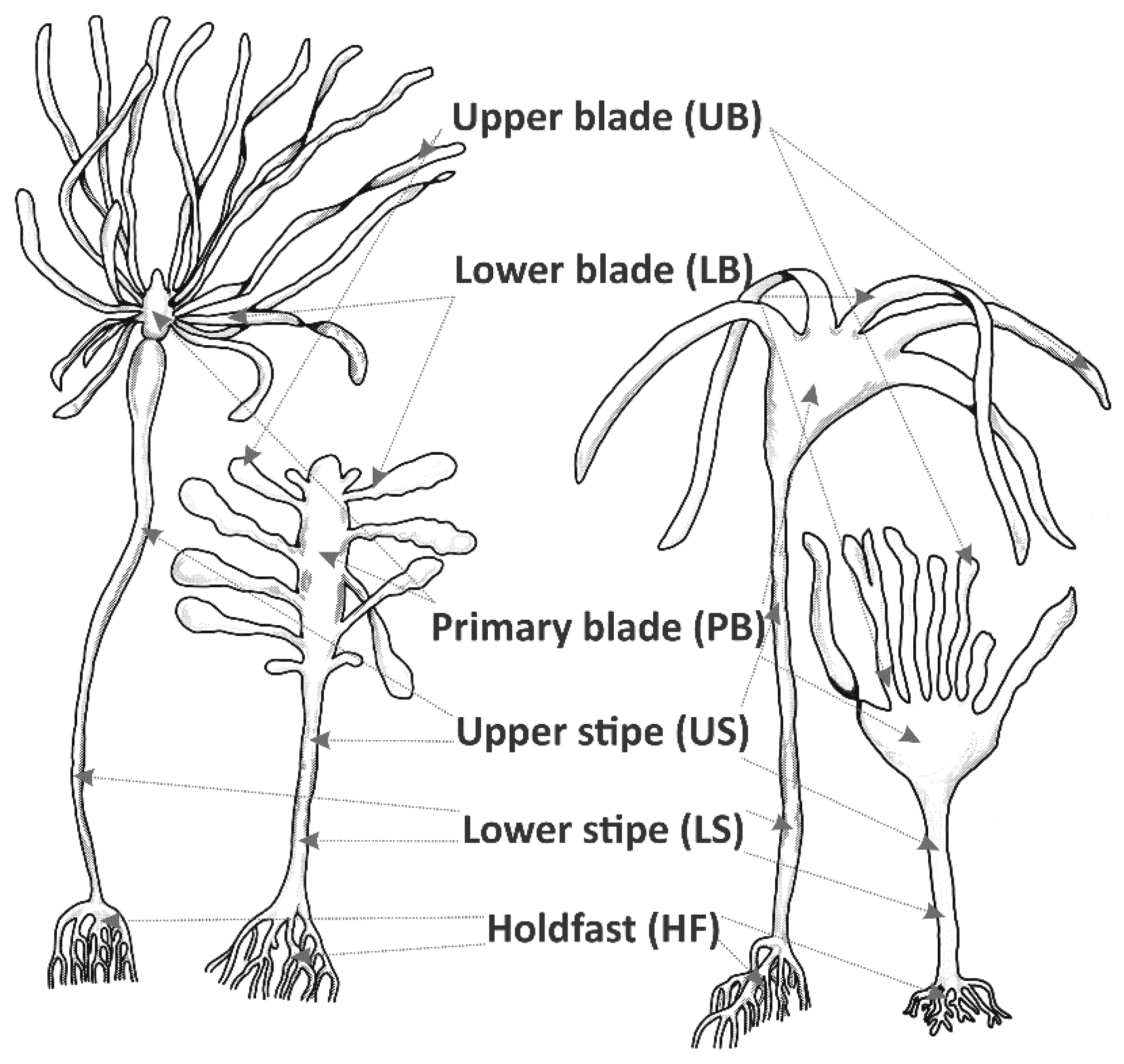
2.2. Material Collection
2.3. Sample Processing and Microscopy
2.4. Statistical Analysis
3. Results
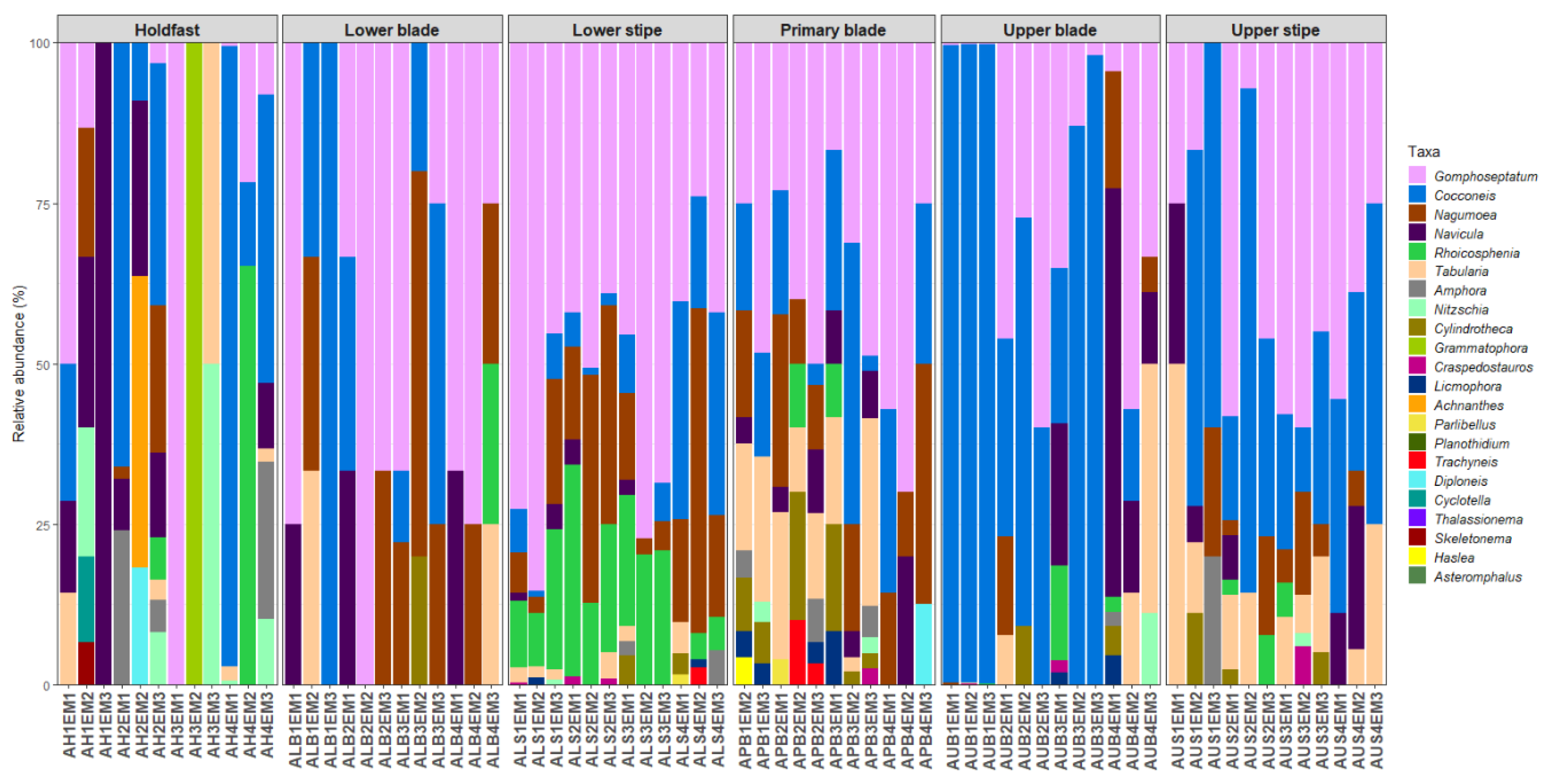
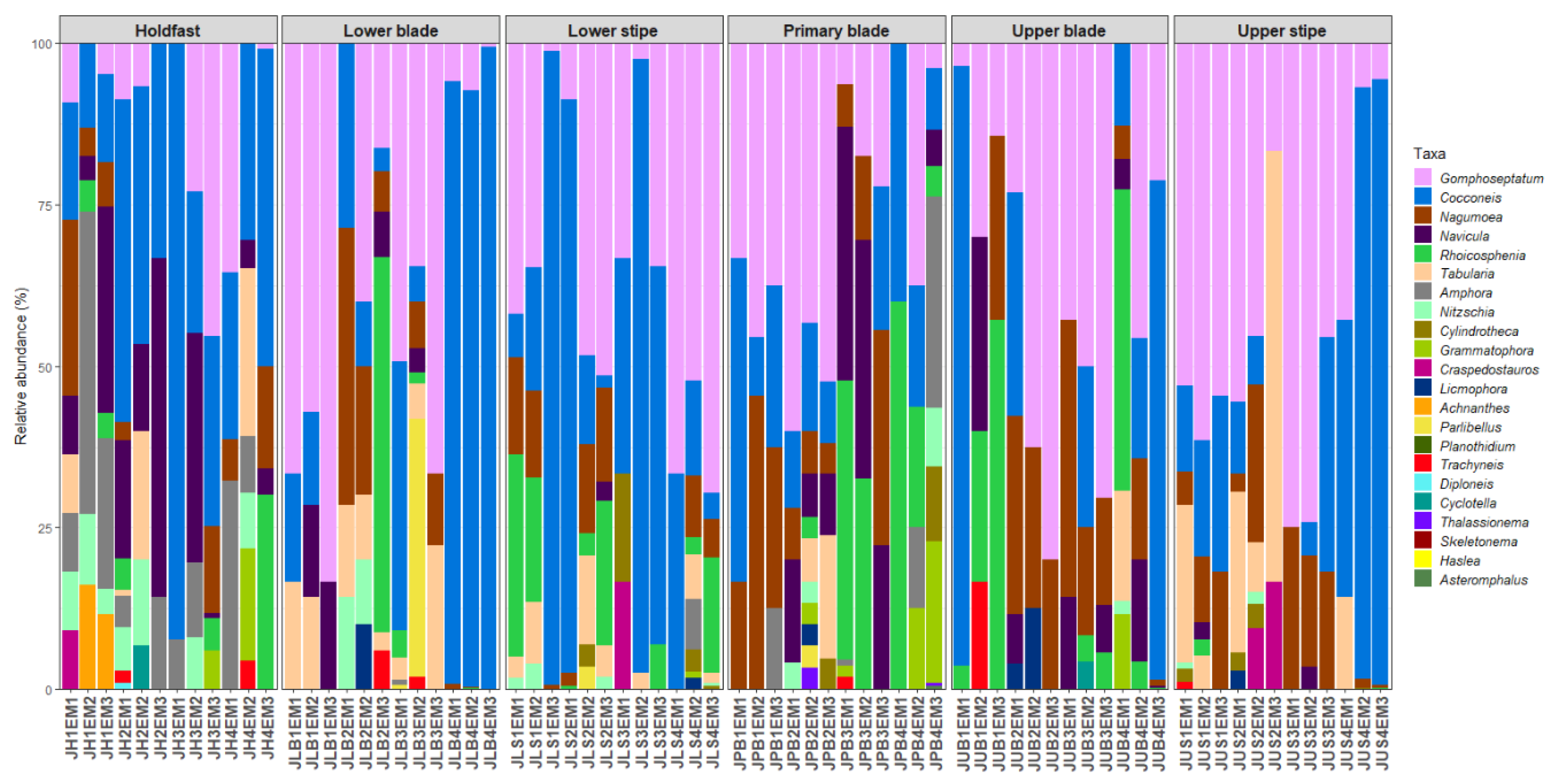
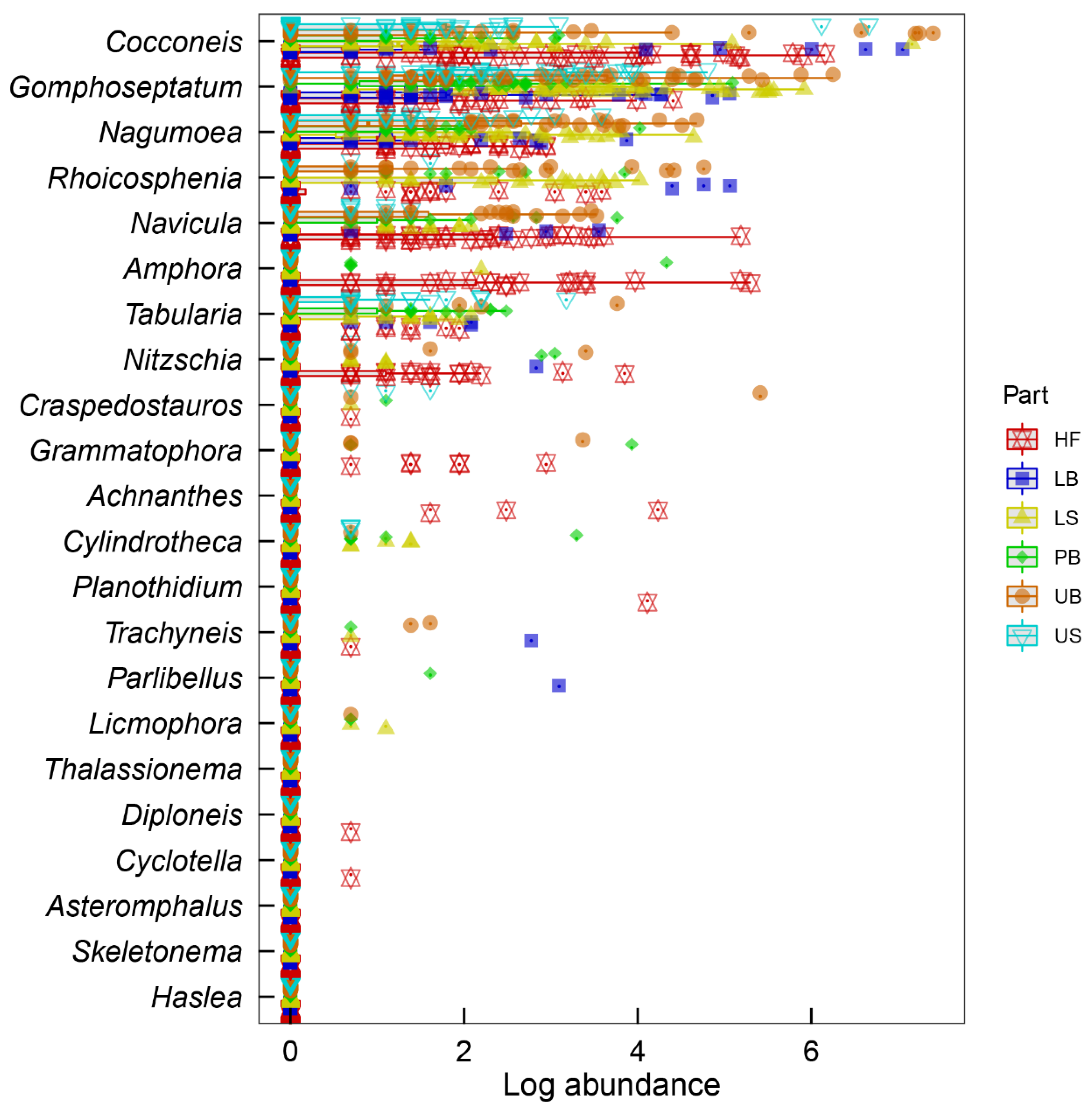
3.1. Diatom Abundances
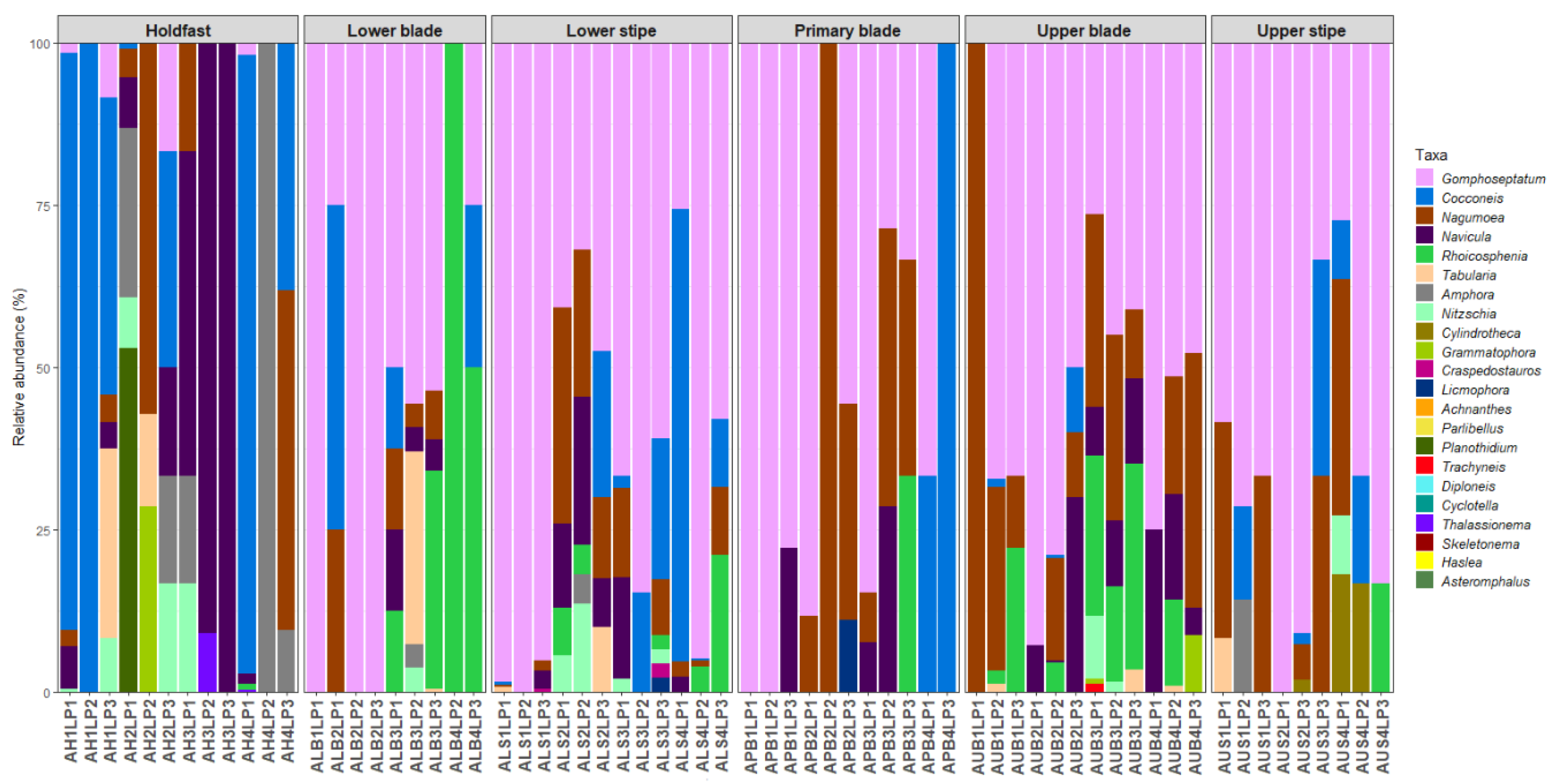
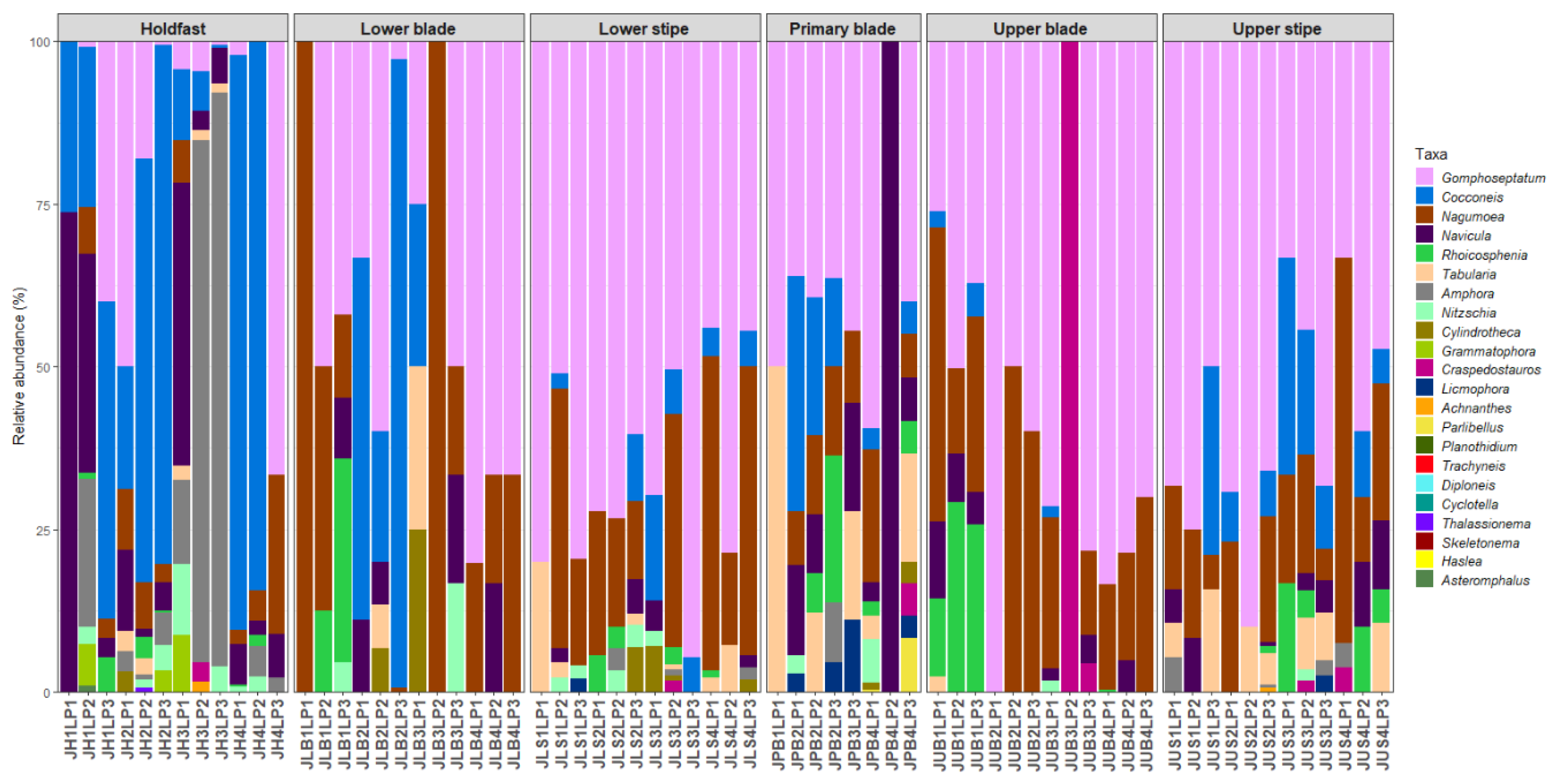
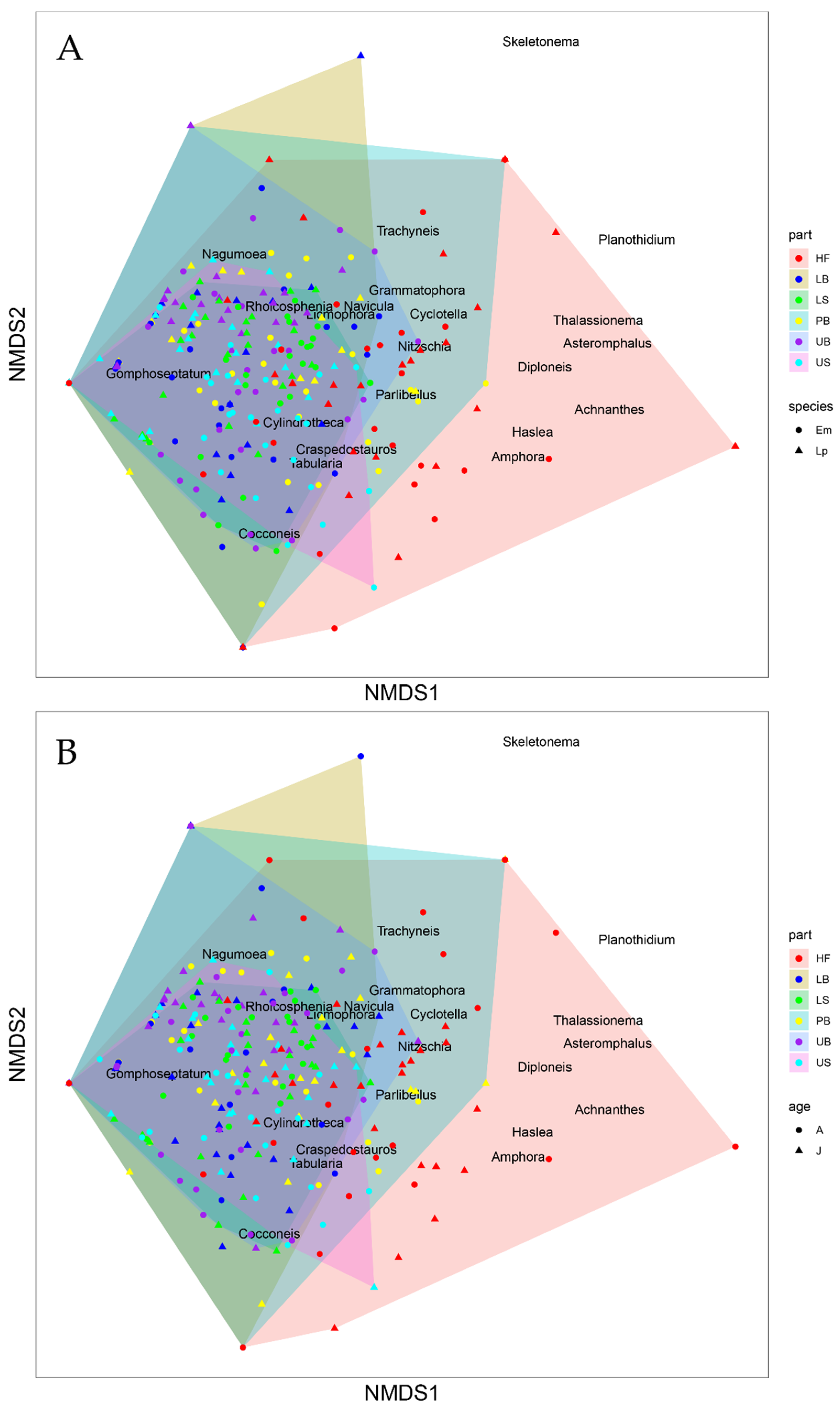
3.2. Diatom Community Composition
3.3. Host Kelp Species, Age, and Thallus Part Effects on Diatom Communities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Krumhansl, K.A.; Okamoto, D.K.; Rassweiler, A.; Novak, M.; Bolton, J.J.; Cavanaugh, K.C.; Connell, S.D.; Johnson, C.R.; Konar, B.; Ling, S.D.; et al. Global patterns of kelp forest change over the past half-century. Proc. Natl. Acad. Sci. USA 2016, 113, 13785–13790. [Google Scholar] [CrossRef] [Green Version]
- Arnold, M.; Teagle, H.; Brown, M.P.; Smale, D.A. The structure of biogenic habitat and epibiotic assemblages associated with the global invasive kelp Undaria pinnatifida in comparison to native macroalgae. Biol. Invasions 2016, 18, 661–676. [Google Scholar] [CrossRef]
- Leclerc, J.C.; Riera, P.; Lévêque, L.; Davoult, D. Contrasting temporal variation in habitat complexity and species abundance distributions in four kelp forest strata. Hydrobiologia 2016, 777, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Booth, W.E. Contribution by diatoms to marine algal host-epiphyte photosynthesis. Bot. Mar. 1987, 30, 129–140. [Google Scholar] [CrossRef]
- Snoeijs, P. Distribution of epiphytic diatom species composition, diversity and biomass on different macroalgal hosts along seasonal and salinity gradients in the Baltic Sea. Diatom Res. 1994, 9, 189–211. [Google Scholar] [CrossRef]
- Siqueiros-Beltrones, D.A.; Serviere-Zaragoza, E.; Argumedo-Hernandez, U. Epiphytic diatoms of Macrocystis pyrifera (L.) C. Ag. from the Baja California Peninsula, México. Oceánides 2002, 17, 3–39. [Google Scholar]
- Al-Handal, A.Y.; Wulff, A. Marine epiphytic diatoms from the shallow sublittoral zone in Potter Cove, King George Island, Antarctica. Bot. Mar. 2008, 51, 411–435. [Google Scholar] [CrossRef]
- Totti, C.; Poulin, M.; Romagnoli, T.; Perrone, C.; Pennesi, C.; de Stefano, M. Epiphytic diatom communities on intertidal seaweeds from Iceland. Pol. Biol. 2009, 32, 1681–1691. [Google Scholar] [CrossRef]
- Majewska, R.; Kuklinski, P.; Balazy, P.; Yokoya, N.S.; Martins, A.P.; De Stefano, M. A comparison of epiphytic diatom communities on Plocamium cartilagineum (Plocamiales, Florideophyceae) from two Antarctic areas. Polar Biol. 2015, 38, 189–205. [Google Scholar] [CrossRef]
- Medlin, L.K.; Juggins, S. Multivariate analyses document host specificity, differences in the diatom metaphyton vs. epiphyton, and seasonality that structure the epiphytic diatom community. Estuar. Coast. Shelf Sci. 2018, 213, 314–330. [Google Scholar] [CrossRef] [Green Version]
- Medlin, L.K.; Fryxell, G.A.; Cox, E.R. Successional sequences of microbial colonisation on three species of Rhodophycean macroalgae. Ann. Bot. 1985, 56, 399–413. [Google Scholar] [CrossRef]
- Majewska, R.; Gambi, M.C.; Totti, C.M.; De Stefano, M. Epiphytic diatom communities of Terra Nova Bay, Ross Sea, Antarctica: Structural analysis and relations to algal host. Antarct. Sci. 2013, 25, 501–513. [Google Scholar] [CrossRef]
- Costa, M.M.D.S.; Pereira, S.M.B.; Da Silva-Cunha, M.D.G.G.; De Arruda, P.C.; Eskinazi-Leça, E. Community structure of epiphytic diatoms on seaweeds in Northeastern Brazil. Bot. Mar. 2016, 59, 231–240. [Google Scholar] [CrossRef]
- Majewska, R.; Convey, P.; De Stefano, M. Summer epiphytic diatoms from Terra Nova Bay and Cape Evans (Ross Sea, Antarctica)—A synthesis and final conclusions. PLoS ONE 2016, 11, e0153254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, M.M.D.S.; Pereira, S.M.B.; De Arruda, P.C.; Leca, E.E. Quantitative variation of epiphytic diatoms in Galaxaura rugosa (Nemaliales: Rhodophyta). Mar. Biodiv. Rec. 2014, 7, 1–7. [Google Scholar] [CrossRef]
- Chung, M.H.; Lee, K.S. Species composition of the epiphytic diatoms on the leaf tissues of three Zostera species distributed on the southern coast of Korea. Algae 2008, 23, 75–81. [Google Scholar] [CrossRef]
- Jennings, J.G.; Steinberg, P.D. Phlorotannins versus other factors affecting epiphyte abundance on the kelp Ecklonia radiata. Oecologia 1997, 109, 461–473. [Google Scholar] [CrossRef]
- Moss, B.L. The control of epiphytes by Halidrys siliquosa (L.) Lyngb. (Phaeophyta, Cystoseiraceae). Phycologia 1982, 21, 185–191. [Google Scholar] [CrossRef]
- Russell, G.; Veltkamp, C.J. Epiphyte survival on skin-shedding macrophytes. Mar. Ecol. Prog. Ser. 1984, 18, 149–153. [Google Scholar] [CrossRef]
- Nylund, G.M.; Pavia, H. Chemical versus mechanical inhibition of fouling in the red alga Dilsea carnosa. Mar. Ecol. Prog. Ser. 2005, 299, 111–121. [Google Scholar] [CrossRef]
- Halat, L.; Galway, M.E.; Gitto, S.; Garbary, D.J. Epidermal shedding in Ascophyllum nodosum (Phaeophyceae): Seasonality, productivity and relationship to harvesting. Phycologia 2015, 54, 599–608. [Google Scholar] [CrossRef]
- Yamamoto, K.; Endo, H.; Yoshikawa, S.; Ohkia, K.; Kamiya, M. Various defense ability of four sargassacean algae against the red algal epiphyte Neosiphonia harveyi in Wakasa Bay, Japan. Aquat. Bot. 2013, 105, 11–17. [Google Scholar] [CrossRef]
- Mayombo, N.A.S.; Majewska, R.; Smit, A.J. Diatoms associated with two South African kelp species: Ecklonia maxima and Laminaria pallida. Afr. J. Mar. Sci. 2019, 41, 221–229. [Google Scholar] [CrossRef]
- Russell, G. Formation of an ectocarpoid epiflora on blades of Laminaria digitata. Mar. Ecol. Prog. Ser. 1983, 11, 181–187. [Google Scholar] [CrossRef]
- Russell, G. Parallel growth patterns in algal epiphytes and Laminaria blades. Mar. Ecol. Prog. Ser. 1983, 13, 149–153. [Google Scholar] [CrossRef]
- Amsler, C.D.; Okogbue, I.N.; Landry, D.M.; Amsler, M.O.; McClintock, J.B.; Baker, B.J. Potential chemical defences against diatom fouling in Antarctic macroalgae. Bot. Mar. 2005, 48, 318–322. [Google Scholar] [CrossRef]
- Cundell, A.M.; Sleeter, T.D.; Mitchell, R. Microbial Populations Associated with the Surface of the Brown Alga Ascophyllum nodosum. Microb. Ecol. 1977, 4, 81–91. [Google Scholar] [CrossRef]
- Walsh, G.; Wepener, V. The influence of land use on water quality and diatom community structures in urban and agriculturally stressed rivers. Water SA 2009, 35, 579–594. [Google Scholar] [CrossRef]
- Witon, E.; Witkowski, A. Holocene diatoms (Bacillariophyceae) from Faeroe Islands Fjords, Northern Atlantic Ocean. II. Distribution and taxonomy of marine taxa with special reference to benthic forms. Diatom Res. 2006, 21, 175–215. [Google Scholar] [CrossRef]
- Desianti, N.; Potapova, M.; Enache, M.; Belton, T.J.; Velinsky, D.J.; Thomas, R.; Mead, J. Sediment diatoms as environmental indicators in New Jersey coastal lagoons. J. Coast. Res. 2017, 78, 127–140. [Google Scholar] [CrossRef]
- Giffen, M.H. Marine littoral diatoms from the Gordon’s Bay region of False Bay Cape Province, South Africa. Bot. Mar. 1971, 14, 1–16. [Google Scholar] [CrossRef]
- Giffen, M.H. Diatoms of the marine littoral of Steenberg’s Cove in St. Helena Bay, Cape Province, South Africa. Bot. Mar. 1973, 16, 32–48. [Google Scholar] [CrossRef]
- Giffen, M.H. A further account of the marine littoral diatoms of the Saldanha Bay Lagoon, Cape Province, South Africa. Bot. Mar. 1976, 19, 379–394. [Google Scholar] [CrossRef] [Green Version]
- Bolton, J.J. Marine phytogeography of the Benguela Upwelling Region on the west coast of Southern Africa: A temperature dependent approach. Bot. Mar. 1986, 29, 251–256. [Google Scholar] [CrossRef]
- Brown, A.C.; Jarman, N. Coastal marine habitats. In Biogeography and Ecology of Southern Africa; Werger, M.J.A., Ed.; Monographiae Biologicae, Springer: Dordrecht, The Netherlands, 1978; Volume 31. [Google Scholar]
- Anderson, R.J.; Carrick, P.; Levitt, G.J.; Share, A. Holdfasts of adult kelp Ecklonia maxima provide refuges from grazing for recruitment of juvenile kelps. Mar. Ecol. Prog. Ser. 1997, 159, 265–273. [Google Scholar] [CrossRef]
- Hasle, G.R.; Syvertsen, E.E. Marine diatoms. In Identifying marine phytoplankton; Tomas, C.R., Ed.; Academic Press: San Diego, CA, USA, 1997; pp. 5–385. [Google Scholar]
- Von Stosch, H.A. Pleurax, seine Synthese und seine Verwendung zur Einbettung und Darstellung der Zellwände von Diatomeen, Peridineen und anderen Algen, sowie für eine neue Methode zur Elektivfärbung von Dinoflagellaten-Panzern. Arch. Protistenkd. 1974, 116, 132–141. [Google Scholar]
- Medlin, L.K.; Round, F.E. Taxonomic studies of marine Gomphonemoid diatoms. Diatom Res. 1986, 1, 205–225. [Google Scholar] [CrossRef]
- Witkowski, A.; Kociolek, J.P.; Kurzydlowski, K.J. Valve ultrastructure of two new genera of marine canal-bearing diatoms (Bacillariophyceae). Phycologia 2011, 50, 170–181. [Google Scholar] [CrossRef]
- Lobban, C.S.; Schefter, M.; Jordan, R.W.; Arai, Y.; Sasaki, A.; Theriot, E.C.; Ashworth, M.; Ruck, E.C.; Pennesi, C. Coral-reef diatoms (Bacillariophyta) from Guam: New records and preliminary checklist, with emphasis on epiphytic species from farmer-fish territories. Micronesica 2012, 43, 237–479. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing, 3.5.3; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package; R Package Version 2.5-5; Creative Commons: San Francisco, CA, USA, 2019. [Google Scholar]
- Wang, Y.I.; Naumann, U.; Wright, S.T.; Warton, D.I. mvabund—An R package for model-based analysis of multivariate abundance data. Methods. Ecol. Evol. 2012, 3, 471–474. [Google Scholar] [CrossRef]
- Wang, Y.; Naumann, U.; Eddelbuettel, D.; Warton, D. Mvabund: Statistical Methods for Analysing Multivariate Abundance Data; R Package Version 3.13.1; Creative Commons: San Francisco, CA, USA, 2018. [Google Scholar]
- Anderson, M.J.; Ellingsen, K.E.; McArdle, B.H. Multivariate dispersion as a measure of beta diversity. Ecol. Lett. 2006, 9, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Warton, D.I.; Wright, S.T.; Wang, Y. Distance-based multivariate analyses confound location and dispersion effects. Methods. Ecol. Evol. 2012, 3, 89–101. [Google Scholar] [CrossRef]
- Benesh, D.P.; Kalbe, M. Experimental parasite community ecology: Intraspecific variation in a large tapeworm affects community assembly. J. Anim. Ecol. 2016, 85, 1004–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majewska, R.; Gambi, M.C.; Totti, C.M.; Pennesi, C.; De Stefano, M. Growth form analysis of epiphytic diatom communities of Terra Nova Bay (Ross Sea, Antarctica). Pol. Biol. 2013, 36, 73–86. [Google Scholar] [CrossRef]
- Polifrone, M.; Viera-Rodríguez, M.A.; Pennesi, C.; Conte, M.T.; Del Pino, A.S.; Stroobant, M.; De Stefano, M. Epiphytic diatoms on Gelidiales (Rhodophyta) from Gran Canaria (Spain). Eur. J. Phycol. 2020. [Google Scholar] [CrossRef]
- Siqueiros-Beltrones, D.A.; Argumedo-Hernandez, U.; Landa-Cansigno, C. Uncommon species diversity values in epiphytic diatom assemblages of the kelp Eisenia arborea. Hidrobiológica 2016, 26, 61–76. [Google Scholar] [CrossRef]
- Wuchter, C.; Marquardt, J.; Krumbein, W.E. The epizoic diatom community on four bryozoan species from Helgoland (German Bight, North Sea). Helgol. Mar. Res. 2008, 57, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Steinberg, P.D. Algal chemical defense against herbivores: Allocation of phenolic compounds in the kelp Alaria marginata. Science 1984, 223, 405–407. [Google Scholar] [CrossRef] [PubMed]
- Tugwell, S.; Branch, G.M. Differential polyphenolic distribution among tissues in the kelps Ecklonia maxima, Laminaria pallida and Macrocystis angustifolia in relation to plant-defence theory. J. Exp. Mar. Biol. Ecol. 1989, 129, 219–230. [Google Scholar] [CrossRef]
- Dubois, A.; Iken, K. Seasonal variation in kelp phlorotannins in relation to grazer abundance and environmental variables in the Alaskan sublittoral zone. Algae 2012, 27, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Amsler, C.D.; Amsler, M.O.; Curtis, M.D.; McClintock, J.B.; Baker, B.J. Impacts of gastropods on epiphytic microalgae on the brown macroalga Himantothallus grandifolius. Antarct. Sci. 2019, 31, 89–97. [Google Scholar] [CrossRef]
- Toth, G.B.; Pavia, H. Intraplant habitat and feeding preference of two gastropod herbivores inhabiting the kelp Laminaria hyperborea. J. Mar. Biol. Assoc. UK 2002, 82, 243–247. [Google Scholar] [CrossRef]
- Christie, H.; Jørgensen, N.M.; Norderhaug, K.M. Bushy or smooth, high or low; importance of habitat architecture and vertical position for distribution of fauna on kelp. J. Sea Res. 2007, 58, 198–208. [Google Scholar] [CrossRef]
- Steneck, R.S.; Watling, L. Feeding capabilities and limitation of herbivorous Molluscs: A functional group approach. Mar. Biol. 1982, 68, 299–319. [Google Scholar] [CrossRef]
- Thompson, R.C.; Johnson, L.E.; Hawkins, S.J. A method for spatial and temporal assessment of gastropod grazing intensity in the field: The use of radula scrapes on wax surfaces. J. Exp. Mar. Biol. Ecol. 1997, 218, 63–76. [Google Scholar] [CrossRef]
- Johnson, C.R.; Mann, K.H. The importance of plant defence abilities to the structure of subtidal seaweed communities: The kelp Laminaria longicruris de la Pylaie survives grazing by the snail Lacuna vincta (Montagu) at high population densities. J. Exp. Mar. Biol. Ecol. 1986, 97, 231–267. [Google Scholar] [CrossRef]
- Klochkova, T.A.; Pisareva, N.A.; Park, J.S.; Lee, J.H.; Han, J.W.; Klochkova, N.G.; Kim, G.H. An endophytic diatom, Pseudogomphonema sp. (Naviculaceae, Bacillariophyceae), lives inside the red alga Neoabbottiella (Halymeniaceae, Rhodophyta). Phycologia 2014, 53, 205–214. [Google Scholar] [CrossRef]
- Lewin, J.; Lewin, R.A. Culture and nutrition of some apochlorotic diatoms of the genus Nitzschia. J. Gen. Microbiol. 1967, 46, 361–367. [Google Scholar] [CrossRef] [Green Version]
- Sieburth, J.M.N. Studies on algal substances in the sea. III. The production of extracellular organic matter by littoral marine algae. J. Exp. Mar. Biol. Ecol. 1969, 3, 290–309. [Google Scholar] [CrossRef]
- Armstrong, E.; Rogerson, A.; Leftley, J.W. Utilisation of seaweed carbon by three surface-associated heterotrophic protists, Stereomyxa ramosa, Nitzschia alba and Labyrinthula sp. Aquat. Microb. Ecol. 2000, 21, 49–57. [Google Scholar] [CrossRef]
- Armstrong, E.; Rogerson, A.; Leftley, J.W. The abundance of heterotrophic Protists associated with intertidal seaweeds. Estuar. Coast. Shelf Sci. 2000, 50, 415–424. [Google Scholar] [CrossRef]
- Fierer, N.; Nemergut, D.; Knight, R.; Craine, J.M. Changes through time: Integrating microorganisms into the study of succession. Res. Microb. 2010, 161, 635–642. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substatum | Mean Diatom Abundance | SD | ||
|---|---|---|---|---|
| Ecklonia maxima | Adult | Holdfast | 39 | 51 |
| Lower stipe | 139 | 105 | ||
| Upper stipe | 19 | 14 | ||
| Primary blade | 21 | 15 | ||
| Lower blade | 4 | 2 | ||
| Upper blade | 404 | 647 | ||
| Ecklonia maxima | Juvenile | Holdfast | 104 | 116 |
| Lower stipe | 198 | 400 | ||
| Upper Stipe | 139 | 258 | ||
| Primary blade | 43 | 66 | ||
| Lower blade | 248 | 395 | ||
| Upper Blade | 120 | 263 | ||
| Laminaria pallida | Adult | Holdfast | 73 | 116 |
| Lower stipe | 120 | 130 | ||
| Upper stipe | 9 | 15 | ||
| Primary blade | 7 | 5 | ||
| Lower blade | 24 | 69 | ||
| Upper blade | 110 | 118 | ||
| Laminaria pallida | Juvenile | Holdfast | 187 | 161 |
| Lower stipe | 49 | 32 | ||
| Upper stipe | 41 | 54 | ||
| Primary blade | 38 | 77 | ||
| Lower blade | 54 | 110 | ||
| Upper blade | 155 | 201 | ||
| Holdfast | 100 | 127 | ||
| Lower stipe | 126 | 217 | ||
| Upper stipe | 52 | 138 | ||
| Primary blade | 27 | 52 | ||
| Lower blade | 82 | 224 | ||
| Upper blade | 197 | 376 | ||
| Adult | 80 | 221 | ||
| Juvenile | 114 | 218 | ||
| Ecklonia maxima | 122 | 285 | ||
| Laminaria pallida | 72 | 117 | ||
| Factor | df | SS | R2 | F | p * |
|---|---|---|---|---|---|
| Species | 1 | 1.701 | 0.029 | 9.972 | 0.0001 |
| Age | 1 | 0.730 | 0.012 | 4.278 | 0.0005 |
| Part | 5 | 8.752 | 0.147 | 10.261 | 0.0001 |
| Species × Age | 1 | 0.196 | 0.003 | 1.146 | 0.3248 |
| Species × Part | 5 | 1.377 | 0.023 | 1.615 | 0.0163 |
| Age × Part | 5 | 2.034 | 0.034 | 2.384 | 0.0002 |
| Species × Age × Part | 5 | 1.838 | 0.031 | 2.155 | 0.0003 |
| Residual | 252 | 42.989 | 0.721 | ||
| Total | 275 | 59.617 | 1.000 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mayombo, N.A.S.; Majewska, R.; Smit, A.J. An Assessment of the Influence of Host Species, Age, and Thallus Part on Kelp-Associated Diatoms. Diversity 2020, 12, 385. https://doi.org/10.3390/d12100385
Mayombo NAS, Majewska R, Smit AJ. An Assessment of the Influence of Host Species, Age, and Thallus Part on Kelp-Associated Diatoms. Diversity. 2020; 12(10):385. https://doi.org/10.3390/d12100385
Chicago/Turabian StyleMayombo, Ntambwe Albert Serge, Roksana Majewska, and Albertus J. Smit. 2020. "An Assessment of the Influence of Host Species, Age, and Thallus Part on Kelp-Associated Diatoms" Diversity 12, no. 10: 385. https://doi.org/10.3390/d12100385
APA StyleMayombo, N. A. S., Majewska, R., & Smit, A. J. (2020). An Assessment of the Influence of Host Species, Age, and Thallus Part on Kelp-Associated Diatoms. Diversity, 12(10), 385. https://doi.org/10.3390/d12100385