Genetic Diversity and Population Structure in a Vitis spp. Core Collection Investigated by SNP Markers



Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Material
2.2. DNA Extraction and SNP Genotyping
2.3. Data Analysis
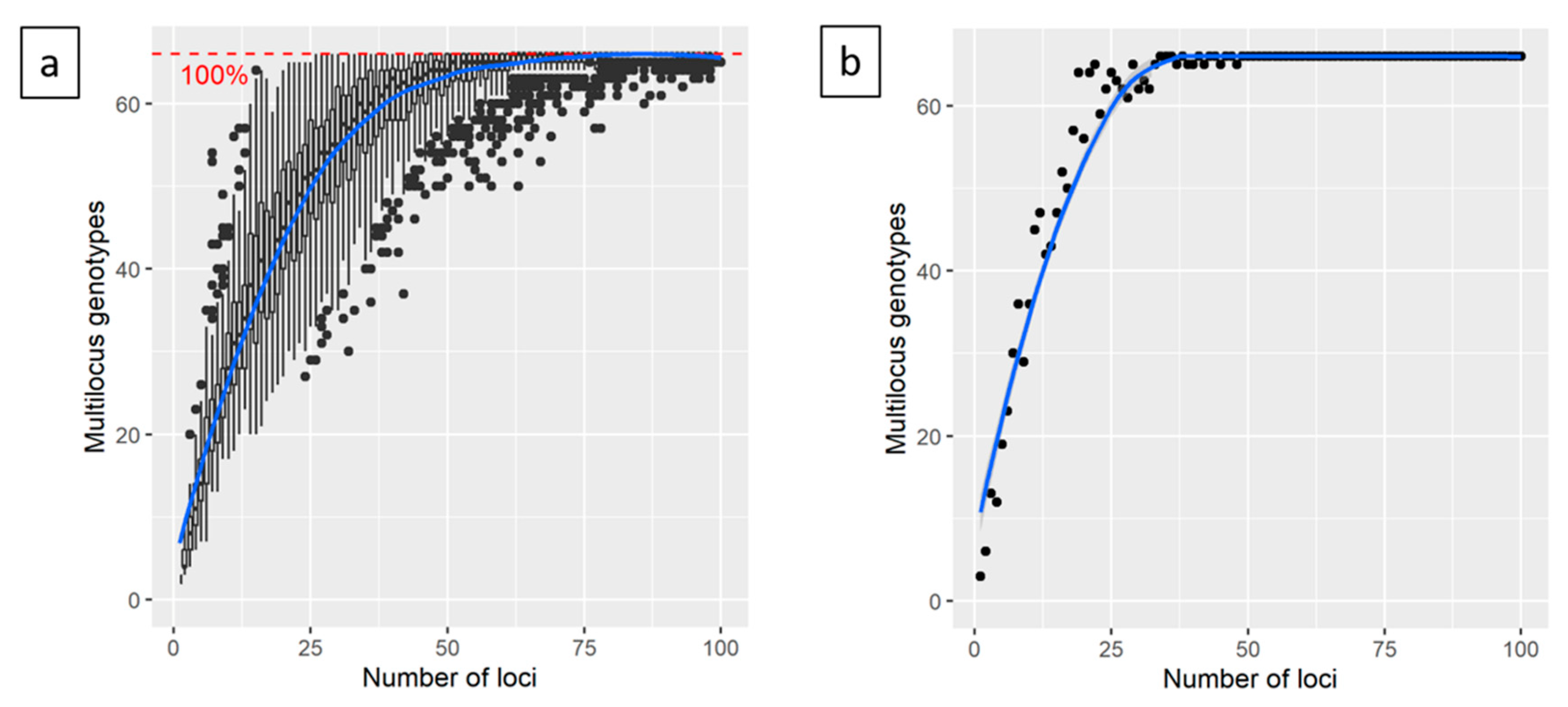
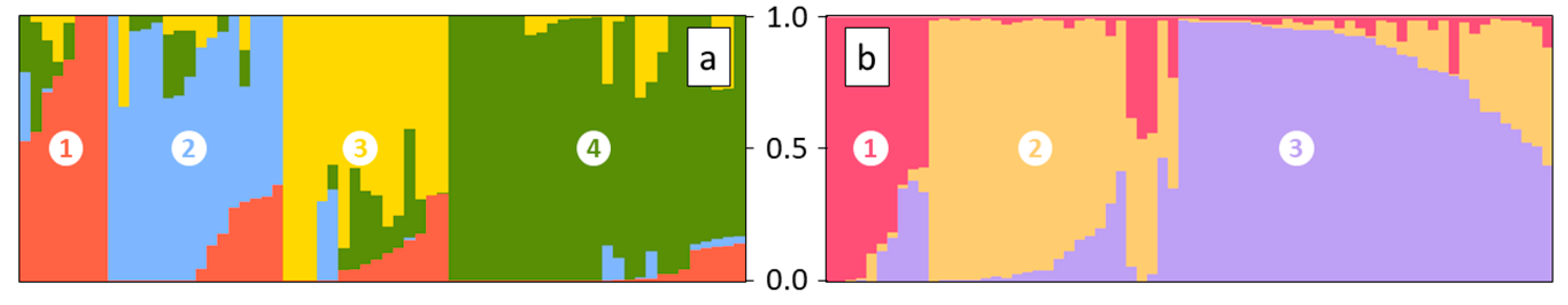
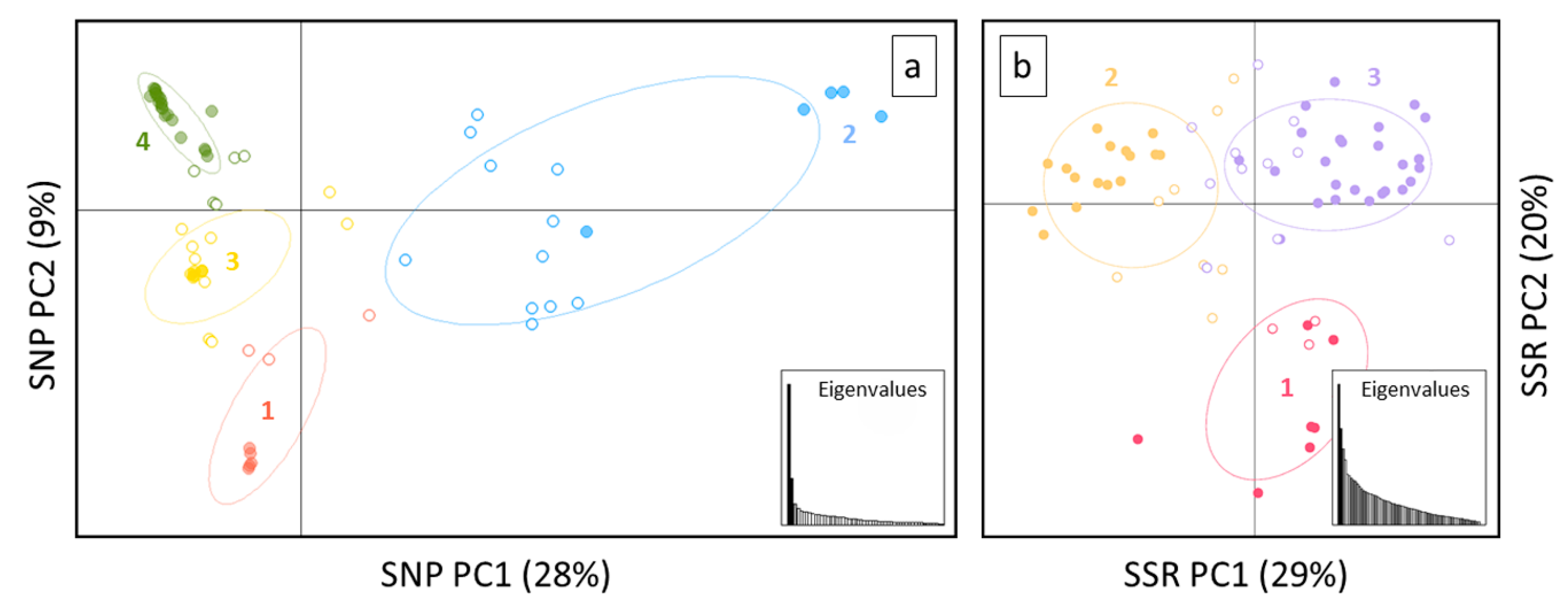
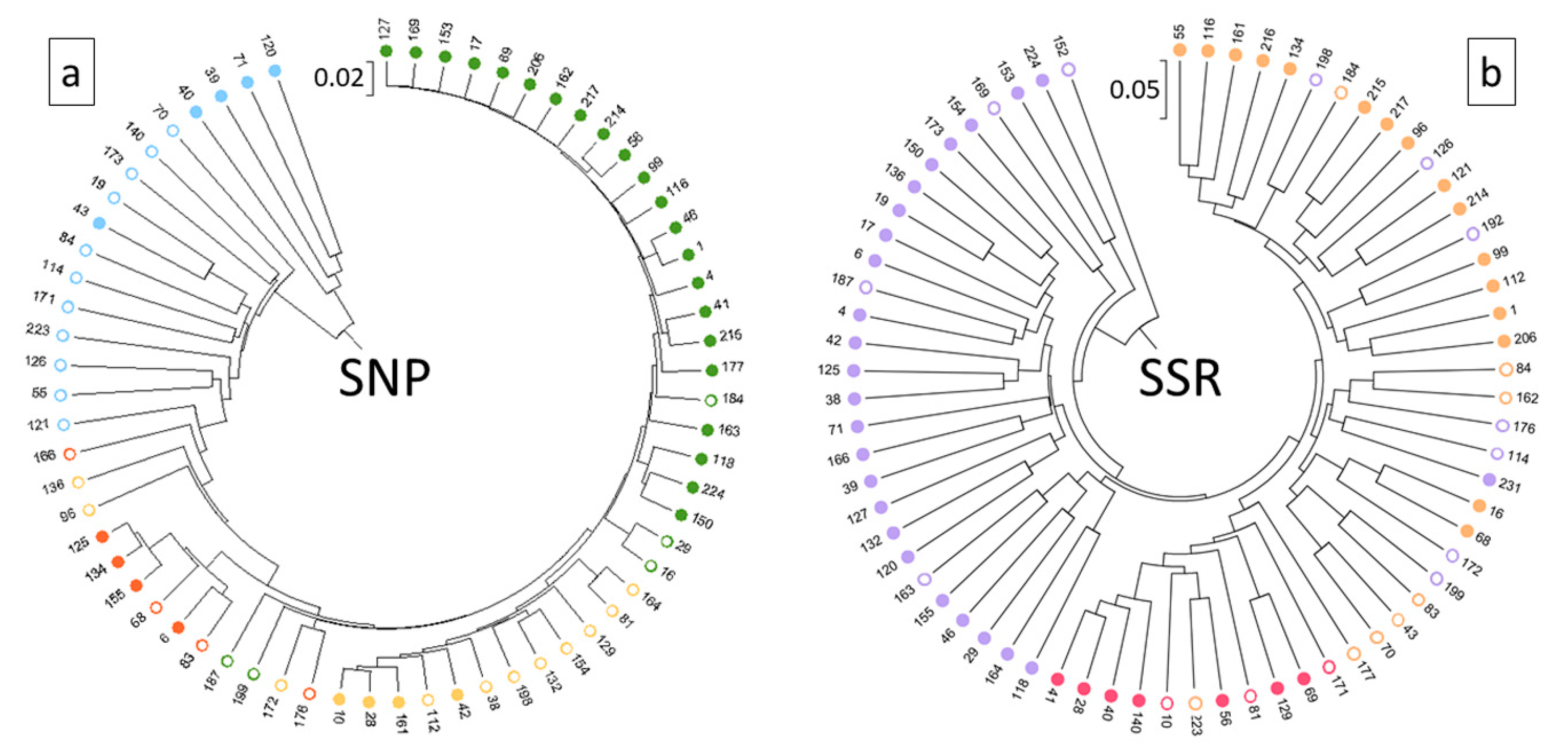
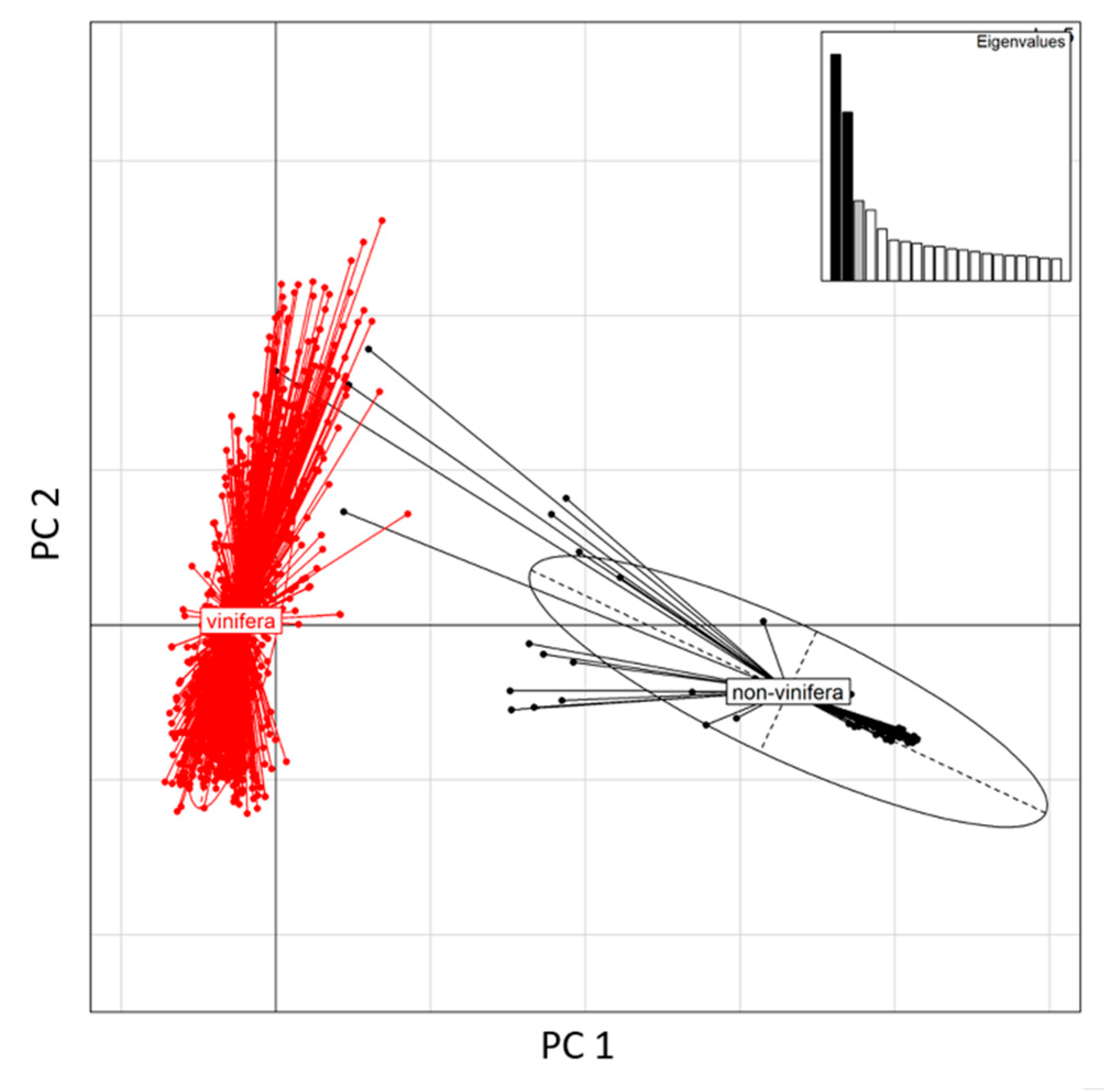
3. Results
4. Discussion
4.1. The 18k SNP Genotyping Array is A Suitable Tool to Characterize Non-vinifera Germplasm
4.2. SNP Profiles Reveal A High Level of Admixture
4.3. SNPs Performed Well in Discriminating Non-vinifera and vinifera Germplasm
4.4. From SSR to SNP Genotyping
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Granett, J.; Walker, M.A.; Kocsis, L.; Omer, A.D. Biology and management of grape phylloxera. Annu. Rev. Entomol. 2001, 46, 387–412. [Google Scholar] [CrossRef]
- Brancadoro, L.; Rabotti, G.; Scienza, A.; Zocchi, G. Mechanisms of Fe-efficiency in roots of Vitis spp. in response to iron deficiency stress. Plant Soil 1995, 171, 229–234. [Google Scholar] [CrossRef]
- Ollat, N.; Peccoux, A.; Papura, D.; Esmenjaud, D.; Marguerit, E.; Tandonnet, J.P.; Bordenave, L.; Cookson, S.J.; Barrieu, F.; Rossdeutsch, L.; et al. Rootstocks as a component of adaptation to environment. In Grapevine in a Changing Environment: A Molecular and Ecophysiological Perspective, 1st ed.; Gerós, H., Chaves, M.A., Gil, H.M., Delrot, S., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2016; pp. 68–108. [Google Scholar]
- Köse, B.; Horuz, A.; Akınoğlu, G. Phenological changes of leaf nutrients in Vitis labrusca L. grape grafted on different rootstocks in heavy clay soil conditions. Erwerbs-Obstbau 2016, 58, 211–217. [Google Scholar]
- Corso, M.; Bonghi, C. Grapevine rootstock effects on abiotic stress tolerance. Plant Sci. 2014, 1, 108–113. [Google Scholar] [CrossRef]
- Keller, M. The Science of Grapevines: Anatomy and Physiology, 2nd ed.; Academic Press: Cambridge, MA, USA, 2015; pp. 1–552. [Google Scholar]
- Riaz, S.; Pap, D.; Uretsky, J.; Laucou, V.; Boursiquot, J.M.; Kocsis, L.; Walker, A. Genetic diversity and parentage analysis of grape rootstocks. Theor. Appl. Genet. 2019, 132, 1847–1860. [Google Scholar] [CrossRef]
- Bacilieri, R.; Lacombe, T.; Le Cunff, L.; Vecchi-staraz, M.; Di Laucou, V.; Genna, B.; Péros, J.; This, P.; Boursiquot, J. Genetic structure in cultivated grapevines is linked to geography and human selection. BMC Plant Biol. 2013, 13, 25. [Google Scholar] [CrossRef] [Green Version]
- Cipriani, G.; Spadotto, A.; Jurman, I.; Di, G.; Crespan, M.; Meneghetti, S.; Frare, E.; Vignani, R.; Cresti, M.; Morgante, M.; et al. The SSR-based molecular profile of 1005 grapevine (Vitis vinifera L.) accessions uncovers new synonymy and parentages, and reveals a large admixture amongst varieties of different geographic origin. Theor. Appl. Genet. 2010, 121, 1569–1585. [Google Scholar] [CrossRef]
- Emanuelli, F.; Lorenzi, S.; Grzeskowiak, L.; Catalano, V.; Stefanini, M.; Troggio, M.; Myles, S.; Martinez-Zapater, J.M.; Zyprian, E.; Moreira, F.M.; et al. Genetic diversity and population structure assessed by SSR and SNP markers in a large germplasm collection of grape. BMC Plant Biol. 2013, 13, 39. [Google Scholar] [CrossRef] [Green Version]
- Laucou, V.; Lacombe, T.; Dechesne, F.; Siret, R.; Bruno, J.P.; Dessup, M.; Dessup, T.; Ortigosa, P.; Parra, P.; Roux, C.; et al. High throughput analysis of grape genetic diversity as a tool for germplasm collection management. Theor. Appl. Genet. 2011, 122, 1233–1245. [Google Scholar] [CrossRef]
- Crespan, M.; Meneghetti, S.; Cancellier, S. Identification and genetic relationship of the principal rootstocks cultivated in Italy. Am. J. Enol. Vitic. 2009, 60, 349–356. [Google Scholar]
- Cseh, A.; Taller, J.; Podmaniczky, P.; Kocsis, L. Comparative analysis of the most widespread grapevine rootstock lines in the world, the teleki lines, with microsatellite markers. Cereal Res. Commun. 2006, 34, 773–776. [Google Scholar] [CrossRef]
- De Andrés, M.T.; Cabezas, J.A.; Cervera, M.T.; Borrego, J.; Martínez-Zapater, J.M.; Jouve, N. Molecular characterization of grapevine rootstocks maintained in germplasm collections. Am. J. Enol. Vitic. 2007, 58, 75–86. [Google Scholar]
- Dzhambazova, T.; Hvarleva, T.; Tsvetkov, I.Y.; Atanassov, A. Characterization of grapevine rootstocks using microsatellite markers. Biotechnol. Biotech. Eq. 2007, 21, 58–62. [Google Scholar] [CrossRef]
- Jahnke, G.; Májer, J.; Szoke, B.; Varga, P.; Kocsisné Molnár, G.; Tarczal, E.; Kocsis, L. Analysis of grape rootstocks by microsatellite markers. Acta Hortic. 2014, 1046, 617–626. [Google Scholar] [CrossRef]
- Sefc, K.M.; Regner, F.; Glössl, J.; Steinkellner, H. Genotyping of grapevine and rootstock cultivars using microsatellite markers. Vitis 1998, 37, 15–20. [Google Scholar]
- Upadhyay, A.; Saboji, M.D.; Reddy, S.; Deokar, K.; Karibasappa, G.S. AFLP and SSR marker analysis of grape rootstocks in Indian grape germplasm. Sci. Hortic. 2007, 112, 176–183. [Google Scholar] [CrossRef]
- Migliaro, D.; De Lorenzis, G.; Di Lorenzo, G.S.; De Nardi, B.; Gardiman, M.; Failla, O.; Brancadoro, L.; Crespan, M. Grapevine non-vinifera genetic diversity assessed by SSR markers as a starting-point for new rootstock breeding programs. Am. J. Enol. Vitic. 2019, 70, 4. [Google Scholar] [CrossRef]
- Lin, H.; Walker, M.A. Identifying grape rootstocks with simple sequence repeat (SSR) DNA markers. Am. J. Enol. Vitic. 1998, 49, 403–407. [Google Scholar]
- Cabezas, J.A.; Ibáñez, J.; Lijavetzky, D.; Vélez, D.; Bravo, G.; Rodríguez, V.; Carreño, I.; Jermakow, A.M.; Carreño, J.; Ruiz-García, L.; et al. A 48 SNP set for grapevine cultivar identification. BMC Plant Biol. 2011, 11, 153. [Google Scholar] [CrossRef] [Green Version]
- Cunha, J.; Ibáñez, J.; Teixeira-Santos, M.; Brazão, J.; Fevereiro, P.; Martínez-Zapater, J.M.; Eiras-Dias, J.E. Characterisation of the Portuguese grapevine germplasm with 48 single-nucleotide polymorphisms. Aust. J. Grape Wine Res. 2016, 22, 504–516. [Google Scholar] [CrossRef]
- De Lorenzis, G.; Mercati, F.; Bergamini, C.; Cardone, M.F.; Lupini, A.; Mauceri, A.; Caputo, A.R.; Abbate, L.; Barbagallo, M.G.; Antonacci, D.; et al. SNP genotyping elucidates the genetic diversity of Magna Graecia grapevine germplasm and its historical origin and dissemination. BMC Plant Biol. 2019, 19, 7. [Google Scholar] [CrossRef]
- Laucou, V.; Launay, A.; Bacilieri, R.; Lacombe, T.; Adam-Blondon, A.F.; Bérard, A.; Chauveau, A.; De Andrés, M.T.; Hausmann, L.; Ibáñez, J.; et al. Extended diversity analysis of cultivated grapevine Vitis vinifera with 10K genome-wide SNPs. PLoS ONE 2018, 13, 1–27. [Google Scholar] [CrossRef]
- Myles, S.; Boyko, A.R.; Owens, C.L.; Brown, P.J.; Grassi, F.; Aradhya, M.K.; Prins, B.; Reynolds, A.; Chia, J.-M.; Ware, D.; et al. Genetic structure and domestication history of the grape. Proc. Natl. Acad. Sci. USA 2011, 108, 3530–3535. [Google Scholar] [CrossRef] [Green Version]
- Ruffa, P.; Raimondi, S.; Boccacci, P.; Abbà, S.; Schneider, A. The key role of “Moscato bianco” and “Malvasia aromatica di Parma” in the parentage of traditional aromatic grape varieties. Tree Genet. Genomes. 2016, 12, 50. [Google Scholar] [CrossRef]
- Myles, S.; Chia, J.; Hurwitz, B.; Simon, C.; Zhong, G.Y.; Ware, D. Rapid genomic characterization of the genus Vitis. PLoS ONE 2010, 5, e8219. [Google Scholar] [CrossRef] [Green Version]
- Marrano, A.; Birolo, G.; Prazzoli, M.L.; Lorenzi, S.; Valle, G.; Grando, M.S. SNP-Discovery by RAD-sequencing in a germplasm collection of wild and cultivated grapevines (V. vinifera L.). PLoS ONE 2017, 12, e0170655. [Google Scholar] [CrossRef]
- Guo, D.-L.; Zhao, H.-L.; Li, Q.; Zhang, G.-H.; Jiang, J.-F.; Liu, C.-H.; Yu, Y.-H. Genome-wide association study of berry-related traits in grape Vitis vinifera L. based on genotyping-by-sequencing markers. Hortic. Res. 2019, 6, 11. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Fan, X.; Zhang, Y.; Jiang, J.; Liu, C. Identification of favorable SNP alleles and candidate genes for seedlessness in Vitis vinifera L. using genome-wide association mapping. Euphytica 2017, 213, 136. [Google Scholar] [CrossRef]
- De Lorenzis, G.; Squadrito, M.; Rossoni, M.; di Lorenzo, G.S.; Brancadoro, L.; Scienza, A. Study of intra-varietal diversity in biotypes of Aglianico and Muscat of Alexandria (Vitis vinifera L.) cultivars. Aust. J. Grape Wine Res. 2017, 23, 132–142. [Google Scholar] [CrossRef]
- Degu, A.; Morcia, C.; Tumino, G.; Hochberg, U.; Toubiana, D.; Mattivi, F.; Schneider, A.; Bosca, P.; Cattivelli, L.; Terzi, V.; et al. Metabolite profiling elucidates communalities and differences in the polyphenol biosynthetic pathways of red and white Muscat genotypes. Plant Physiol. Biochem. 2015, 86, 24–33. [Google Scholar] [CrossRef]
- Mercati, F.; De Lorenzis, G.; Brancadoro, L.; Lupini, A.; Abenavoli, M.R.; Barbagallo, M.G.; Di Lorenzo, R.; Scienza, A.; Sunseri, F. High-throughput 18K SNP array to assess genetic variability of the main grapevine cultivars from Sicily. Tree Genet. Genomes. 2016, 12, 59. [Google Scholar] [CrossRef] [Green Version]
- Sunseri, F.; Lupini, A.; Mauceri, A.; De Lorenzis, G.; Araniti, F.; Brancadoro, L.; Dattola, A.; Gullo, G.; Zappia, R.; Mercati, F. Single nucleotide polymorphism profiles reveal an admixture genetic structure of grapevine germplasm from Calabria, Italy, uncovering its key role for the diversification of cultivars in the Mediterranean Basin. Aust. J. Grape Wine Res. 2018, 24, 345–359. [Google Scholar] [CrossRef]
- De Lorenzis, G.; Chipashvili, R.; Failla, O.; Maghradze, D. Study of genetic variability in Vitis vinifera L. germplasm by high-throughput Vitis18kSNP array: The case of Georgian genetic resources. BMC Plant Biol. 2015, 15, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; Gupta, S.; Jin, L. PEAS V1.0: A package for elementary analysis of SNP data. Mol. Ecol. Resour. 2010, 10, 1085–1088. [Google Scholar] [CrossRef]
- Kamvar, Z.N.; Tabima, J.F.; Grünwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/ or sexual reproduction. PeerJ 2014, 2, e281. [Google Scholar] [CrossRef] [Green Version]
- Caroli, S.; Santoni, S.; Ronfort, J. AMaCAID: A useful tool for accurate marker choice for accession identification and discrimination. Mol. Ecol. Resour. 2011, 11, 733–738. [Google Scholar] [CrossRef]
- R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.eea.europa.eu/data-and-maps/indicators/oxygen-consuming-substances-in-rivers/r-development-core-team-2006 (accessed on 15 March 2006).
- Frichot, E.; Franc, O. LEA: An R package for landscape and ecological association studies. Methods Ecol. Evol. 2015, 6, 925–929. [Google Scholar] [CrossRef]
- Jombart, T. adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [Green Version]
- Nei, M. Genetic distance between populations. Am. Nat. 1972, 106, 283–292. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Sefc, K.; Regner, F.; Turetschek, E.; Glössl, J.; Steinkeller, H. Identification of microsatellite sequences in Vitis riparia and their applicability for genotyping of different Vitis species. Genome 1999, 42, 367–373. [Google Scholar] [CrossRef]
- D’Onofrio, C.; De Lorenzis, G.; Giordani, T.; Natali, L.; Cavallini, A. Retrotransposon-based molecular markers for grapevine species and cultivars identification. Tree Genet. Genomes 2010, 6, 451–466. [Google Scholar] [CrossRef]
- Guo, D.; Guo, M.; Hou, X.; Zhang, G. Molecular diversity analysis of grape varieties based on iPBS markers. Biochem. Syst. Ecol. 2014, 52, 27–32. [Google Scholar] [CrossRef]
- Mullins, M.G.; Bouquet, A.; Williams, L.E. Biology of the Grapevine, 1st ed.; Cambridge University Press: Cambridge, UK, 1992; pp. 80–146. [Google Scholar]
- Aradhya, M.; Wang, Y.; Walker, M.A.; Prins, B.H.; Koehmstedt, A.M.; Velasco, D.; Gerrath, J.M.; Dangle, G.S.; Preece, J.E. Genetic diversity, structure, and patterns of differentiation in the genus Vitis. Plant Syst. Evol. 2013, 299, 317–330. [Google Scholar] [CrossRef]
- Wan, Y.; Schwaninger, H.R.; Baldo, A.M.; Labate, J.A.; Zhong, G.Y.; Simon, C.J. A phylogenetic analysis of the grape genus (Vitis L.) reveals broad reticulation and concurrent diversification during neogene and quaternary climate change. BMC Evol. Biol. 2013, 13, 141. [Google Scholar] [CrossRef] [Green Version]
- Maul, E.; Sudharma, K.N.; Kecke, S.; Marx, G.; Müller, C.; Audeguin, L.; Boselli, M.; Boursiquot, J.M.; Bucchetti, B.; Cabello, R.; et al. The European Vitis Database (www.eu-vitis.de): A technical innovation through an online uploading and interactive modification system. VITIS 2012, 51, 79–85. [Google Scholar]
- Wahlund, S. Zusammensetzung von populationen und korrelationserscheinungen von standpunkt der vererbungslehre aus betrachtet. Hereditas 1928, 11, 65–106. [Google Scholar] [CrossRef]
- Klein, L.L.; Miller, A.J.; Ciotir, C.; Hyma, K.; Uribe-Convers, S.; Londo, J. High-throughput sequencing data clarify evolutionary relationships among North American Vitis species and improve identification in USDA Vitis germplasm collections. Am. J. Bot. 2018, 105, 215–226. [Google Scholar] [CrossRef]
- Chagné, D.; Crowhurst, R.N.; Troggio, M.; Davey, M.W.; Gilmore, B.; Lawley, C.; Vanderzande, S.; Hellens, R.P.; Kumar, S.; Cestaro, A.; et al. Genome-wide SNP detection, validation, and development of an 8K SNP array for apple. PLoS ONE 2012, 7, e31745. [Google Scholar] [CrossRef] [Green Version]
- Felcher, K.J.; Coombs, J.J.; Massa, A.N.; Hansey, C.N.; Hamilton, J.P.; Veilleux, R.E.; Buell, C.R.; Douches, D.S. Integration of two diploid potato linkage maps with the potato genome sequence. PLoS ONE 2012, 7, e36347. [Google Scholar] [CrossRef] [Green Version]
- Ganal, M.W.; Durstewitz, G.; Polley, A.; Berard, A.; Buckler, E.S.; Charcosset, A.; Clarke, J.D.; Graner, E.M.; Hansen, M.; Joets, J.; et al. A Large Maize (Zea mays L.) SNP genotyping array: Development and germplasm genotyping, and genetic mapping to compare with the B73 reference genome. PLoS ONE 2011, 6, e28334. [Google Scholar] [CrossRef] [Green Version]
- Sim, S.-C.; Durstewitz, G.; Plieske, J.; Wieseke, R.; Ganal, M.W.; Van Deynze, A.; Hamilton, J.P.; Buell, C.R.; Causse, M.; Wijeratne, S.; et al. Development of a large SNP genotyping array and generation of high density genetic maps in tomato. PLoS ONE 2012, 7, e40563. [Google Scholar] [CrossRef]
- Verde, I.; Bassil, N.; Scalabrin, S.; Gilmore, B.; Lawley, C.T.; Gasic, K.; Micheletti, D.; Rosyara, U.R.; Cattonaro, F.; Vendramin, E.; et al. Development and evaluation of a 9K SNP Array for peach by internationally coordinated SNP detection and validation in breeding germplasm. PLoS ONE 2012, 7, e35668. [Google Scholar] [CrossRef]
- Bérard, A.; Le Paslier, M.C.; Dardevet, M.; Exbrayat-Vinson, F.; Bonnin, I.; Cenci, A.; Haudry, A.; Brunel, D.; Ravel, C. High-throughput single nucleotide polymorphism genotyping in wheat (Triticum spp.). Plant Biotechnol. J. 2009, 7, 364–374. [Google Scholar]
- Ha, B.K.; Hussey, R.S.; Boerma, H.R. Development of SNP assays for marker-assisted selection of two southern root-knot nematode resistance QTL in soybean. Crop Sci. 2007, 47, 73–82. [Google Scholar] [CrossRef]
- Tian, F.; Bradbury, P.J.; Brown, P.J.; Hung, H.; Sun, Q.; Flint-Garcia, S.; Rocheford, T.R.; McMullen, M.D.; Holland, J.B.; Buckler, E.S. Genome-wide association study of leaf architecture in the maize nested association mapping population. Nat. Genet. 2011, 43, 159–162. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession ID | Genotype | Pedigree |
|---|---|---|
| 1 | 101.14 Millardet et de Grasset | V. riparia × V. rupestris |
| 4 | 110 Richter | V. rupestris × V. berlandieri |
| 6 | 1202 C | V. vinifera × V. rupestris |
| 10 | 161.49 Couderc | V. berlandieri × V. riparia |
| 16 | 3309 Couderc | V. riparia × V. rupestris |
| 17 | 333 Ecole de Montpellier or Tisserand | V. vinifera × V. berlandieri |
| 19 | 41 B Millardet et de Grasset | V. vinifera × V. berlandieri |
| 28 | Cosmo 10 | V. berlandieri × V. riparia |
| 29 | Dog Ridge | V. rupestris × V. candicans |
| 38 | Isabella | V. labrusca × V. vinifera |
| 39 | Jacquez | V. aestivalis × V. vinifera |
| 40 | Geilweilerhof V.348 | V. vinifera |
| 41 | Kober 5BB | V. berlandieri × V. riparia |
| 42 | LN 33 or LLYOD’S NUMBER 33 | V. riparia × V. longii × V. vinifera |
| 43 | Malegue 44.53 | V. riparia × V. cordifolia × V. rupestris |
| 46 | Salt Creek | Unknown |
| 55 | Vitis riparia Fabre | V. riparia |
| 56 | Vitis riparia Gloire de Montpellier | V. riparia |
| 68 | Genotype 01 | Unknown |
| 69 | Genotype 02 | V. berlandieri × V. riparia × V. cinerea |
| 70 | Genotype 03 | V. berlandieri × V. riparia × V. cordifolia × V. rupestris |
| 71 | Genotype 04 | Unknown |
| 81 | Genotype 15 | V. berlandieri × V. riparia × V. rupestris |
| 83 | Genotype 17 | V. berlandieri × V. riparia |
| 84 | Genotype 18 | Unknown |
| 96 | Genotype 29 | V. riparia × V. vinifera × ? |
| 99 | Genotype 33 | V. riparia × V. longii × ? |
| 112 | Genotype 46 | Unknown |
| 114 | Genotype 48 | Unknown |
| 116 | Vitis riparia Lombard | V. riparia |
| 118 | Genotype 52 | Unknown |
| 120 | Genotype 54 | Unknown |
| 121 | Genotype 55 | Unknown |
| 125 | Vitis labrusca Muncy | V. labrusca |
| 126 | Genotype 60 | Unknown |
| 127 | Genotype 61 | Unknown |
| 129 | Genotype 63 | V. berlandieri × V. riparia × ? |
| 132 | Genotype 66 | Unknown |
| 134 | Genotype 68 | Unknown |
| 136 | Genotype 70 | V. vinifera × V. berlandieri × V. riparia × V. candicans |
| 140 | Genotype 74 | V. berlandieri × V. riparia |
| 150 | Kober 125 AA | V. berlandieri × V. riparia |
| 152 | Genotype 86 | Unknown |
| 153 | Genotype 87 | Unknown |
| 154 | Genotype 88 | Unknown |
| 155 | Genotype 89 | Unknown |
| 161 | Genotype 95 | Unknown |
| 162 | Genotype 96 | Unknown |
| 163 | Genotype 97 | Unknown |
| 164 | Genotype 98 | Unknown |
| 166 | Genotype 100 | Unknown |
| 169 | Genotype 103 | Unknown |
| 171 | Genotype 105 | Unknown |
| 172 | Genotype 106 | V. riparia × V. rupestris × ? |
| 173 | Genotype 107 | Unknown |
| 176 | Genotype 110 | Unknown |
| 177 | Genotype 111 | Unknown |
| 184 | Genotype 118 | Unknown |
| 187 | 143 B Millardet et De Grasset | V. vinifera × ? |
| 192 | 202-4 Millardet et De Grasset | V. riparia × V. longii × ? |
| 198 | 33 Ecole de Montpellier | V. berlandieri × V. riparia |
| 199 | 420 B Millardet et De Grasset | V. berlandieri × V. riparia |
| 206 | Dufour 11 F | V. riparia × V. rupestris |
| 214 | Genotype 148 | Unknown |
| 215 | Vitis riparia Sericea | V. riparia |
| 216 | Vitis riparia Sombre | V. riparia |
| 217 | Vitis riparia Tomenteux | V. riparia |
| 223 | Genotype 157 | V. labrusca × V. riparia × V. rupestris |
| 224 | Genotype 158 | Unknown |
| 231 | Genotype 165 | Unknown |
| Marker | Plant Material | N | Ho | He |
|---|---|---|---|---|
| SNP | Core collection | 66 | 0.143 | 0.157 |
| Group 1 | 8 | 0.100 | 0.095 | |
| Group 2 | 16 | 0.303 | 0.280 | |
| Group 3 | 15 | 0.125 | 0.099 | |
| Group 4 | 27 | 0.071 | 0.067 | |
| SSR | Core collection | 70 | 0.823 | 0.879 |
| Group 1 | 10 | 0.882 | 0.730 | |
| Group 2 | 24 | 0.779 | 0.771 | |
| Group 3 | 36 | 0.836 | 0.891 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bianchi, D.; Brancadoro, L.; De Lorenzis, G. Genetic Diversity and Population Structure in a Vitis spp. Core Collection Investigated by SNP Markers. Diversity 2020, 12, 103. https://doi.org/10.3390/d12030103
Bianchi D, Brancadoro L, De Lorenzis G. Genetic Diversity and Population Structure in a Vitis spp. Core Collection Investigated by SNP Markers. Diversity. 2020; 12(3):103. https://doi.org/10.3390/d12030103
Chicago/Turabian StyleBianchi, Davide, Lucio Brancadoro, and Gabriella De Lorenzis. 2020. "Genetic Diversity and Population Structure in a Vitis spp. Core Collection Investigated by SNP Markers" Diversity 12, no. 3: 103. https://doi.org/10.3390/d12030103
APA StyleBianchi, D., Brancadoro, L., & De Lorenzis, G. (2020). Genetic Diversity and Population Structure in a Vitis spp. Core Collection Investigated by SNP Markers. Diversity, 12(3), 103. https://doi.org/10.3390/d12030103