Citizen Science Confirms the Rarity of Fruit Bat Pollination of Baobab (Adansonia digitata) Flowers in Southern Africa

 ,
,
Abstract
:1. Introduction
2. Methods
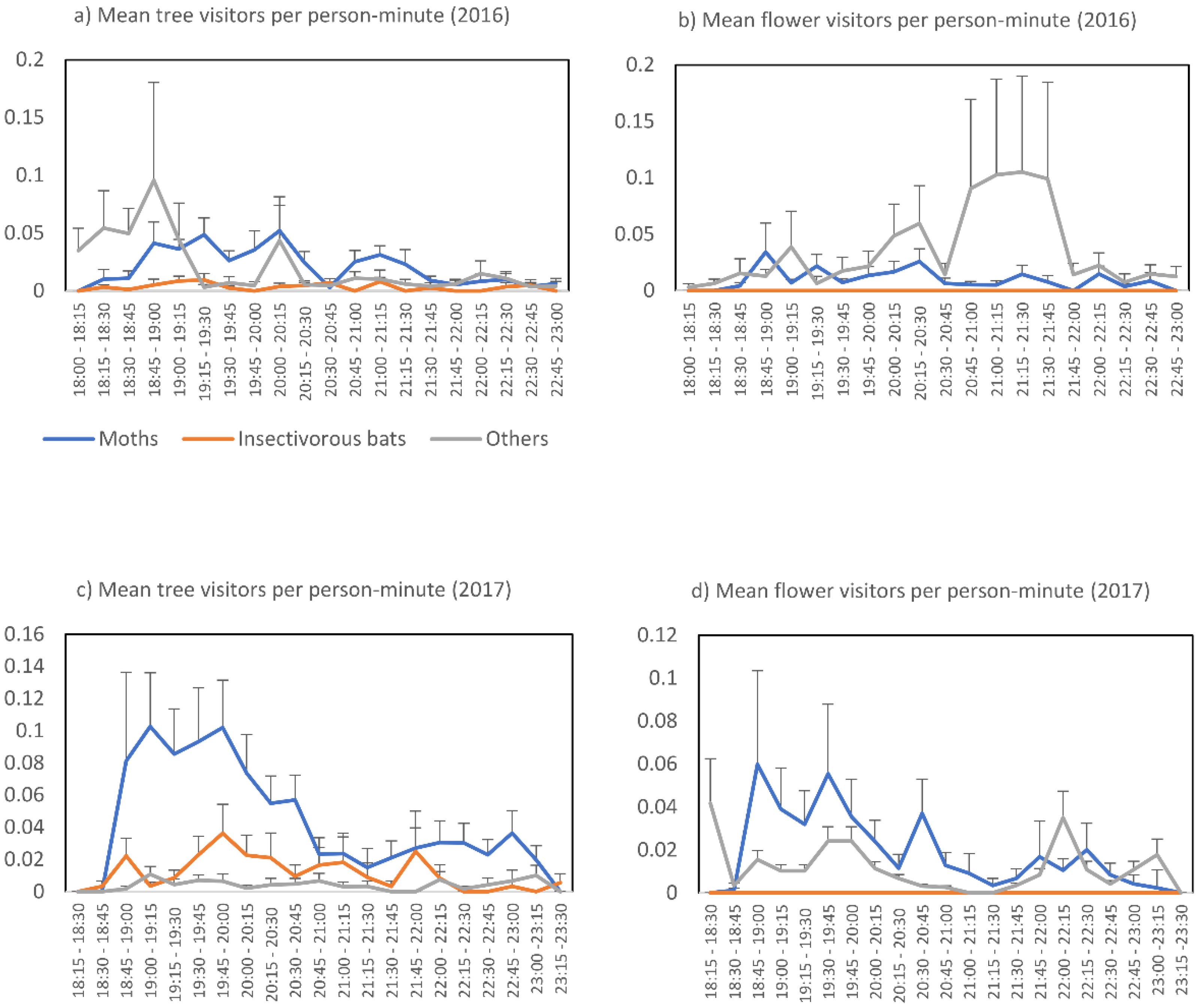
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A. Poster Advertising 2017 Baobab Blitz

Appendix B. Copy of Blank Form Used for Baobab Blitz (2017)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Interval | Moth Visitors to Tree | Moth Visitors to Flower | Bat Visitors to Tree | Bat Visitors to Flower | Other* Visitors to Tree | Other* Visitors to Flower | Comments |
| 18:00–18:15 | |||||||
| 18:15–18:30 | |||||||
| 18:30–18:45 | |||||||
| 18:45–19:00 | |||||||
| 19:00–19:15 | |||||||
| 19:15–19:30 | |||||||
| 19:30–19:45 | |||||||
| 19:45–20:00 | |||||||
| 20:00–20:15 | |||||||
| 20:15–20:30 | |||||||
| Interval | Moth visitors to tree | Moth visitors to flower | Bat visitors to tree | Bat visitors to flower | Other* visitors to tree | Other* visitors to flower | Comments |
| 20:30–20:45 | |||||||
| 20:45–21:00 | |||||||
| 21:00–21:15 | |||||||
| 21:15–21:30 | |||||||
| 21:30–21:45 | |||||||
| 21:45–22:00 | |||||||
| 22:00–22:15 | |||||||
| 22:15–22:30 | |||||||
| 22:30–22:45 | |||||||
| 22:45–23:00 | |||||||
| Interval | Moth visitors to tree | Moth visitors to flower | Bat visitors to tree | Bat visitors to flower | Other* visitors to tree | Other* visitors to flower | Comments |
| 23:00–23:15 (optional) | |||||||
| 23:15–23:30 (optional) | |||||||
| 23:30–23:45 (optional) | |||||||
| 23:45–24:00 (optional) |
Appendix C. Baobab Blitz Information Packet
- Torch (with extra batteries)
- ○
- Red torch, if possible
- Chairs
- Cold drinks
- Cell phone camera (to take a photo of your tree and flower on that tree) or a normal camera that you can upload the photo.
- GPS or Smart phone to record your location (lat and long) or the farm name and where the tree is located.
- Clipboard with pen (extra one just in case)
- Datasheets, see attached via email (see BB Datasheet)
- Watch or phone (to record time)
- Other recommendations:
- ○
- Table
- ○
- Insect repellent


- Eat fruit and nectar
- Larger that insectivorous bats (usually)
- Do not echolocate
- Our ears can detect their metallic “ping ping” calls.
- Forage for many hours throughout the night.
- Should land on the flower to drink the nectar from behind the petals.
- Eat insects
- Use echolocation
- Call is above most people’s hearing
- Usually smaller and faster flying than fruit bats.
- Tend to forage most just after sunset and again before sunrise
- Will not land on the flower but might fly fast through the tree chasing an insect.
Appendix D
| (a) Moth Visitors to Tree | Estimate | Std. Error | z Value | Pr (>|z|) |
|---|---|---|---|---|
| (Intercept) = 0 | −0.3373 | 0.5174 | −0.652 | 0.514 |
| as.factor (Year) 2017 = 0 | 0.4832 | 0.3218 | 1.502 | 0.133 |
| as.factor (Expert) Yes = 0 | 0.5736 | 0.4097 | 1.400 | 0.161 |
| as.factor (Weather) Fine = 0 | 0.6001 | 0.3986 | 1.505 | 0.132 |
| (b) Moth visitors to flower | ||||
| (Intercept) = 0 | −1.9575 | 0.9562 | −2.047 | 0.0406 * |
| as.factor (Year) 2017 = 0 | 0.3737 | 0.4953 | 0.755 | 0.4505 |
| as.factor (Expert) Yes = 0 | 0.9865 | 0.7280 | 1.355 | 0.1754 |
| as.factor (Weather) Fine = 0 | 1.1070 | 0.7165 | 1.545 | 0.1223 |
| (c) Bat visitors to tree | ||||
| (Intercept) = 0 | −2.6659 | 0.9627 | −2.769 | 0.00562 ** |
| as.factor (Year) 2017 = 0 | 0.9878 | 0.5339 | 1.850 | 0.06429 |
| as.factor (Expert) Yes = 0 | 1.1892 | 0.7956 | 1.495 | 0.13501 |
| as.factor (Weather) Fine = 0 | 0.5044 | 0.6074 | 0.831 | 0.40625 |
| (d) Other visitors to tree | ||||
| (Intercept) = 0 | −1.5791 | 1.6047 | −0.984 | 0.325 |
| as.factor (Year) 2017 = 0 | −1.4977 | 1.0930 | −1.370 | 0.171 |
| as.factor (Expert) Yes = 0 | 1.2967 | 1.4412 | 0.900 | 0.368 |
| as.factor (Weather) Fine = 0 | 0.9304 | 1.0392 | 0.895 | 0.371 |
| (e) Other visitors to flower | ||||
| (Intercept) = 0 | −0.8228 | 2.0139 | −0.409 | 0.683 |
| as.factor (Year) 2017 = 0 | −1.1185 | 1.1535 | −0.970 | 0.332 |
| as.factor (Expert) Yes = 0 | 1.7130 | 1.9606 | 0.874 | 0.382 |
| as.factor (Weather) Fine = 0 | 0.1434 | 1.0045 | 0.143 | 0.886 |
References
- Harris, B.J.; Baker, H.G. Pollination of flowers by bats in Ghana. Niger. Field 1959, 24, 154–159. [Google Scholar]
- Ayensu, E.S. Plant & bat interactions in West Africa. Ann. Mo. Bot. Gard. 1974, 61, 702–727. [Google Scholar]
- Fujita, M.S. Flying fox (Chiroptera: Pteropodidae) pollination, seed dispersal, and economic importance. In A Tabular Summary; Bat Conservation International (Resource Publication No.2): Austin, TX, USA, 1991. [Google Scholar]
- Barclay, R.M.R. Do plants pollinated by flying fox bats (Megachiroptera) provide an extra calcium reward in their nectar? Biotropica 2002, 34. [Google Scholar] [CrossRef]
- Hodgkinson, R.; Balding, S.T.; Zubaid, A.; Kunz, T.H. Fruit Bats (Chiroptera: Pteropodidae) as seed dispersers and pollinators in a lowland Malaysian rain forest. Biotropica 2003, 35, 491–502. [Google Scholar] [CrossRef]
- Andriafidison, D.; Andrianaivoarivelo, R.A.; Ramilijaona, O.R.; Andrianaivoarivelo, R.A.; Racey, P.A.; Razafindrakoto, N.; Jenkins, R.K.B. Nectarivory by endemic Malagasy fruit bats during the dry season. Biotropica 2006, 38, 85–90. [Google Scholar]
- Bumrungsri, S.; Harbit, A.; Benzie, C.; Carmouche, K.; Sridith, K.; Racey, P. The pollination ecology of two species of Parkia (Mimosaceae) in southern Thailand. J. Trop. Ecol. 2008, 24, 467–475. [Google Scholar] [CrossRef]
- Bumrungsri, S.; Lang, D.; Harrower, C.; Sripaoraya, E.; Kitpipit, K.; Racey, P.A. The dawn bat, Eonycteris spelaea Dobson (Chiroptera: Pteropodidae) feeds mainly on pollen of economically important food plants in Thailand. Acta Chiropterol. 2013, 15, 95–104. [Google Scholar] [CrossRef]
- Fleming, T.H.; Geiselman, C.; Kress, W.J. The evolution of bat pollination: A phylogenetic perspective. Ann. Bot. 2009, 104, 1017–1043. [Google Scholar] [CrossRef] [PubMed]
- Faigri, K.; Van der Pijl, L. The Principles of Pollination Ecology; Pergamon Press: Oxford, UK, 1979. [Google Scholar]
- Fenster, C.B.; Armbruster, W.S.; Wilson, P.; Dudash, M.R.; Thomson, J.D. Pollination syndromes and floral specialization. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 375–403. [Google Scholar] [CrossRef]
- Waser, N.M. Specialization and generalization in plant-pollinator interactions: An historical perspective. In Plant-Pollinator Interactions: From Specialization to Generalization; Waser, N.M., Ollerton, J., Eds.; University of Chicago: Chicago, IL, USA, 2006; pp. 3–17. [Google Scholar]
- Nathan, P.T.; Raghuram, H.; Elangovan, V.; Karuppudurai, T.; Marimuthu, G. Bat pollination of kapok tree Ceiba pentandra. Curr. Sci. 2005, 88, 1679–1681. [Google Scholar]
- Srithongchuay, T.; Bumrungsri, S.; Sripao-raya, E. The pollination ecology of the late-successional tree, Oroxylum indicum (Bignoniaceae) in Thailand. J. Trop. Ecol. 2008, 24, 477–484. [Google Scholar] [CrossRef]
- Bumrungsri, S.; Sripaoraya, E.; Chongsiri, T.; Sridith, K.; Racey, P.A. The pollination ecology of durian (Durio zibethinus, Bombacaceae) in southern Thailand. J. Trop. Ecol. 2009, 25, 85–92. [Google Scholar] [CrossRef]
- Sidibe, M.; Williams, J.T. Baobab; Adansonia Digitata International Centre for Underutilised Crops: Southampton, UK, 2002. [Google Scholar]
- Wickens, G.E.; Lowe, P. The Baobabs: Pachycauls of Africa, Madagascar and Australia; Springer: Berlin, Germany, 2008. [Google Scholar]
- Venter, S.M.; Witkowski, E.T.F. Baobab (Adansonia digitata L.) density, size-class distribution and population trends between four land-use types in northern Venda, South Africa. For. Ecol. Manag. 2010, 259, 294–300. [Google Scholar] [CrossRef]
- Leong Pock Tsy, J.M.; Lumaret, R.; Mayne, D.; Vall, A.O.; Abutaba, Y.I.; Sagna, M.; Rakotondralambo Raoseta, S.O.; Danthu, P. Chloroplast DNA phylogeography suggests a West African centre of origin for the baobab, Adansonia digitata L. (Bombacoideae, Malvaceae). Mol. Ecol. 2009, 18, 1707–1715. [Google Scholar] [CrossRef]
- Baum, D.A. The comparative pollination and floral biology of baobabs (Adansonia, Bombacaceae). Ann. Mo. Bot. Gard. 1995, 82, 322–348. [Google Scholar] [CrossRef]
- Start, T. Pollination of the baobab (Adansonia digitata L.) by the fruit bat Rousettus. East Afr. Wildl. J. 1972, 10, 71–72. [Google Scholar] [CrossRef]
- Venter, S.M.; Glennon, K.L.; Witkowski, E.T.F.; Baum, D.; Cron, G.V.; Tivakudze, R.; Karimi, N. Baobabs (Adansonia digitata L.) are self-incompatible and ‘male’ trees can produce fruit if hand-pollinated. South Afr. J. Bot. 2017, 109, 263–268. [Google Scholar] [CrossRef]
- Wilson, D.E.; Mittermeier, R.A. Handbook of the Mammals of the World, Volume 9: Bats; Lynx Edicions: Barcelona, Spain, 2019. [Google Scholar]
- Monadjem, A.; Taylor, P.J.; Cotterill, F.P.D.; Schoeman, M.C. Bats of Southern and South-Central Africa: A Biogeographic and Taxonomic Synthesis; Wits University Press: Johannesburg, South Africa, 2010; p. 596. [Google Scholar]
- Welford, W.; Venter, S.; Dohse, C. Harvesting from the tree of life–responsible commercialization of baobab in South Africa and Malawi. In Ecological Sustainability for Non-Timber Forest Products; Shackleton, C.M., Pandey, A.K., Ticktin, T., Eds.; Oxon: Routledge, UK, 2015. [Google Scholar]
- Dickinson, J.L.; Shirk, J.; Bonter, D.; Bonney, R.; Crain, R.L.; Martin, J.; Phillips, T.; Purcell, K. The current state of citizen science as a tool for ecological research and public engagement. Front. Ecol. Environ. 2012, 10, 291–297. [Google Scholar] [CrossRef] [Green Version]
- Mucina, L.; Rutherford, M.C. The Vegetation of South Africa, Lesotho and Swaziland; South African Biodiversity Instutute: Pretoria, South Africa, 2006. [Google Scholar]
- Robinson, G.A. Limpopo Valley National Park: Towards Trans-Frontier Conservation in South Africa; South African National Parks: Pretoria, South Africa, 1996. [Google Scholar]
- Van Wilgen, N.J.; Goodall, V.; Holness, S.; Chown, S.L.; McGeoch, M.A. Rising temperatures and changing rainfall patterns in South Africa’s national parks. Int. J. Climatol. 2016, 36, 709–721. [Google Scholar] [CrossRef]
- Van Wilgen, N.J.; Herbst, M. Taking Stock of Parks in a Changing World: The SANParks Global Environmental Change Assessment; SANParks: Cape Town, South Africa, 2017. [Google Scholar]
- Venter, S.M.; Witkowski, E.T.F. Phenology, flowering and fruit-Set patterns of baobabs, Adansonia digitata, in southern Africa. For. Ecol. Manag. 2019, 453, 117593. [Google Scholar] [CrossRef]
- Everyone Can Become A Baobab Scientist. Available online: https://zoutpansberger.co.za/articles/news/39227/2016-10-15/everyone-can-become-a-baobab-blitz-scientist (accessed on 13 March 2020).
- More Research on Baobab Pollinators. Available online: https://zoutpansberger.co.za/articles/news/41377/2017-03-23/more-research-needed-on-baobab-pollinators (accessed on 13 March 2020).
- Help Solve Baobab Flower Mystery. Available online: https://www.zoutpansberger.co.za/articles/news/45072/2017-11-25/help-solve-baobab-flower-pollination-mystery (accessed on 13 March 2020).
- Djossa, B.A.; Toni, H.C.; Adekanmbi, I.D.; Tognon, F.K.; Sinsin, B.A. Do flying foxes limit flower abortion in African baobab (Adansonia digitata)? Case study in Benin, West Africa. Fruits 2015, 70, 281–287. [Google Scholar] [CrossRef] [Green Version]
- Pettersson, S.; Ervik, E.; Knudsen, J.T. Floral scent of bat-pollinated species: West Africa vs. the New World. Biol. J. Linn. Soc. 2004, 82, 161–168. [Google Scholar] [CrossRef]
- Venter, S.M.; Witkowski, E.T.F. Fruits of our labour: Contribution of commercial baobab (Adansonia digitata L.) fruit harvesting to the livelihoods of marginalized people in northern Venda, South Africa. Agrofor. Syst. 2012, 87, 159–172. [Google Scholar] [CrossRef]
- Omondi, M.; Rimberia, F.K.; Wainaina, C.M.; Mukundi, J.B.N.; Orina, J.; Gebauer, J.; Kehlenbeck, K. Fruit morphological diversity and productivity of baobab (Adansonia digitata L.) in coastal and lower eastern Kenya. For. Trees Livelihoods 2019, 28, 266–280. [Google Scholar] [CrossRef]




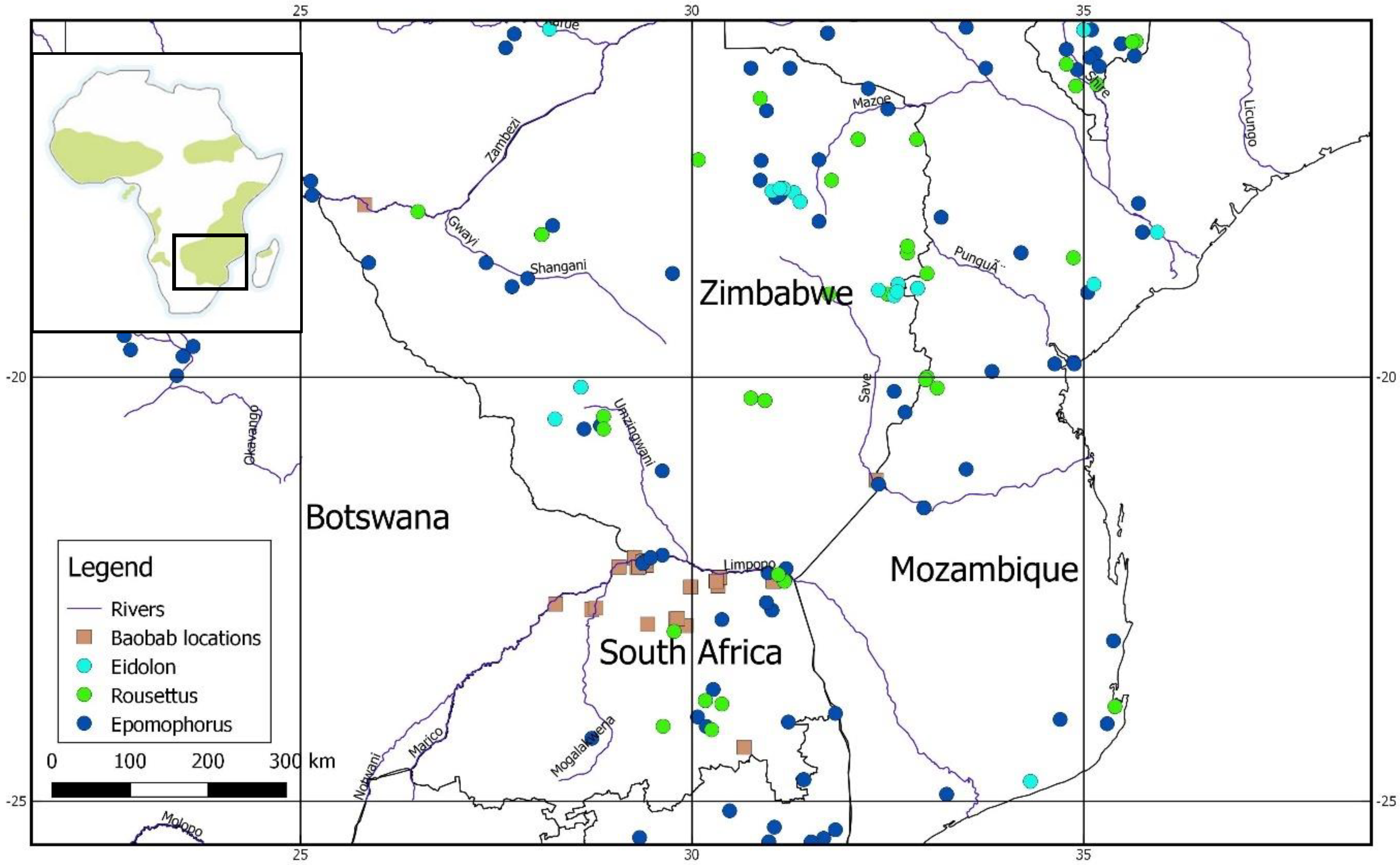
| Locality Name | Lat. | Long. | Date | N | Age Range | Weather | Wind | Experience Level (Researchers/Students Present or Absent) | Time Start | Time End | No. Flowers Open | Land Use |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hector and Hazel’s Farm | −22.87 | 29.79 | 6-Nov-16 | 6 | 24 | clear | slight breeze | Present | 18:30 | 23:00 | 27 | cattle and game farm |
| Goro | −22.94 | 29.43 | 19-Nov-16 | 3 | 24 | partly cloudy | slight breeze | Present | 18:00 | 21:45 | 55 | game farm |
| Mapesu Private Game Reserve | −22.15 | 29.26 | 18-Nov-16 | 4 | 17–38 | rain | strong wind | Absent | 18:00 | 23:00 | 39 | game farm |
| The Oaks Village | −24.37 | 30.66 | 18-Nov-16 | 2 | 22–32 | overcast | slight breeze | Absent | 17:30 | 22:30 | 8 | cattle farm |
| Skirbeek 73MT | −22.48 | 30.33 | 18-Nov-16 | 1 | 57 | overcast | strong wind | Absent | 18:00 | 22:00 | 4 | game farm |
| Mogalakwena Game Reserve | −22.76 | 28.72 | 18-Nov-16 | 7 | 20–42 | overcast | no wind | Present | 18:00 | 21:00 | - | game farm |
| Mogalakwena Game Reserve | −22.74 | 28.77 | 19-Nov-16 | 6 | 20–48 | rain | slight breeze | Present | 18:00 | 23:00 | 8 | game farm |
| Mmabolela Estates, Farm Tuli, district Maasstroom | −22.70 | 28.26 | 18-Nov-16 | 2 | 68–69 | partly cloudy | strong wind | Present | 19:00 | 22:00 | 21 | game farm |
| Chilo Gorge Lodge, Zimbabwe | −21.23 | 32.35 | 25-Nov-16 | 3 | 45–55 | overcast | strong wind | Present | 18:00 | 23:00 | 130 | game farm |
| Elgen (Skelmwater Research Plot) | −22.50 | 29.98 | 19-Nov-16 | 3 | 45–64 | partly cloudy | slight breeze | Absent | 18:00 | 23:00 | 35 | game farm |
| Safari Lodge, Victoria Falls, Zimbabwe | −17.92 | 25.82 | 18-Nov-16 | - | - | - | - | Absent | 18:00 | 22:45 | - | tourist lodge |
| Pafuri River Lodge | −22.37 | 31.52 | 19-Nov-16 | 2 | clear | no wind | Present | 18:00 | 23:15 | 200 | game farm | |
| Pafuri River Lodge | −22.37 | 31.52 | 20-Nov-16 | 2 | clear | strong wind | Present | 18:00 | 23:00 | 95 | game farm | |
| Wallacedale | −22.95 | 29.92 | 3-Dec-16 | 2 | 20–40 | clear | no wind | Present | 18:15 | 23:00 | 74 | homestead |
| Mumie 36NT | −22.43 | 30.31 | 24-Nov-17 | 4 | 50–62 | clear | slight breeze | Absent | 18:30 | 23:00 | - | - |
| Kineaiv-Smith | −22.87 | 29.81 | 24-Nov-17 | 2 | 30 | clear | no wind | Present | 18:30 | 23:00 | - | - |
| Princess Hill-1 | −22.86 | 29.81 | 24-Nov-17 | 2 | 30–40 | overcast | no wind | Absent | 18:15 | 23:15 | - | game farm |
| Princess Hill-2 | −22.87 | 29.81 | 24-Nov-17 | 2 | 50–55 | overcast | no wind | Absent | 18:15 | 23:30 | - | game farm |
| Venetia Nature Reserve (Faure Camp)) | −22.44 | 33.55 | 15-Nov-17 | 3 | 24–30 | clear | no wind | Present | 18:00 | 21:45 | - | nature reserve |
| Mapungubwe National Park (Picnic Baobab) | −22.20 | 29.38 | 16-Nov-17 | 3 | 24–30 | overcast | slight breeze | Present | 18:45 | 21:15 | - | world heritage site |
| Mapungubwe National Park (Camp Baobab) | −22.24 | 29.42 | 18-Nov-17 | 3 | 24–30 | clear | no wind | Present | 18:40 | 21:00 | - | world heritage site |
| Venetia Nature Reserve (Venetia Gate) | −22.26 | 29.32 | 23-Nov-17 | 5 | 20–54 | clear | strong breeze | Present | 18:30 | 23:15 | - | nature reserve |
| Mapungubwe National Park (Swing Baobab) | −22.20 | 29.37 | 17-Nov-17 | 3 | 24–30 | slight breeze | no wind | Present | 18:30 | 21:00 | - | world heritage site |
| Mapungubwe National Park (Camp Baobab) | −22.24 | 29.42 | 23-Nov-17 | 3 | 24–30 | overcast | strong wind | Present | 18:15 | 23:30 | - | world heritage site |
| Mapungubwe National Park (Zebra Pan) | −22.19 | 29.41 | 21-Nov-17 | 3 | 24–30 | clear | no wind | Present | 18:30 | 24:00 | - | world heritage site |
| Venetia Nature Reserve (Faure Camp) | −22.44 | 33.55 | 14-Nov-17 | 6 | 24–54 | clear | no wind | Present | 18:30 | 21:45 | - | nature reserve |
| Venetia Nature Reserve (Venetic Gate) | −22.26 | 29.32 | 20-Nov-17 | 3 | 24–30 | clear | no wind | Present | 18:45 | 23:30 | - | nature reserve |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taylor, P.J.; Vise, C.; Krishnamoorthy, M.A.; Kingston, T.; Venter, S. Citizen Science Confirms the Rarity of Fruit Bat Pollination of Baobab (Adansonia digitata) Flowers in Southern Africa. Diversity 2020, 12, 106. https://doi.org/10.3390/d12030106
Taylor PJ, Vise C, Krishnamoorthy MA, Kingston T, Venter S. Citizen Science Confirms the Rarity of Fruit Bat Pollination of Baobab (Adansonia digitata) Flowers in Southern Africa. Diversity. 2020; 12(3):106. https://doi.org/10.3390/d12030106
Chicago/Turabian StyleTaylor, Peter J., Catherine Vise, Macy A. Krishnamoorthy, Tigga Kingston, and Sarah Venter. 2020. "Citizen Science Confirms the Rarity of Fruit Bat Pollination of Baobab (Adansonia digitata) Flowers in Southern Africa" Diversity 12, no. 3: 106. https://doi.org/10.3390/d12030106
APA StyleTaylor, P. J., Vise, C., Krishnamoorthy, M. A., Kingston, T., & Venter, S. (2020). Citizen Science Confirms the Rarity of Fruit Bat Pollination of Baobab (Adansonia digitata) Flowers in Southern Africa. Diversity, 12(3), 106. https://doi.org/10.3390/d12030106