Factors Influencing the Distribution of Invasive Hybrid (Myriophyllum Spicatum x M. Sibiricum) Watermilfoil and Parental Taxa in Minnesota



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Field Sampling and Data Collection
2.3. Genetics
2.4. Data Analysis
3. Results
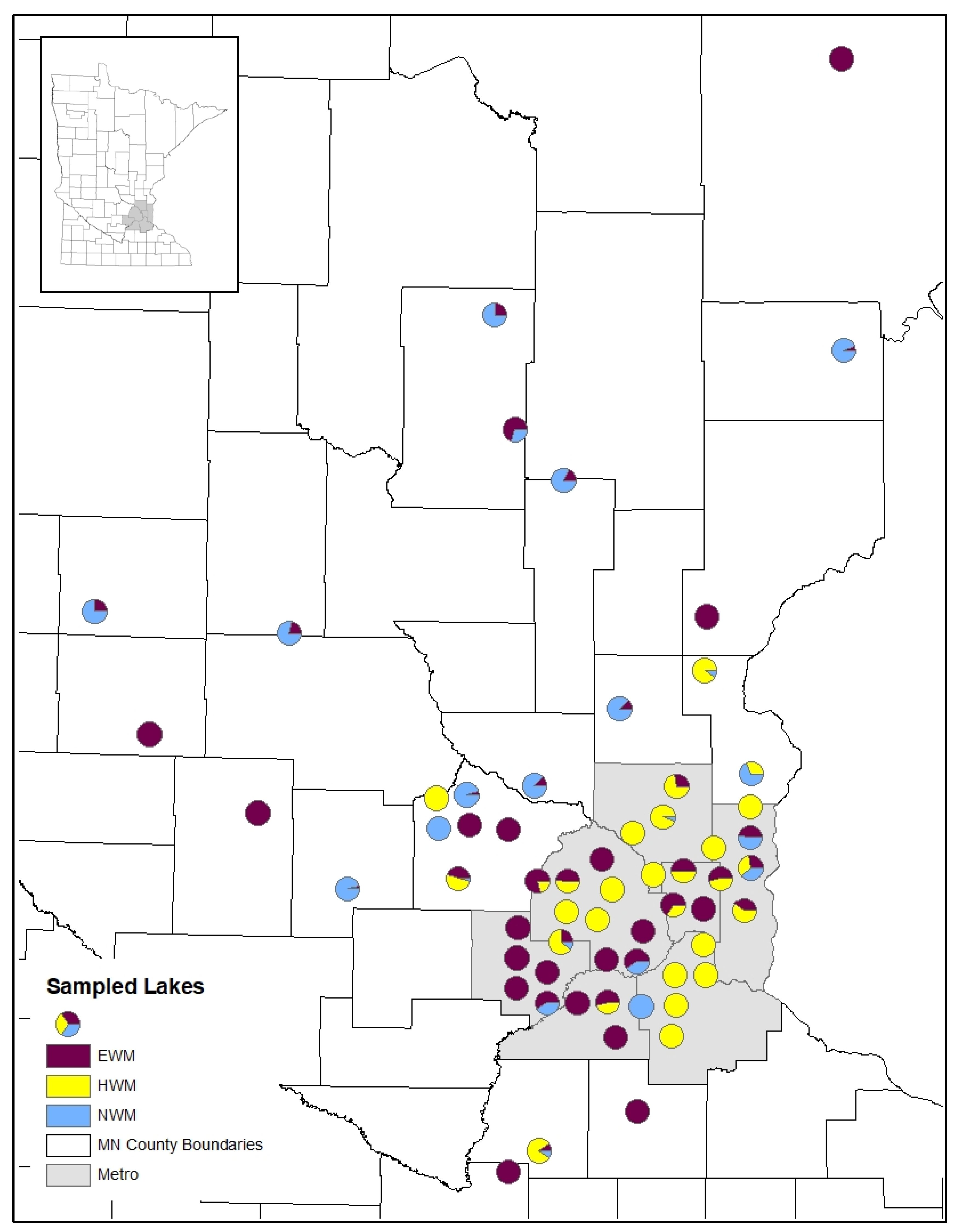
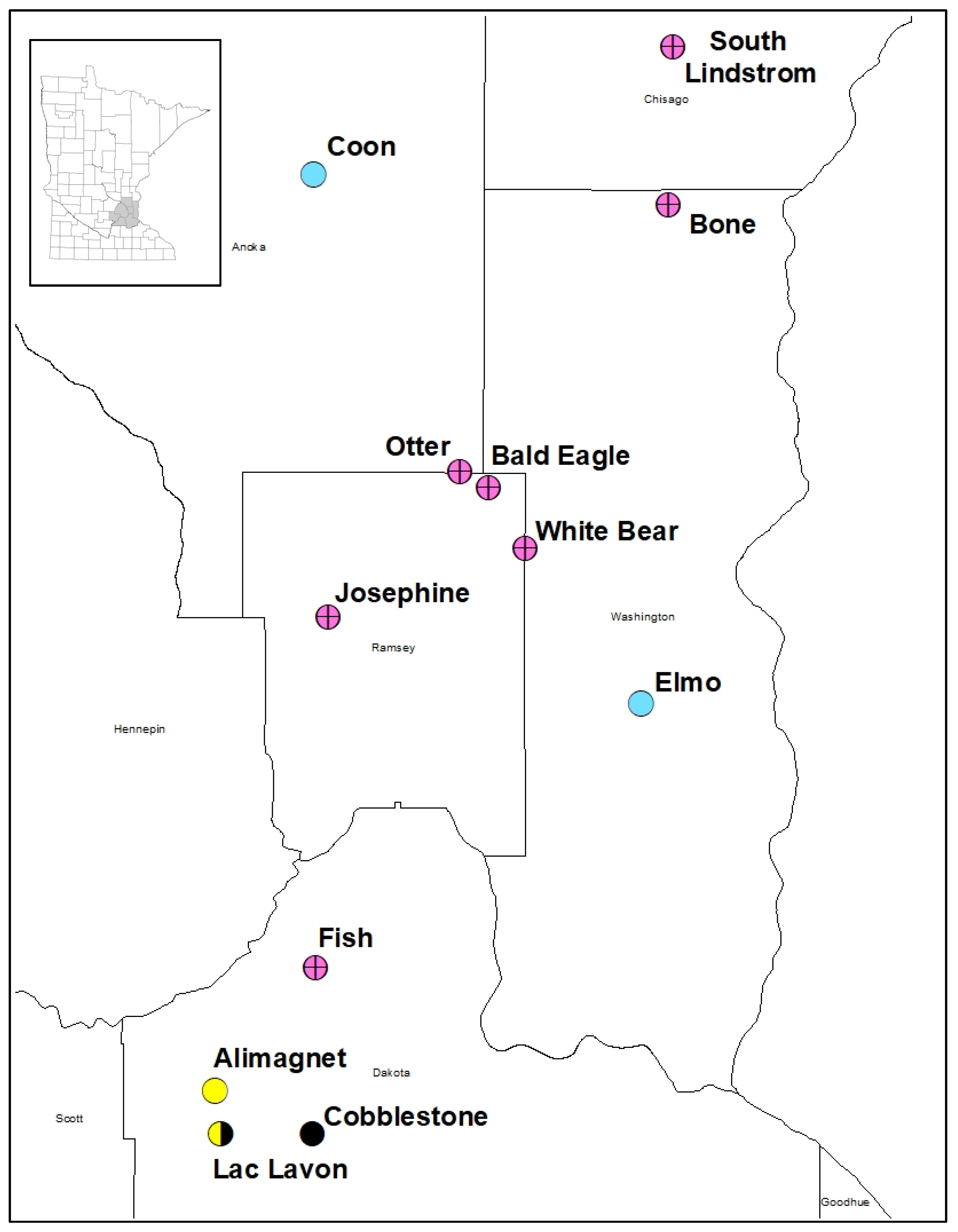
3.1. Watermilfoil Distribution
3.2. Watermilfoil Genotyping
3.3. Environmental Variables Analysis
3.4. Infestation-Associated VariablesAanalysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| County | Number of Infested Lakes | Proportion | Number Sampled | Proportion Sampled |
|---|---|---|---|---|
| * Hennepin | 46 | 0.144 | 11 | 0.177 |
| Wright | 40 | 0.125 | 7 | 0.113 |
| * Carver | 27 | 0.084 | 5 | 0.081 |
| * Ramsey | 27 | 0.084 | 6 | 0.097 |
| * Dakota | 22 | 0.069 | 5 | 0.081 |
| * Washington | 22 | 0.069 | 4 | 0.065 |
| * Anoka | 13 | 0.041 | 4 | 0.065 |
| Chisago | 12 | 0.038 | 2 | 0.032 |
| Crow Wing | 11 | 0.034 | 2 | 0.032 |
| * Scott | 10 | 0.031 | 2 | 0.032 |
| Meeker | 9 | 0.028 | 1 | 0.016 |
| Le Sueur | 8 | 0.025 | 1 | 0.016 |
| Rice | 7 | 0.022 | 1 | 0.016 |
| Itasca | 6 | 0.019 | 0 | 0.000 |
| Stearns | 6 | 0.019 | 0 | 0.000 |
| Kandiyohi | 5 | 0.016 | 1 | 0.016 |
| Sherburne | 5 | 0.016 | 1 | 0.016 |
| Blue Earth | 4 | 0.013 | 1 | 0.016 |
| Cass | 4 | 0.013 | 0 | 0.000 |
| Douglas | 4 | 0.013 | 1 | 0.016 |
| Isanti | 4 | 0.013 | 1 | 0.016 |
| Pine | 4 | 0.013 | 1 | 0.016 |
| Pope | 4 | 0.013 | 1 | 0.016 |
| Carlton | 3 | 0.009 | 1 | 0.016 |
| St. Louis | 3 | 0.009 | 1 | 0.016 |
| Morrison | 2 | 0.006 | 0 | 0.000 |
| Todd | 2 | 0.006 | 1 | 0.016 |
| Waseca | 2 | 0.006 | 0 | 0.000 |
| Winona | 2 | 0.006 | 0 | 0.000 |
| Kanabec | 1 | 0.003 | 0 | 0.000 |
| McLeod | 1 | 0.003 | 0 | 0.000 |
| Mille Lacs | 1 | 0.003 | 1 | 0.016 |
| Olmsted | 1 | 0.003 | 0 | 0.000 |
| Polk | 1 | 0.003 | 0 | 0.000 |
| Lake name | County | Lake ID | Lake Area (hectares) | Maximum Depth (m) | Year of Eurasian Infestation |
|---|---|---|---|---|---|
| Alimagnet | Dakota | 19-0021 | 41.9 | 3.5 | 2012 |
| Auburn | Carver | 10-004401 | 117.6 | 25.6 | 1989 |
| Bald Eagle | Ramsey | 62-0002 | 423.6 | 11.0 | 1989 |
| Ballantyne | Blue Earth | 07-0054 | 150.0 | 17.7 | 2012 |
| Bay | Crow Wing | 18-0034 | 938.8 | 22.6 | 1992 |
| Big Marine | Washington | 82-0052 | 728.1 | 18.9 | 2004 |
| Bone | Washington | 82-0054 | 89.6 | 9.1 | 2006 |
| Cedar | Hennepin | 27-0039 | 66.3 | 15.5 | 1990 |
| Cedar | Wright | 86-0227 | 319.8 | 32.9 | 2010 |
| Christmas | Hennepin | 27-0137 | 108.1 | 26.5 | 1992 |
| Chub | Carlton | 09-0008 | 126.8 | 8.5 | 2009 |
| Cobblestone | Dakota | 19-0456 | 14.1 | 5.5 | 2011 |
| Constance | Wright | 86-0051 | 70.7 | 7.0 | 2016 |
| Coon | Anoka | 02-0042 | 599.4 | 8.2 | 2003 |
| Crooked | Anoka | 02-0084 | 46.5 | 7.9 | 1990 |
| East Rush | Chisago | 13-006901 | 599.2 | 7.3 | 1992 |
| Elmo | Washington | 82-0106 | 103.9 | 42.7 | 2005 |
| Emily | Crow Wing | 18-0203 | 291.7 | 4.0 | 2014 |
| Fish | Dakota | 19-0057 | 12.4 | 10.2 | 2009 |
| Fox | Rice | 66-0029 | 126.1 | 14.3 | 2009 |
| German | Le Seuer | 40-0063 | 320.4 | 15.5 | 2002 |
| Gervais | Ramsey | 62-0007 | 95.1 | 12.5 | 1995 |
| Gilbert Pit | St. Louis | 69-1306 | 102.8 | 135.0 | 1999 |
| Gilchrist | Pope | 61-0072 | 136.0 | 7.3 | 1996 |
| Green | Kandiyohi | 34-0079 | 2250.3 | 33.5 | 2000 |
| Ham | Anoka | 02-0053 | 77.1 | 6.7 | 2013 |
| Harriet | Hennepin | 27-0016 | 138.1 | 26.5 | 1991 |
| Howard | Wright | 86-0199 | 301.5 | 11.9 | 2003 |
| Independence | Hennepin | 27-0176 | 342.7 | 17.7 | 1989 |
| Indian | Wright | 86-0223 | 56.4 | 9.5 | 2003 |
| Josephine | Ramsey | 62-0057 | 47.0 | 13.4 | 2012 |
| Lac Lavon | Dakota | 19-0446 | 26.7 | 9.8 | 1988 |
| Little Birch | Todd | 77-0089 | 339.7 | 27.1 | 2003 |
| Locke | Wright | 86-0168 | 56.7 | 14.9 | 2011 |
| McCarron | Ramsey | 62-0054 | 29.7 | 17.4 | 2000 |
| McMahon | Scott | 70-0050 | 65.7 | 4.3 | 2007 |
| Mille Lacs | Mille Lacs | 48-0002 | 51,891.3 | 12.8 | 1998 |
| Minnetonka Grays’ | Hennepin | 27-013301 | 74.6 | 11.0 | 1987 |
| Minnetonka North Arm | Hennepin | 27-013313 | 127.1 | 17.7 | 1987 |
| Minnetonka Smiths’ | Hennepin | 27-013302 | 184.1 | 9.1 | 1987 |
| Minnie-Belle | Meeker | 47-0119 | 240.2 | 14.9 | 2010 |
| Mitchell | Hennepin | 27-0070 | 46.1 | 5.8 | 2002 |
| Mitchell | Sherburne | 71-0081 | 68.6 | 10.1 | 2007 |
| Oscar | Douglas | 21-0257 | 471.7 | 7.6 | 1992 |
| Otter | Anoka | 02-0003 | 127.0 | 6.4 | 1989 |
| Phalen | Ramsey | 62-0013 | 80.0 | 27.7 | 1997 |
| Piersons | Carver | 10-0053 | 108.0 | 12.2 | 1991 |
| Pokegama | Pine | 58-0142 | 601.5 | 7.6 | 2005 |
| Rebecca | Hennepin | 27-0192 | 106.5 | 9.1 | 1989 |
| Riley | Carver | 10-0002 | 119.9 | 14.9 | 1990 |
| Schmidt | Hennepin | 27-0102 | 18.1 | 7.6 | 1990 |
| Somers | Wright | 86-0230 | 61.3 | 6.4 | 2013 |
| South Lindstrom | Chisago | 13-0028 | 184.0 | 10.4 | 2010 |
| Spectacle | Isanti | 30-0135 | 98.2 | 15.7 | 2007 |
| Staring | Hennepin | 27-0078 | 67.6 | 4.9 | 2015 |
| Steiger | Carver | 10-0045 | 67.1 | 11.3 | 2001 |
| Sugar | Wright | 86-0233 | 406.2 | 21.0 | 1990 |
| Swede | Carver | 10-0095 | 175.2 | 3.7 | 2008 |
| Thomas | Dakota | 19-0067 | 16.8 | 2.4 | 2011 |
| Turtle | Ramsey | 62-0061 | 182.1 | 8.5 | 2000 |
| Upper Prior | Scott | 70-0072 | 157.9 | 15.2 | 2000 |
| White Bear | Washington | 82-0167 | 982.5 | 25.3 | 1988 |
| Counts per Taxon | Genotype Counts per Lake | ||||||
|---|---|---|---|---|---|---|---|
| Lake | County | EWM | HWM | NWM | EWM | HWM | NWM |
| Alimagnet | Dakota | 20 | 1 | ||||
| Auburn | Carver | 24 | 1 | ||||
| Bald Eagle | Ramsey | 35 | 43 | 50 | 1 | 1 | 3 |
| Ballantyne | Blue Earth | 20 | 1 | ||||
| Bay | Crow Wing | 14 | 6 | 1 | 3 | ||
| Big Marine | Washington | 12 | 13 | 1 | 8 | ||
| Bone | Washington | 19 | 1 | ||||
| Cedar | Hennepin | 5 | 1 | ||||
| Cedar | Wright | 20 | 6 | ||||
| Christmas | Hennepin | 48 | 33 | 1 | 5 | ||
| Chub | Carlton | 1 | 19 | 1 | 1 | ||
| Cobblestone | Dakota | 2 | 1 | ||||
| Constance | Wright | 17 | 1 | ||||
| Coon | Anoka | 11 | 29 | 1 | 2 | ||
| Crooked | Anoka | 20 | 3 | ||||
| East Rush | Chisago | 18 | 2 | 1 | 1 | ||
| Elmo | Washington | 16 | 23 | 1 | 1 | ||
| Emily | Crow Wing | 2 | 6 | 1 | 6 | ||
| Fish | Dakota | 20 | 1 | ||||
| Fox | Rice | 20 | 2 | ||||
| German | Le Seuer | 1 | 9 | 1 | 1 | 5 | 1 |
| Gervais | Ramsey | ||||||
| Gilbert Pit | St. Louis | 9 | 1 | ||||
| Gilchrist | Pope | 20 | 1 | ||||
| Green | Kandiyohi | 2 | 1 | ||||
| Ham | Anoka | 97 | 6 | 1 | 1 | ||
| Harriet | Hennepin | 20 | 1 | ||||
| Howard | Wright | 9 | 10 | 1 | 1 | 6 | 1 |
| Independence | Hennepin | 43 | 44 | 1 | 1 | ||
| Indian | Wright | 1 | 1 | ||||
| Josephine | Ramsey | 19 | 1 | ||||
| Lac Lavon | Dakota | 20 | 5 | ||||
| Little Birch | Todd | 4 | 15 | 1 | 6 | ||
| Locke | Wright | ||||||
| McCarron | Ramsey | 21 | 11 | 1 | 1 | ||
| McMahon | Scott | 4 | 1 | ||||
| Mille Lacs | Mille Lacs | 2 | 10 | 1 | 2 | ||
| Minnetonka-Grays | Hennepin | 54 | 5 | ||||
| Minnetonka-North Arm | Hennepin | 20 | 7 | ||||
| Minnetonka-Smiths | Hennepin | 14 | 37 | 6 | 2 | 10 | 4 |
| Minnie-Belle | Meeker | 1 | 25 | 1 | 5 | ||
| Mitchell | Hennepin | 24 | 16 | 1 | 3 | ||
| Mitchell | Sherburne | 5 | 34 | 1 | 3 | ||
| Oscar | Douglas | 5 | 15 | 1 | 5 | ||
| Otter | Anoka | 64 | 2 | ||||
| Phalen | Ramsey | 4 | 1 | ||||
| Piersons | Carver | 19 | 1 | ||||
| Pokegama | Pine | 5 | 1 | ||||
| Rebecca | Hennepin | 21 | 8 | 1 | 1 | ||
| Riley | Carver | 21 | 1 | ||||
| Schmidt | Hennepin | 62 | 2 | ||||
| Somers | Wright | 2 | 1 | ||||
| South Lindstrom | Chisago | 9 | 19 | 1 | 4 | ||
| Spectacle | Isanti | 3 | 22 | 1 | 4 | ||
| Staring | Hennepin | 8 | 1 | ||||
| Steiger | Carver | 20 | 1 | ||||
| Sugar | Wright | 1 | 19 | 1 | 5 | ||
| Swede | Carver | 13 | 1 | ||||
| Thomas | Dakota | 5 | 2 | ||||
| Turtle | Ramsey | 6 | 6 | 1 | 1 | ||
| Upper Prior | Scott | 14 | 10 | 2 | 2 | ||
| White Bear | Washington | 24 | 12 | 1 | 1 | ||
References
- Galatowitsch, S.M.; Anderson, N.O.; Ascher, P.D. Invasiveness in wetland plants in temperate North America. Wetlands 1999, 19, 733–755. [Google Scholar] [CrossRef]
- Ellstrand, N.C.; Schierenbeck, K.A. Hybridization as a stimulus for the evolution of invasiveness in plants? Euphytica 2006, 97, 7043–7050. [Google Scholar] [CrossRef] [Green Version]
- Moody, M.L.; Les, D.H. Evidence of hybridity in invasive watermilfoil (Myriophyllum) populations. Proc. Natl. Acad. Sci. USA 2002, 99, 14867–14871. [Google Scholar] [CrossRef] [Green Version]
- Hovick, S.M.; Whitney, K.D. Hybridisation is associated with increased fecundity and size in invasive taxa: Meta-analytic support for the hybridisation-invasion hypothesis. Ecol. Lett. 2014, 17, 1464–1477. [Google Scholar] [CrossRef]
- Mitchell, N.; Campbell, L.G.; Ahern, J.R.; Paine, K.C.; Giroldo, A.B.; Whitney, K.D. Correlates of hybridization in plants. Evol. Lett. 2019, 3–6, 570–585. [Google Scholar] [CrossRef] [Green Version]
- Moody, M.L.; Les, D.H. Geographic distribution and genotypic composition of invasive hybrid watermilfoil (Myriophyllum spicatum x M. sibiricum) populations in North America. Biol. Invasions. 2007, 9, 559–570. [Google Scholar] [CrossRef]
- Schierenbeck, K.A.; Ellstrand, N.C. Hybridization and the evolution of invasiveness in plants and other organisms. Biol. Invasions. 2009, 11, 1093–1105. [Google Scholar] [CrossRef]
- Gaskin, J.F. The role of hybridization in facilitating tree invasion. AoB Plants 2016, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Thum, R.A.; Lennon, J.T.; Connor, J.; Smagula, A.P. A DNA fingerprinting approach for distinguishing among native and non-native milfoils. Lake Reserv. Manag. 2006, 22, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Wilcox, C.L.; Motomura, H.; Matsunuma, M.; Bowen, B.W. Phylogeography of Lionfishes (Pterois) indicate taxonomic over splitting and hybrid origin of the invasive Pterois volitans. J. Hered. 2018, 109, 162–175. [Google Scholar] [CrossRef] [Green Version]
- Moody, M.L.; Palomino, N.; Weyl, P.S.R.; Coetzee, J.A.; Newman, R.M.; Harms, N.E.; Liu, X.; Thum, R.A. Unraveling the biogeographic origins of the Eurasian watermilfoil (Myriophyllum spicatum) invasion in North America. Am. J. Bot. 2016, 103, 709–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- U.S. Geological Survey. Available online: https://nas.er.usgs.gov/queries/FactSheet.aspx?SpeciesID=237. (accessed on 18 December 2019).
- State of Minnesota, Department of Natural Resources. Invasive Species Program. In Invasive Species of Aquatic Plants and Wild Animals in MN: Annual Report for 2017; MN Department of Natural Resources: St. Paul, MN, USA, 2017. [Google Scholar]
- Madsen, J.D.; Sutherland, J.; Bloomfield, J.; Eichler, L.; Boylen, C. The decline of native vegetation under dense Eurasian watermilfoil canopies. J. Aquat. Plant Manag. 1991, 29, 94–99. [Google Scholar]
- Eiswerth, M.; Dowaldson, S.; Johnson, W. Potential environmental impacts and economic damages of Eurasian watermilfoil (Myriophyllum spicatum) in western Nevada and northeastern California. Weed Sci. 2000, 14, 511–518. [Google Scholar]
- Newroth, P.R. A review of Eurasian water milfoil impacts and management in British Columbia. Proceedings of the First International Symposium on Watermilfoil (Myriophyllum spicatum) and related Halorgaceae Species. Aquatic Plant Management Society: Vicksburg, MS, USA, 1985. In Proceedings of the First International Symposium on Watermilfoil (Myriophyllum spicatum) and related Halorgaceae Species, Vancouver, BC, Canada, 23–24 July 1985; Aquatic Plant Management Society: Vicksburg, MS, USA, 1985. [Google Scholar]
- Bartodziej, W.; Ludlow, J. Aquatic Vegetation Monitoring by Natural Resources Agencies in the United States. Lake Reserv. Manag. 1997, 13, 109–117. [Google Scholar] [CrossRef] [Green Version]
- LaRue, E.A.; Zuellig, M.P.; Netherland, M.D.; Heilman, M.A.; Thum, R.A. Hybrid watermilfoil lineages are more invasive and less sensitive to a commonly used herbicide than their exotic parent (Eurasian watermilfoil). Evol. Appl. 2013, 6, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Thum, R.T.; McNair, J.N. Inter- and intraspecific hybridization affects germination and vegetative growth in Eurasian watermilfoil. J. Aquat. Plant Manag. 2018, 56, 24–30. [Google Scholar]
- Nault, M.E.; Barton, M.; Hauxwell, J.; Heath, E.; Hoyman, T.; Mikulyuk, A.; Netherland, M.; Provost, S.; Skogerboe, J.; Van Egeren, S. Evaluation of large-scale low-concentration 2,4-D treatments for Eurasian and hybrid watermilfoil control across multiple Wisconsin lakes. Lake Reserv. Manag. 2018, 34, 115–129. [Google Scholar] [CrossRef]
- Parks, S.R.; McNair, J.N.; Hausler, P.; Tyning, P.; Thum, R.A. Divergent responses of cryptic invasive watermilfoil to treatment with auxinic herbicides in a large Michigan Lake. Lake Reserv. Manag. 2016, 32, 366–372. [Google Scholar] [CrossRef]
- Thum, R.T.; Parks, S.; McNair, J.N.; Tyning, P.; Hausler, P.; Chadderton, L.; Tucker, A.; Monfils, A. Survival and vegetative regrowth of Eurasian and hybrid watermilfoil following operational treatment with auxinic herbicides in Gun Lake, Michigan. J. Aquat. Plant Manag. 2017, 55, 103–107. [Google Scholar]
- Berger, S.T.; Netherland, M.D.; MacDonald, G.E. Evaluating fluridone sensitivity of multiple hybrid and Eurasian watermilfoil accessions under mesocosm conditions. J. Aquat. Plant Manag. 2012, 50, 135–146. [Google Scholar]
- Berger, S.T.; Netherland, M.D.; MacDonald, G.E. Laboratory Documentation of Multiple-Herbicide Tolerance to Fluridone, Norflurazon, and Topramazone in a Hybrid Watermilfoil (Myriophyllum spicatum × M. sibiricum) Population. Weed Sci. 2015, 63, 235–241. [Google Scholar] [CrossRef]
- Berger, S.T.; Netherland, M.D.; MacDonald, G.E. Development of a Rapid Assay to Detect Reduced Fluridone Sensitivity in Invasive Watermilfoils. Weed Technol. 2017, 29, 605–610. [Google Scholar] [CrossRef]
- Thum, R.A.; Heilman, M.A.; Hausler, P.J.; Huberty, L.E.; Tyning, P.; Wcisel, D.J.; Zuellig, M.P.; Berger, S.; Netherland, M.D. Field and laboratory documentation of reduced fluridone sensitivity by a hybrid watermilfoil biotype (Myriophyllum spicatum x Myriophyllum sibiricum). J. Aquat. Plant Manag. 2012, 50, 141–146. [Google Scholar]
- Berger, S.T. Characterization of a Suspected Herbicide Tolerant Hybrid Watermilfoil (Myriophyllum spicatum x M. sibiricum). Master’s Thesis, University of Florida, Gainesville, FL, USA, 2011. [Google Scholar]
- Sturtevant, A.P.; Hatley, N.; Pullman, G.D.; Sheick, R.; Shorez, D.; Bordine, A.; Mausolf, R.; Lewis, A.; Sutter, R.; Mortimer, A. Molecular characterization of Eurasian watermilfoil, northern watermilfoil, and the invasive interspecific hybrid in Michigan lakes. J. Aquat. Plant Manag. 2009, 47, 128–135. [Google Scholar]
- Borrowman, K.R.; Sager, E.P.; Thum, R.A. Distribution of biotypes and hybrids of Myriophyllum spicatum and associated Euhrychiopsis lecontei in lakes of Central Ontario, Canada. Lake Reserv. Manag. 2014, 30, 94–104. [Google Scholar] [CrossRef]
- Thum, R.A.; Chorak, G.; Newman, R.; Eltawely, J.; Latimore, J.; Elgin, E.; Parks, S. Genetic diversity and differentiation in populations of invasive Eurasian (Myriophyllum spicatum) and hybrid (M. spicatum x M. sibiricum) watermilfoil. Invas. Plant Sci. Manag. under review.
- Roley, S.S.; Newman, R.M. Predicting Eurasian watermilfoil invasions in MN. Lake Reserv. Manag. 2008, 24, 361–369. [Google Scholar] [CrossRef] [Green Version]
- Kanankege, K.S.T.; Alkhamis, M.A.; Perez, A.M.; Phelps, N.B.D. Zebra mussels and Eurasian watermilfoil reporting patterns in MN. J. Great Lakes Res. 2018, 44, 458–466. [Google Scholar] [CrossRef]
- Grafé, S.F.; Boutin, C.; Pick, F.R. A PCR-RFLP method to detect hybridization between the invasive Eurasian watermilfoil (Myriophyllum spicatum) and the native northern watermilfoil (Myriophyllum sibiricum), and its application in Ontario Lakes. Bot. 2015, 93, 117–121. [Google Scholar] [CrossRef]
- Wu, Z.; Ding, Z.; Yu, D.; Xu, X. Development of microsatellite markers in the hexaploidy aquatic macrophyte, Myriophyllum spicatum (Haloragaceae). Appl. Plant Sci. 2013, 2, 1–3. [Google Scholar]
- Clark, L.V.; Jasieniuk, M. POLYSAT: An R package for polyploid microsatellite analysis. Mol. Ecol. Resour. 2011, 11, 562–566. [Google Scholar] [CrossRef] [PubMed]
- French, A.; Macedo, M.; Poulsen, J.; Waterson, T.; Yu, A. Multivariate analysis of variance (MANOVA); San Francisco State University: San Francisco, CA, USA, 2008. [Google Scholar]
- Pashnick, J.; Thum, R.A. Comparison of molecular markers to distinguish genotypes of Eurasian watermilfoil, northern watermilfoil, and their hybrids. J. Aquat. Plant Manag. 2020, 58, 61–71. [Google Scholar]
- Zuellig, M.P.; Thum, R.A. Multiple introductions of invasive watermilfoil and recurrent hybridization with native northern watermilfoil in North America. J. Aquat. Plant Manag. 2012, 50, 1–19. [Google Scholar]
- Wu, Z.; Ding, Z.; Yu, D.; Xu, X. Influence of niche similarity on hybridization between Myriophyllum sibiricum and M. spicatum. J. Evol. Biol. 2015, 28, 1465–1475. [Google Scholar] [CrossRef] [PubMed]
- LaRue, E.A.; Grimm, D.; Thum, R.A. Laboratory crosses and genetic analysis of natural populations demonstrate sexual viability of invasive hybrid watermilfoils (Myriophyllum spicatum x M. sibiricum). Aquat. Bot. 2013, 109, 49–53. [Google Scholar] [CrossRef]
- Guo, Q. Plant Hybridization: The role of human disturbance and biological invasion. Divers. Distrib. 2014, 20, 1345–1354. [Google Scholar] [CrossRef]
- Taylor, L.L.; McNair, J.N.; Guastello, P.; Pashnick, J.; Thum, R.A. Heritable variation for vegetative growth rate in ten distinct genotypes of hybrid watermilfoil. J. Aquat. Plant Manag. 2017, 55, 51–57. [Google Scholar]
- Couch, R.; Nelson, E. Myriophllum spicatum in North America. In Proceedings of the First International Symposium on Watermilfoil (Myriophyllum spicatum) and Related Haloragaceae Species, Vancouver, BC, Canada; Anderson, L.J.W., Ed.; Aquatic Plant Management Society: Vicksburg, MS, USA, 1985; pp. 8–18. [Google Scholar]
- Eckert, C.G. The loss of sex in clonal plants. Evol. Ecol. 2002, 11, 501–520. [Google Scholar]
- Reynolds, C.; Nelson, N.A.F.; Cumming, G.S. The role of waterbirds in the dispersal of aquatic alien and invasive species. Divers. Distrib. 2015, 21, 744–754. [Google Scholar] [CrossRef] [Green Version]
- Nichols, S.A. Evaluation of invasions and declines of submersed macrophytes for the upper Great Lakes region. Lake Reserv. Manag. 1994, 10, 29–33. [Google Scholar] [CrossRef]
- Roshon, R.D.; McCann, J.H.; Thompson, D.G.; Stephenson, G.R. Effects of seven forestry management herbicides on Myriophyllum sibiricum, as compared with other nontarget aquatic organisms. Can. J. For. Res. 1999, 29, 1158–1169. [Google Scholar] [CrossRef]
- Valley, R.D.; Newman, R.M. Competitive interactions between Eurasian watermilfoil and northern watermilfoil in experimental tanks. J. Aquat. Plant Manag. 1998, 36, 121–126. [Google Scholar]


| EWM Only | HWM Only | NWM Only | EWM and HWM | NWM and HWM | EWM and NWM | All Three Taxa | Total | |
|---|---|---|---|---|---|---|---|---|
| Greater Minnesota | 8 | 1 | 1 | 0 | 2 | 10 | 2 | 24 |
| Metro area | 10 | 12 | 0 | 8 | 1 | 3 | 2 | 36 |
| Total | 18 | 13 | 1 | 8 | 3 | 13 | 4 | 60 |
| Taxon | Total Number of Unique Genotypes | Average Watermilfoil Genotype Richness/Lake |
|---|---|---|
| EWM | 6 | 2.5 ± 0.3 |
| HWM | 51 | 2.7 ± 0.3 |
| NWM | 81 | 3.5 ± 0.3 |
| Number of Lakes | Lake Area (ha) | Max Depth (m) | Secchi Depth (m) | Littoral Area (ha) | Trophic State Index | ||
|---|---|---|---|---|---|---|---|
| EWM | Statewide Metro area Greater Minnesota | 41 21 20 | 299 ± 62 231 ± 55 379 ± 116 | 17.5 ± 3.2 15.2 ± 2.1 20.4 ± 6.6 | 2.5 ± 0.3 2.4 ± 0.3 2.7 ± 0.4 | 159 ± 28 142 ± 35 178 ± 45 | 53 ± 2 54 ± 2 52 ± 2 |
| HWM | Statewide Metro area Greater Minnesota | 26 21 5 | 202 ± 45 164 ± 33 363 ± 186 | 12.3 ± 1.5 12.7 ± 1.9 10.9 ± 1.4 | 2.4 ± 0.2 2.5 ± 0.2 1.8 ± 0.4 | 122 ± 29 109 ± 32 174 ± 68 | 53 ± 1 52 ± 2 60 ± 1 |
| NWM | Statewide Metro area Greater Minnesota | 21 6 15 | 314 ± 52 261 ± 109 321 ± 60 | 14.3 ± 1.7 12.9 ± 3.3 15.6 ± 2.1 | 2.8 ± 0.3 3.0 ± 0.7 2.6 ± 0.3 | 177 ± 31 175 ± 71 167 ± 34 | 51 ± 2 49 ± 3 51 ± 2 |
| MANOVA p-value Taxon Region Taxon by Region | 0.519 0.134 0.908 | 0.399 0.420 0.755 | 0.566 0.765 0.568 | 0.649 0.409 0.898 | 0.562 0.398 0.229 |
| Coefficient | p-Value | |
|---|---|---|
| EWM Intercept Lake area Littoral area Maximum depth Secchi depth Trophic state index | −0.75 0.02 −0.02 0.05 −0.23 0.02 | 0.90 0.12 0.24 0.36 0.71 0.84 |
| HWM Intercept Lake area Littoral area Maximum depth Secchi depth Trophic state index | 0.62 −0.004 0.007 −0.04 0.18 −0.01 | 0.90 0.32 0.32 0.30 0.72 0.85 |
| NWM Intercept Lake area Littoral area Maximum depth Secchi depth Trophic state index | −2.72 −0.005 0.01 −0.05 0.92 0.002 | 0.61 0.09 * 0.04 * 0.07 * 0.11 0.976 |
| Number of Lakes | Age of Infestation (Years) | Parking Spaces at Water Access | Management Score | Distance to Nearest Infestation (km) | ||
|---|---|---|---|---|---|---|
| EWM | Statewide Metro area Greater Minnesota | 41 21 20 | 16.6 ± 1.3 19.7 ± 1.8 13.2 ± 1.7 | 22.0 ± 4.1 31.9 ± 7.1 11.5 ± 2.6 | 1.0 ± 0.1 0.9 ± 0.2 1.0 ± 0.2 | 20.8 ± 3.5 7.9 ± 1.3 34.1 ± 5.6 |
| HWM | Statewide Metro area Greater Minnesota | 26 21 5 | 19.2 ± 1.8 20.2 ± 2.1 15.0 ± 2.9 | 27.7 ± 6.4 29.5 ± 7.5 21.6 ± 10.9 | 1.2 ± 0.2 1.3 ± 0.2 1.2 ± 0.6 | 11.3 ± 2.2 7.2 ± 0.8 30.7 ± 7.1 |
| NWM | Statewide Metro area Greater Minnesota | 21 6 15 | 17.8 ± 1.9 21.2 ± 4.1 16.4 ± 2.1 | 23.0 ± 4.4 35.8 ± 8.5 17.8 ± 4.6 | 1.1 ± 0.2 1.0 ± 0.4 1.1 ± 0.3 | 29.4 ± 5.3 12.9 ± 3.0 36.0 ± 6.6 |
| MANOVA p-value Taxon Region Taxon by Region | 0.341 0.004 * 0.897 | 0.671 0.006 * 0.703 | 0.578 0.354 0.205 | 0.338 0.018 * 0.772 |
| Coefficient | p-Value | |
|---|---|---|
| EWM Intercept Region Age of infestation Parking spaces Management score Distance to nearest infestation | 0.43 1.21 0.01 0.01 −0.51 0.06 | 0.65 0.11 0.71 0.45 0.13 0.55 |
| HWM Intercept Region Age of infestation Parking spaces Management score Distance to nearest infestation | 0.04 −1.71 0.01 0.01 0.31 −0.07 | 0.97 0.01 * 0.73 0.44 0.34 0.43 |
| NWM Intercept Region Age of infestation Parking spaces Management score Distance to nearest infestation | −2.10 2.48 0.01 0.02 −0.08 −0.02 | 0.04 * 0.001 * 0.78 0.19 0.81 0.63 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eltawely, J.A.; Newman, R.M.; Thum, R.A. Factors Influencing the Distribution of Invasive Hybrid (Myriophyllum Spicatum x M. Sibiricum) Watermilfoil and Parental Taxa in Minnesota. Diversity 2020, 12, 120. https://doi.org/10.3390/d12030120
Eltawely JA, Newman RM, Thum RA. Factors Influencing the Distribution of Invasive Hybrid (Myriophyllum Spicatum x M. Sibiricum) Watermilfoil and Parental Taxa in Minnesota. Diversity. 2020; 12(3):120. https://doi.org/10.3390/d12030120
Chicago/Turabian StyleEltawely, Jasmine A., Raymond M. Newman, and Ryan A. Thum. 2020. "Factors Influencing the Distribution of Invasive Hybrid (Myriophyllum Spicatum x M. Sibiricum) Watermilfoil and Parental Taxa in Minnesota" Diversity 12, no. 3: 120. https://doi.org/10.3390/d12030120
APA StyleEltawely, J. A., Newman, R. M., & Thum, R. A. (2020). Factors Influencing the Distribution of Invasive Hybrid (Myriophyllum Spicatum x M. Sibiricum) Watermilfoil and Parental Taxa in Minnesota. Diversity, 12(3), 120. https://doi.org/10.3390/d12030120