Knowing the Enemy: Inducible Defences in Freshwater Zooplankton



Abstract
:1. Introduction
2. Inducible Defences
2.1. Behavioural Defences
2.2. Morphological Defences
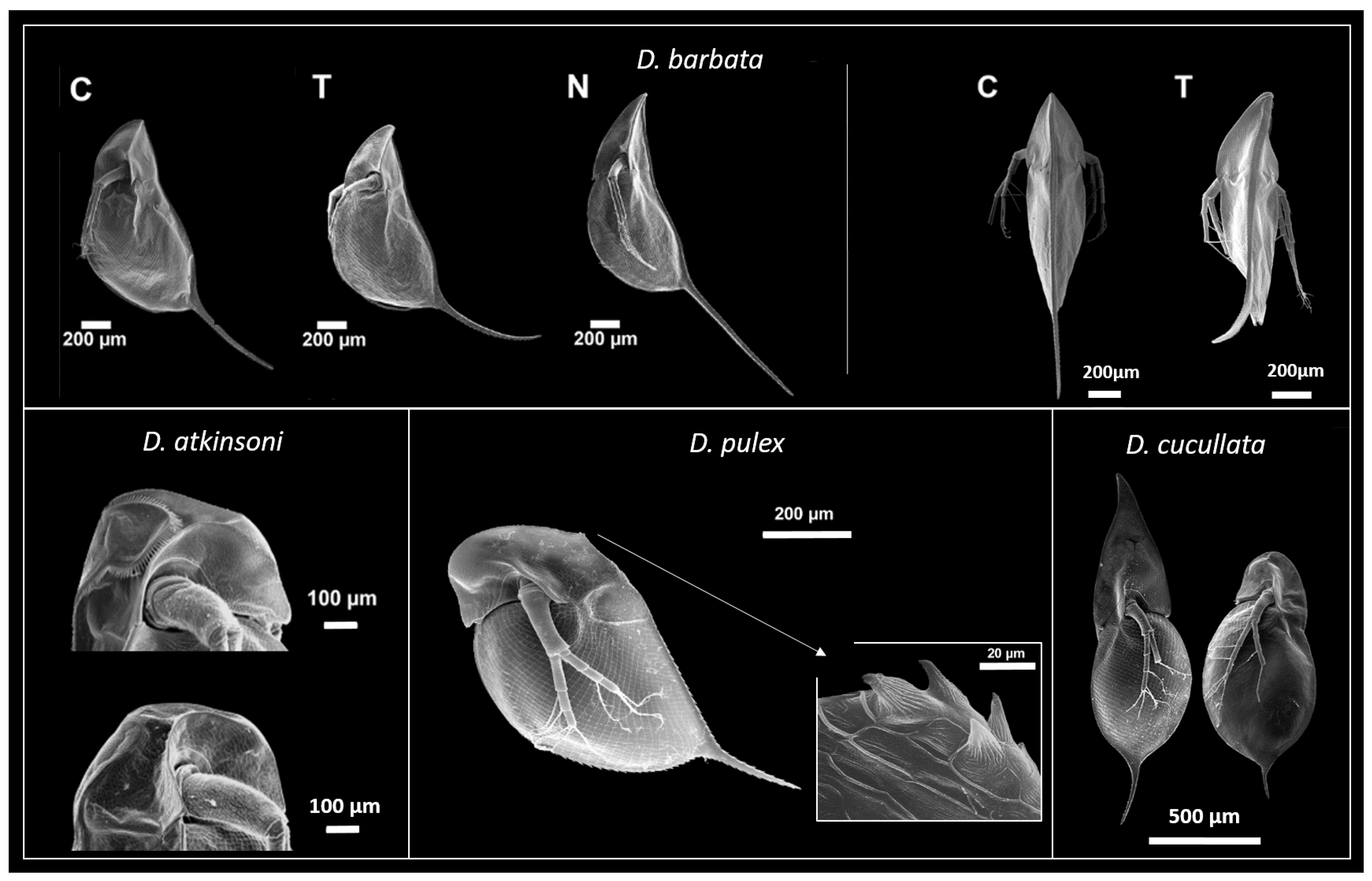
2.2.1. Defences against Vertebrate Predators
2.2.2. Defences against Invertebrate Predators
2.3. Life-History Defences
2.4. Conceptual Approaches for Multipredator Studies
3. Costs for Inducible Defence
4. Defence-Inducing Cues
5. Conclusions and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
| Zooplankton Species | Predator | Behaviour | Morphology | Life-History | Single Clone | Multiple Clones | Field Study | Laboratory Study | Source |
|---|---|---|---|---|---|---|---|---|---|
| Argyrodiaptomus falcifer | Cnesterodon decemmaculatus | x | n.a. | n.a. | x | [58] | |||
| Bestiolina similis | Amphiprion ocellaris | x | n.a. | n.a. | x | [239] | |||
| Bosmina coregoni | Fish | x | n.a. | n.a. | x | [80] | |||
| Bosmina longirostris | Acanthocyclops verinalis | x | x | x | [238] | ||||
| Copepoda | x | x | n.a. | n.a. | x | [106] | |||
| Coregonus alba | x | x | n.a. | n.a. | x | [117] | |||
| Craspedacusta sowerbii | x | x | n.a. | n.a. | x | [168] | |||
| Fish | x | n.a. | n.a. | x | [80] | ||||
| Leptodora kindtii | x | x | n.a. | n.a. | x | [164] | |||
| x | x | n.a. | n.a. | x | [13,106] | ||||
| Mesocyclops leuckarti | x | x | x | x | [95] | ||||
| Mesocyclops sp. | x | x | x | [170] | |||||
| Bosmina fatalis | Copepoda | x | x | n.a. | n.a. | x | [106] | ||
| Leptodora kindtii | x | x | n.a. | n.a. | x | [164] | |||
| x | x | n.a. | n.a. | x | [13,106] | ||||
| Mesocyclops leuckarti | x | x | x | x | [95] | ||||
| Mesocyclops sp. | x | x | x | [170] | |||||
| Bosmina sp. | Fish | x | n.a. | n.a. | x | [54] | |||
| Mochlonyx fuliginosus | x | n.a. | n.a. | x | [54] | ||||
| Brachionus calyciflorus | Asplanchna brightwelli | x | x | x | [179] | ||||
| x | x | n.a. | n.a. | x | [178] | ||||
| Asplanchna sieboldi | x | x | x | [177] | |||||
| Asplanchna sp. | x | x | x | [233] | |||||
| Daphnia pulex | x | x | x | [179] | |||||
| Brachionus havaensis | Asplanchna sieboldi | x | x | x | [177] | ||||
| Brachionus macracanthus | Asplanchna sieboldi | x | x | x | [177] | ||||
| Brachionus patulus | Asplanchna sieboldi | x | x | x | [177] | ||||
| Brachionus rubens | Asplanchna sieboldi | x | x | x | [177] | ||||
| Brachionus variabilis | Asplanchna girodi | x | x | x | x | [92] | |||
| Daphnia pulex | x | x | x | x | [92] | ||||
| Chydorus Sphaericus | Various | x | x | x | [80] | ||||
| Ceriodaphnia cornuta | Aeolosoma sp. | x | x | x | x | [102] | |||
| Skiffia lermae | x | x | x | x | [102] | ||||
| Ceriodaphnia dubia | Aeolosoma sp. | x | x | x | x | [102] | |||
| Chaoborus sp. | x | n.a. | n.a. | x | [91] | ||||
| Cnesterodon decemmaculatus | x | n.a. | n.a. | x | [58] | ||||
| Macrobrachium borellii | x | n.a. | n.a. | x | [91] | ||||
| Melanotaenia duboulayi | x | n.a. | n.a. | x | [98] | ||||
| Skiffia lermae | x | x | x | x | [102] | ||||
| Trichodactylus borellianus | x | n.a. | n.a. | x | [91] | ||||
| Ceriodaphnia reticulata | Mesocyclops leuckarti | x | x | x | x | [95] | |||
| Rutilus rutilus | x | x | x | x | [81] | ||||
| Copepoda | Gambusia hubbsi | x | n.a. | n.a. | x | [33] | |||
| Rutilus rutilus | x | x | n.a. | n.a. | x | [65] | |||
| Cyclops abyssorum | Fish | x | n.a. | n.a. | x | [48] | |||
| Daphnia ambigua | Chaoborus sp. | x | x | x | [143] | ||||
| Daphnia atkinsoni | Triops cancriformis | x | x | x | [154,155] | ||||
| Daphnia barbata | Notonecta glauca | x | x | x | [28] | ||||
| Triops cancriformis | x | x | x | [28,157] | |||||
| Daphnia cucullata | Chaoborus flavicans | x | x | x | [22,223,233] | ||||
| Chaoborus sp. | x | x | x | [143] | |||||
| Cyclops sp. | x | x | x | [202,223] | |||||
| Leptodora kindtii | x | x | x | [202] | |||||
| Fish | x | n.a. | n.a. | x | [80] | ||||
| Phoxinus phoxinus | x | x | x | [66] | |||||
| Rutilus rutilus | x | x | x | [205] | |||||
| Daphnia curvirostris | Chaoborus crystallinus | x | x | x | [107] | ||||
| Daphnia dentifera | Chaoborus punctipennis | x | x | x | [212] | ||||
| Lepomis macrochirus | x | x | x | [212] | |||||
| Daphnia galeata | Chaoborus sp. | x | x | x | x | [195] | |||
| Gasterosteus aculeatus | x | x | x | [222] | |||||
| Fish | x | n.a. | n.a. | x | [54,60] | ||||
| Lepomis macrochirus | x | x | x | [101] | |||||
| Leucaspius delineatus | x | x | x | [60] | |||||
| Leuciscus idus | x | x | x | x | [4] | ||||
| x | x | x | x | [195] | |||||
| Mochlonyx fuliginosus | x | n.a. | n.a. | x | [54] | ||||
| Perca fluviatilis | x | x | x | [72] | |||||
| x | x | x | [222] | ||||||
| Rhodeus sericeus amarus | x | x | x | [213] | |||||
| Scardinius erythrophthalmus | x | x | x | [203] | |||||
| Daphnia hrbaceki | Chaoborus crystallinus | x | x | x | [107] | ||||
| Daphnia hyalina | Chaoborus sp. | x | x | x | x | [195] | |||
| Leuciscus idus | x | x | x | x | [195] | ||||
| Perca fluviatilis | x | x | x | [72] | |||||
| Poxinus phoxinus | x | x | x | [66] | |||||
| Rutilus rutilus | x | x | x | [205] | |||||
| Daphnia hyalina x galeata | Chaoborus sp. | x | x | x | x | [195] | |||
| Fish | x | x | x | [79] | |||||
| Leuciscus idus | x | x | x | x | [195] | ||||
| Perca fluviatilis | x | x | x | [59,72] | |||||
| x | x | x | [199] | ||||||
| Rutilus rutilus | x | x | x | x | [81] | ||||
| x | x | x | [210] | ||||||
| Rhodeus sericeus amarus | x | x | x | [213] | |||||
| Daphnia laevis | Oreochromis spp. | x | x | x | x | [45] | |||
| Daphnia longicephala | Notonecta glauca | x | x | x | [22,233] | ||||
| x | x | x | x | [248] | |||||
| Notonecta sp. | x | x | x | [143,245] | |||||
| Daphnia longispina | Various | x | x | x | [80] | ||||
| Coregonus alba | x | x | n.a. | n.a. | x | [117] | |||
| Chaoborus flavicans | x | n.a. | n.a. | x | [144] | ||||
| Chaoborus sp. | x | x | x | x | [145] | ||||
| Fish | x | n.a. | n.a. | x | [80] | ||||
| Gambusia holbrooki | x | x | x | [220] | |||||
| Lepomis gibbosus | x | x | x | [220] | |||||
| Notonecta glauca | x | x | x | [30] | |||||
| Scardinius erythrophthalmus | x | x | x | x | [240] | ||||
| Daphnia lumholtzi | Chaoborus flavicans | x | x | x | [131] | ||||
| Chaoborus punctipennis | x | x | x | x | [116] | ||||
| Gasterosteus aculeatus | x | x | x | [131] | |||||
| x | x | x | [233] | ||||||
| Lepomis macrochirus | x | x | x | x | [116] | ||||
| Leucaspius delineatus | x | x | x | x | [132] | ||||
| Phoxinus phoxinus | x | x | x | [22,143] | |||||
| Daphnia magna | Abramis brama | x | x | x | [96] | ||||
| Carassius carassius | x | x | x | [192,193] | |||||
| x | x | x | [232] | ||||||
| Chaoborus crystallinus | x | x | x | [241] | |||||
| x | x | x | [96] | ||||||
| Chaoborus obscuripes | x | x | x | [241] | |||||
| Danio rerio | x | x | x | [96] | |||||
| Fish | x | x | x | x | x | [124] | |||
| x | x | x | [188] | ||||||
| Gambusia holbrooki | x | x | x | [183] | |||||
| Gasterosteus aculeatus | x | x | x | x | [99] | ||||
| Ischnura elegans | x | n.a. | n.a. | x | [88] | ||||
| Lepomis gibbosus | x | x | x | [208] | |||||
| Leucaspius delineatus | x | x | x | [96] | |||||
| x | x | x | [189,194] | ||||||
| Leuciscus idus | x | x | x | [221] | |||||
| x | x | x | [190,206] | ||||||
| Notonecta glauca | x | x | x | [183] | |||||
| Notonecta sp. | x | x | x | [96] | |||||
| Perca fluviatilis | x | x | x | [119,120] | |||||
| x | x | x | [72] | ||||||
| Pseudorasbora parva | x | n.a. | n.a. | x | [90] | ||||
| Rhodeus amarus | x | x | x | [190] | |||||
| x | x | [214] | |||||||
| Rutilus rutilus | x | x | x | [227] | |||||
| Salmo trutta | x | x | x | x | [23] | ||||
| x | x | x | [236] | ||||||
| Triops cancriformis | x | x | x | [156,158] | |||||
| x | x | x | [108] | ||||||
| x | x | x | [191] | ||||||
| x | x | x | [211,214] | ||||||
| Daphnia mendotae | Bythotrephes longimanus | x | x | x | [161] | ||||
| x | x | n.a. | n.a. | x | [55] | ||||
| Chaoborus punctipennis | x | n.a. | n.a. | x | [201] | ||||
| Lepomis macrochirus | x | x | x | [181] | |||||
| Leptodora kindtii | x | n.a. | n.a. | x | [201] | ||||
| Perca flavescens | x | x | n.a. | n.a. | x | [82] | |||
| Daphnia middendorffiana | Phoxinus phoxinus | x | x | x | [66] | ||||
| Daphnia minnehaha | Chaoborus americanus | x | x | x | [27] | ||||
| Chaoborus trivitattus | x | x | x | [27] | |||||
| Daphnia obtusa | Micropterus salmoides | x | x | x | [101] | ||||
| Daphnia pulex | Alburnus alburnus | x | x | x | [49,218,219] | ||||
| x | n.a. | n.a. | x | [49] | |||||
| Alburnus escherichi | x | n.a. | n.a. | x | [246] | ||||
| Chaoborus americanus | x | x | x | [56] | |||||
| x | x | x | [197,204] | ||||||
| x | x | x | [243,247] | ||||||
| x | x | x | [27] | ||||||
| x | x | x | x | [84] | |||||
| x | x | x | x | [244] | |||||
| Chaoborus crystallinus | x | x | x | [107] | |||||
| Chaoborus flavicans | x | x | x | [196] | |||||
| x | x | x | [44] | ||||||
| x | x | x | x | x | [42] | ||||
| x | x | x | [14,148,233] | ||||||
| x | n.a. | n.a. | x | [144] | |||||
| x | x | x | [44] | ||||||
| x | x | x | [109] | ||||||
| x | x | x | x | [198] | |||||
| x | x | x | [211,215] | ||||||
| Chaoborus obscuripes | x | x | x | [30] | |||||
| Chaoborus sp. | x | x | x | [143,245] | |||||
| Chaoborus trivitattus | x | x | x | [44] | |||||
| x | x | x | x | x | [42] | ||||
| x | x | x | x | [182] | |||||
| x | x | x | [27] | ||||||
| x | x | x | [44] | ||||||
| x | x | x | x | [84] | |||||
| Gasterosteus aculeatus | x | x | x | [196] | |||||
| Mesocyclops leuckarti | x | x | x | x | [95] | ||||
| Notemigonus crysoleucas | x | x | x | [44] | |||||
| x | x | x | x | x | [42] | ||||
| x | x | x | [44] | ||||||
| Phoxinus phoxinus | x | x | x | [66] | |||||
| Scardinius erythrophthalmus | x | x | x | [203] | |||||
| Daphnia pulicaria | Chaoborus flavicans | x | x | x | x | x | [42] | ||
| Chaoborus sp. | x | x | x | [242] | |||||
| x | x | x | [97] | ||||||
| Chaoborus trivitattus | x | x | x | x | x | [42] | |||
| Lepomis macrochirus | x | x | x | [207] | |||||
| x | x | x | [181] | ||||||
| x | n.a. | n.a. | x | [57] | |||||
| x | x | x | [97] | ||||||
| Leucaspius delineatus | x | x | x | x | [132] | ||||
| Notemigonus crysoleucas | x | x | x | x | x | [42] | |||
| Perca flavescens | x | x | n.a. | n.a. | x | [82] | |||
| Rutilus rutilus | x | x | x | [210] | |||||
| Daphnia similis | Chaoborus obscuripes | x | x | x | [184] | ||||
| Gasterosteus aculeatus | x | x | x | [184] | |||||
| Lepomis macrochirus | x | x | x | [101] | |||||
| Oreochromis spp. | x | x | x | x | [45] | ||||
| Daphnia similoides sinensis | Cyclops vicinus | x | n.a. | n.a. | x | [123] | |||
| Leptodora richardi | x | n.a. | n.a. | x | [123] | ||||
| Neosalanx taihuensis | x | n.a. | n.a. | x | [123] | ||||
| Daphnia spinulata | Oreochromis spp. | x | x | x | x | [45] | |||
| Daphnia sp. | Coregonus alba | x | x | n.a. | n.a. | x | [117] | ||
| Lepomis macrochirus | x | n.a. | n.a. | x | [61] | ||||
| Rutilus rutilus | x | n.a. | n.a. | x | [83] | ||||
| Diaphanosoma brachyurum | Fish | x | n.a. | n.a. | x | [54,80] | |||
| Mesocyclops leuckarti | x | x | x | x | [95] | ||||
| Mochlonyx fuliginosus | x | n.a. | n.a. | x | [54] | ||||
| Various | x | x | x | [80] | |||||
| Diaphanosoma birgei | Hyphessobrycon bifasciatus | x | x | x | [187] | ||||
| Eubosmina coregoni | Various | x | x | x | [80] | ||||
| Eubosmina coregoni coregoni | Mesocyclops leuckarti | x | x | x | [169] | ||||
| Eubosmina coregoni gibbera | Mesocyclops leuckarti | x | x | x | [169] | ||||
| Eubosmina coregoni kessleri | Mesocyclops leuckarti | x | x | x | [169] | ||||
| Eubosmina coregoni thersites | Mesocyclops leuckarti | x | x | x | [169] | ||||
| Eubosmina longispina | Coregonus alba | x | x | n.a. | n.a. | x | [117] | ||
| Eudiaptomus gracilis | Fish | x | n.a. | n.a. | x | [48] | |||
| Keratella tropica | Asplanchna brightwelli | x | x | n.a. | n.a. | x | [176] | ||
| x | n.a. | n.a. | x | [179] | |||||
| Daphnia pulex | x | x | n.a. | n.a. | x | [176] | |||
| x | n.a. | n.a. | x | [179] | |||||
| Moina macrocopa | Chaoborus obscuripes | x | x | x | [184] | ||||
| Gasterosteus aculeatus | x | x | x | [184] | |||||
| Moina micrura | Gasterosteus aculeatus | x | x | x | [187] | ||||
| Hyphessobrycon bifasciatus | x | x | x | [187] | |||||
| Neomysis integer | Perca fluviatilis | x | n.a. | n.a. | x | [89] | |||
| Notodiaptomus conifer | Chaoborus sp. | x | n.a. | n.a. | x | [91] | |||
| Cnesterodon decemmaculatus | x | n.a. | n.a. | x | [58] | ||||
| Macrobrachium borellii | x | n.a. | n.a. | x | [91] | ||||
| Trichodactylus borellianus | x | n.a. | n.a. | x | [91] | ||||
| Plationus macracanthus | Asplanchna brightwellii | x | x | n.a. | n.a. | x | [178] | ||
| Praunus flexuosus | Perca fluviatilis | x | n.a. | n.a. | x | [89] | |||
| Pseudosida variabilis | Cnesterodon decemmaculatus | x | n.a. | n.a. | x | [58] | |||
| Simocephalus serrulatus | Aeolosoma sp. | x | x | x | x | [102] | |||
| Skiffia lermae | x | x | x | x | [102] | ||||
| Simocephalus vetulus | Aeolosoma sp. | x | x | x | x | [102] | |||
| Skiffia lermae | x | x | x | x | [102] | ||||
| Various zooplankton | Chaoborus flavicans | x | n.a. | n.a. | x | [26] |
References
- Werner, E.E.; Peacor, S.D. A review of trait-mediated indirect interactions in ecological communities. Ecology 2003, 84, 1083–1100. [Google Scholar] [CrossRef] [Green Version]
- Boaden, A.E.; Kingsford, M.J. Predators drive community structure in coral reef fish assemblages. Ecosphere 2015, 6, 1–33. [Google Scholar] [CrossRef]
- Kuchta, S.; Svensson, E.I. Predator-Mediated Natural Selection on the Wings of the Damselfly Calopteryx splendens: Differences in Selection among Trait Types. Am. Nat. 2014, 184, 91–109. [Google Scholar] [CrossRef] [Green Version]
- Tams, V.; Lüneburg, J.; Seddar, L.; Detampel, J.P.; Cordellier, M. Intraspecific phenotypic variation in life history traits of Daphnia galeata populations in response to fish kairomones. PeerJ 2018, 6, e5746. [Google Scholar] [CrossRef] [Green Version]
- Karban, R.; Baldwin, I.T. Induced Responses to Herbivory; University of Chicago Press: Chicago, IL, USA, 2013; Volume 1, pp. 1–330. [Google Scholar]
- Agrawal, A.A. Agrawal Induced responses to herbivory and increased plant performance. Science 1998, 279, 1201–1202. [Google Scholar] [CrossRef] [Green Version]
- Tollrian, R.; Harvell, C.D. The Ecology and Evolution of Induced Defenses, 1st ed.; Princeton University Press: Princeton, NJ, USA, 1999; Volume 1, pp. 1–377. [Google Scholar]
- Dicke, M.; Hilker, M. Induced plant defences: From molecular biology to evolutionary ecology. Basic Appl. Ecol. 2003, 4, 3–14. [Google Scholar] [CrossRef]
- Jeschke, J.M.; Laforsch, C.; Tollrian, R. Animal Prey Defenses. In Encyclopedia of Ecology; Elsevier: Amsterdam, The Netherlands, 2008; pp. 189–194. ISBN 9780080914565. [Google Scholar]
- Spitze, K.; Sadler, T.D. Evolution of a generalist genotype: Multivariate analysis of the adaptiveness of phenotypic plasticity. In Proceedings of the American Naturalist; The University of Chicago Press: Chicago, IL, USA, 1996; Volume 148, pp. 108–123. [Google Scholar]
- Stearns, S.C. The Evolutionary Significance of Phenotypic Plasticity. Bioscience 1989, 39, 436–445. [Google Scholar] [CrossRef] [Green Version]
- Laforsch, C.; Tollrian, R. Cyclomorphosis and Phenotypic Changes. In Encyclopedia of Inland Waters; Elsevier: Amsterdam, The Netherlands, 2009; pp. 643–650. ISBN 9780123706263. [Google Scholar]
- Sakamoto, M.; Nagata, T.; Ha, J.Y.; Kimijima, S.; Hanazato, T.; Chang, K.H. Inducible defenses as factor determining trophic pathways in a food web. Hydrobiologia 2015, 743, 15–25. [Google Scholar] [CrossRef]
- Weiss, L.C.; Albada, B.; Becker, S.M.; Meckelmann, S.W.; Klein, J.; Meyer, M.; Schmitz, O.J.; Sommer, U.; Leo, M.; Zagermann, J.; et al. Identification of Chaoborus kairomone chemicals that induce defences in Daphnia. Nat. Chem. Biol. 2018, 14, 1133–1139. [Google Scholar] [CrossRef]
- Brancelj, A.; Čelhar, T.; Šiško, M. Four different head shapes in Daphnia hyalina (Leydig) induced by the presence of larvae of Chaoborus flavicans (Meigen). Hydrobiologia 1996, 339, 37–45. [Google Scholar] [CrossRef]
- Kats, L.B.; Dill, L.M. The scent of death: Chemosensory assessment of predation risk by prey animals. Ecoscience 1998, 5, 361–394. [Google Scholar] [CrossRef]
- Lass, S.; Spaak, P. Chemically induced anti-predator defences in plankton: A review. In Proceedings of the Hydrobiologia; Springer: Berlin/Heidelberg, Germany, 2003; Volume 491, pp. 221–239. [Google Scholar]
- Korosi, J.B.; Kurek, J.; Smol, J.P. A review on utilizing Bosmina size structure archived in lake sediments to infer historic shifts in predation regimes. J. Plankton Res. 2013, 35, 444–460. [Google Scholar] [CrossRef] [Green Version]
- Riessen, H.P.; Gilbert, J.J. Divergent developmental patterns of induced morphological defenses in rotifers and Daphnia: Ecological and evolutionary context. Limnol. Oceanogr. 2019, 64, 541–557. [Google Scholar] [CrossRef]
- Brown, W.L.; Eisner, T.; Whittaker, R.H. Allomones and Kairomones: Transspecific Chemical Messengers. Bioscience 1970, 20, 21–22. [Google Scholar] [CrossRef]
- Dodson, S.I. The ecological role of chemical stimuli for the zooplankton: Predator-induced morphology in Daphnia. Oecologia 1989, 78, 361–367. [Google Scholar] [CrossRef]
- Laforsch, C.; Beccara, L.; Tollrian, R. Inducible defenses: The relevance of chemical alarm cues in Daphnia. Limnol. Oceanogr. 2006, 51, 1466–1472. [Google Scholar] [CrossRef] [Green Version]
- Pestana, J.L.T.; Baird, D.J.; Soares, A.M.V.M. Predator threat assessment in Daphnia magna: The role of kairomones versus conspecific alarm cues. Mar. Freshw. Res. 2013, 64, 679–686. [Google Scholar] [CrossRef]
- Brooks, J.L.; Dodson, S.I. Predation, body size, and composition of plankton. Science 1965, 150, 28–35. [Google Scholar] [CrossRef]
- Kerfoot, W.C.; Weider, L.J. Experimental paleoecology (resurrection ecology): Chasing Van Valen’s Red Queen hypothesis. Limnol. Oceanogr. 2004, 49, 1300–1316. [Google Scholar] [CrossRef] [Green Version]
- Lagergren, R.; Leberfinger, K.; Stenson, J.A.E. Seasonal and ontogenetic variation in diel vertical migration of Chaoborus flavicans and its effect on depth-selection behavior of other zooplankton. Limnol. Oceanogr. 2008, 53, 1083–1092. [Google Scholar] [CrossRef]
- Riessen, H.P.; Trevett-Smith, J.B. Turning inducible defenses on and off: Adaptive responses of Daphnia to a gape-limited predator. Ecology 2009, 90, 3455–3469. [Google Scholar] [CrossRef]
- Herzog, Q.; Laforsch, C. Modality matters for the expression of inducible defenses: Introducing a concept of predator modality. BMC Biol. 2013, 11, 113. [Google Scholar] [CrossRef] [Green Version]
- Kruppert, S.; Horstmann, M.; Weiss, L.C.; Schaber, C.F.; Gorb, S.N.; Tollrian, R. Push or Pull? The light-weight architecture of the Daphnia pulex carapace is adapted to withstand tension, not compression. J. Morphol. 2016, 277, 1320–1328. [Google Scholar] [CrossRef]
- Kruppert, S.; Horstmann, M.; Weiss, L.C.; Witzel, U.; Schaber, C.F.; Gorb, S.N.; Tollrian, R. Biomechanical properties of predator-induced body armour in the freshwater crustacean Daphnia. Sci. Rep. 2017, 7, 9750. [Google Scholar] [CrossRef] [Green Version]
- Kruppert, S.; Deussen, L.; Weiss, L.C.; Horstmann, M.; Wolff, J.O.; Kleinteich, T.; Gorb, S.N.; Tollrian, R. Zooplankters’ nightmare: The fast and efficient catching basket of larval phantom midges (Diptera: Chaoborus). PLoS ONE 2019, 14, e0214013. [Google Scholar] [CrossRef]
- Colbourne, J.K.; Pfrender, M.E.; Gilbert, D.; Thomas, W.K.; Tucker, A.; Oakley, T.H.; Tokishita, S.; Aerts, A.; Arnold, G.J.; Basu, M.K.; et al. The ecoresponsive genome of Daphnia pulex. Science 2011, 331, 555–561. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.; Zhang, H.; Sha, Y.; Hegg, A.; Ugge, G.E.; Vinterstare, J.; Škerlep, M.; Pärssinen, V.; Herzog, S.D.; Björnerås, C.; et al. Low-latitude zooplankton pigmentation plasticity in response to multiple threats. R. Soc. Open Sci. 2019, 6. [Google Scholar] [CrossRef] [Green Version]
- Lampert, W. Daphnia: Model herbivore, predator and prey. Polish J. Ecol. 2006, 54, 607–620. [Google Scholar]
- Ebert, D. A genome for the environment. Science 2011, 331, 539–540. [Google Scholar] [CrossRef]
- Seda, J.; Petrusek, A. Daphnia as a model organism in limnology and aquatic biology: Introductory remarks. J. Limnol. 2011, 70, 337–344. [Google Scholar] [CrossRef] [Green Version]
- Stollewerk, A. The water flea Daphnia-a “new” model system for ecology and evolution? J. Biol. 2010, 9, 21. [Google Scholar] [CrossRef] [Green Version]
- Weiss, L.C. Sensory ecology of predator-induced phenotypic plasticity. Front. Behav. Neurosci. 2019, 12, 330. [Google Scholar] [CrossRef]
- Christjani, M.; Fink, P.; Von Elert, E. Phenotypic plasticity in three Daphnia genotypes in response to predator kairomone: Evidence for an involvement of chitin deacetylases. J. Exp. Biol. 2016, 219, 1697–1704. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, J.J. Spine development in Brachionus quadridentatus from an Australian billabong: Genetic variation and induction by Asplanchna. In Proceedings of the Hydrobiologia; Springer: Dordrecht, The Netherlands, 2001; Volume 446–447, pp. 19–28. [Google Scholar]
- Juracka, P.J.; Korínek, V.; Petrusek, A. A new Central European species of the Daphnia curvirostris complex, Daphnia hrbaceki sp. nov. (Cladocera, Anomopoda, Daphniidae). Zootaxa 2010, 2718, 1–22. [Google Scholar] [CrossRef]
- Boeing, W.J.; Ramcharan, C.W.; Riessen, H.P. Multiple predator defence strategies in Daphnia pulex and their relation to native habitat. J. Plankton Res. 2006, 28, 571–584. [Google Scholar] [CrossRef] [Green Version]
- Riessen, H.P.; Young, J.D. Daphnia defense strategies in fishless lakes and ponds: One size does not fit all. J. Plankton Res. 2005, 27, 531–544. [Google Scholar] [CrossRef]
- Boeing, W.J.; Ramcharan, C.W.; Riessen, H.P. Clonal variation in depth distribution of Daphnia pulex in response to predator kairomones. Arch. Hydrobiol. 2006, 166, 241–260. [Google Scholar] [CrossRef]
- Tolardo, M.; Da Silva Ferrão-Filho, A.; Santangelo, J.M. Species and clone-dependent effects of tilapia fish (Cichlidae) on the morphology and life-history of temperate and tropical Daphnia. Ecol. Res. 2016, 31, 333–342. [Google Scholar] [CrossRef]
- Stibor, H.; Lampert, W. Components of additive variance in life-history traits of Daphnia hyalina: Seasonal differences in the response to predator signals. Oikos 2000, 88, 129–138. [Google Scholar] [CrossRef]
- Tollrian, R. Predator-induced morphological defenses: Costs, life history shifts, and maternal effects in Daphnia pulex. Ecology 1995, 76, 1691–1705. [Google Scholar] [CrossRef]
- Winder, M.; Buergi, H.R.; Spaak, P. Seasonal vertical distribution of phytoplankton and copepod species in a high-mountain lake. Arch. Hydrobiol. 2003, 158, 197–213. [Google Scholar] [CrossRef]
- Beklioglu, M.; Gozen, A.G.; Yıldırım, F.; Zorlu, P.; Onde, S. Impact of food concentration on diel vertical migration behaviour of Daphnia pulex under fish predation risk. In Proceedings of the Hydrobiologia; Springer: Dordrecht, The Netherlands, 2008; Volume 614, pp. 321–327. [Google Scholar]
- Dawidowicz, P.; Pijanowska, J.; Ciechomski, K. Vertical migration of Chaoborus larvae is induced by the presence of fish. Limnol. Oceanogr. 1990, 35, 1631–1637. [Google Scholar] [CrossRef]
- Perticarrari, A.; Arcifa, M.S.; Rodrigues, R.A. Diel vertical migration of copepods in a Brazilian lake: A mechanism for decreasing risk of Chaoborus predation? Braz. J. Biol. 2004, 64, 289–298. [Google Scholar] [CrossRef]
- Bayly, I.A.E. Aspects of Diel Vertical Migration in Zooplankton, and Its Enigma Variations. In Limnology in Australia; Springer: Dordrecht, The Netherlands, 1986; pp. 349–368. [Google Scholar]
- Cuvier, G. The Animal Kingdom, Arranged After Its Organization; Forming a natural history of animals, and an introduction to comparative anatomy by the late Baron Cuvier; Orr and Co: London, UK, 1854; pp. 1–472. [Google Scholar]
- Wojtal-Frankiewicz, A.; Frankiewicz, P.; Jurczak, T.; Grennan, J.; McCarthy, T.K. Comparison of fish and phantom midge influence on cladocerans diel vertical migration in a dual basin lake. Aquat. Ecol. 2010, 44, 243–254. [Google Scholar] [CrossRef]
- Pangle, K.L.; Peacor, S.D. Non-lethal effect of the invasive predator Bythotrephes longimanus on Daphnia mendotae. Freshw. Biol. 2006, 51, 1070–1078. [Google Scholar] [CrossRef] [Green Version]
- Oram, E.; Spitze, K. Depth selection by Daphnia pulex in response to Chaoborus kairomone. Freshw. Biol. 2013, 58, 409–415. [Google Scholar] [CrossRef]
- Nihongi, A.; Ziarek, J.J.; Uttieri, M.; Sandulli, R.; Zambianchi, E.; Strickler, J.R. Behavioural interseasonal adaptations in Daphnia pulicaria (Crustacea: Cladocera) as induced by predation infochemicals. Aquat. Ecol. 2016, 50, 667–684. [Google Scholar] [CrossRef]
- Gutierrez, M.F.; Gagneten, A.M.; Paggi, J.C. Behavioural responses of two cladocerans and two copepods exposed to fish kairomones. Mar. Freshw. Behav. Physiol. 2011, 44, 289–303. [Google Scholar] [CrossRef]
- Van Gool, E.; Ringelberg, J. What goes down must come up: Symmetry in light-induced migration behaviour of Daphnia. In Proceedings of the Hydrobiologia; Springer: Dordrecht, The Netherlands, 2003; Volume 491, pp. 301–307. [Google Scholar]
- Winder, M.; Spaak, P.; Mooij, W.M. Trade-offs in Daphnia habitat selection. Ecology 2004, 85, 2027–2036. [Google Scholar] [CrossRef]
- Rose, K.C.; Williamson, C.E.; Fischer, J.M.; Connelly, S.J.; Olson, M.; Tucker, A.J.; Noe, D.A. The role of ultraviolet radiation and fish in regulating the vertical distribution of Daphnia. Limnol. Oceanogr. 2012, 57, 1867–1876. [Google Scholar] [CrossRef] [Green Version]
- Huebner, J.D.; Young, D.L.W.; Loadman, N.L.; Lentz, V.J.; Wiegand, M.D. Age-Dependent Survival, Reproduction and Photorepair Activity in Daphnia magna (Straus, 1820) After Exposure to Artificial Ultraviolet Radiation. Photochem. Photobiol. 2006, 82, 1656. [Google Scholar] [CrossRef]
- Ramos-Jiliberto, R.; Dauelsberg, P.; Zúñiga, L.R. Differential tolerance to ultraviolet-B light and photoenzymatic repair in cladocerans from a Chilean lake. Mar. Freshw. Res. 2004, 55, 193–200. [Google Scholar] [CrossRef]
- Leech, D.M.; Williamson, C.E. Is tolerance to UV radiation in zooplankton related to body size, taxon, or lake transparency? Ecol. Appl. 2000, 10, 1530–1540. [Google Scholar] [CrossRef]
- Hylander, S.; Larsson, N.; Hansson, L.A. Zooplankton vertical migration and plasticity of pigmentation arising from simultaneous UV and predation threats. Limnol. Oceanogr. 2009, 54, 483–491. [Google Scholar] [CrossRef]
- Tollrian, R.; Heibl, C. Phenotypic plasticity in pigmentation in Daphnia induced by UV radiation and fish kairomones. Funct. Ecol. 2004, 18, 497–502. [Google Scholar] [CrossRef]
- Loose, C.J.; Dawidowicz, P. Trade-offs in diel vertical migration by zooplankton: The costs of predator avoidance. Ecology 1994, 75, 2255–2263. [Google Scholar] [CrossRef]
- Park, S.; Brett, M.T.; Müller-Navarra, D.C.; Goldman, C.R. Essential fatty acid content and the phosphorus to carbon ratio in cultured algae as indicators of food quality for Daphnia. Freshw. Biol. 2002, 47, 1377–1390. [Google Scholar] [CrossRef] [Green Version]
- Mandal, S.; Abbott Wilkins, R.; Shurin, J.B. Compensatory grazing by Daphnia generates a trade-off between top-down and bottom-up effects across phytoplankton taxa. Ecosphere 2018, 9, e02537. [Google Scholar] [CrossRef] [Green Version]
- Freese, H.M.; Martin-Creuzburg, D. Food quality of mixed bacteria-algae diets for Daphnia magna. Hydrobiologia 2013, 715, 63–76. [Google Scholar] [CrossRef] [Green Version]
- Hartwich, M.; Martin-Creuzburg, D.; Rothhaupt, K.O.; Wacker, A. Oligotrophication of a large, deep lake alters food quantity and quality constraints at the primary producer-consumer interface. Oikos 2012, 121, 1702–1712. [Google Scholar] [CrossRef] [Green Version]
- Brzeziński, T.; Von Elert, E. Predator evasion in zooplankton is suppressed by polyunsaturated fatty acid limitation. Oecologia 2015, 179, 687–697. [Google Scholar] [CrossRef]
- Isanta Navarro, J.; Fromherz, M.; Dietz, M.; Zeis, B.; Schwarzenberger, A.; Martin-Creuzburg, D. Dietary polyunsaturated fatty acid supply improves Daphnia performance at fluctuating temperatures, simulating diel vertical migration. Freshw. Biol. 2019, 64, 1859–1866. [Google Scholar] [CrossRef]
- Wacker, A.; Von Elert, E. Polyunsaturated fatty acids: Evidence for non-substitutable biochemical resources in Daphnia galeata. Ecology 2001, 82, 2507–2520. [Google Scholar] [CrossRef]
- Martin-Creuzburg, D.; Von Elert, E. Good food versus bad food: The role of sterols and polyunsaturated fatty acids in determining growth and reproduction of Daphnia magna. Aquat. Ecol. 2009, 43, 943–950. [Google Scholar] [CrossRef] [Green Version]
- Von Elert, E. Determination of limiting polyunsaturated fatty acids in Daphnia galeata using a new method to enrich food algae with single fatty acids. Limnol. Oceanogr. 2002, 47, 1764–1773. [Google Scholar] [CrossRef]
- Hazel, J.R. Thermal Adaptation in Biological Membranes: Is Homeoviscous Adaptation the Explanation? Annu. Rev. Physiol. 1995, 57, 19–42. [Google Scholar] [CrossRef]
- Martin-Creuzburg, D.; Wacker, A.; Ziese, C.; Kainz, M.J. Dietary lipid quality affects temperature-mediated reaction norms of a freshwater key herbivore. Oecologia 2012, 168, 901–912. [Google Scholar] [CrossRef] [Green Version]
- Gliwicz, Z.M.; Dawidowicz, P.; Maszczyk, P. Low-density anti-predation refuge in Daphnia and Chaoborus? Arch. Hydrobiol. 2006, 167, 101–114. [Google Scholar] [CrossRef]
- Adamczuk, M. Predation follows competition in depth selection behaviour of Cladocera in a deep lake (E Poland). Biol. Lett. 2009, 46, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Dawidowicz, P.; Wielanier, M. Costs of predator avoidance reduce competitive ability of Daphnia. In Proceedings of the Hydrobiologia; Springer: Dordrecht, The Netherlands, 2004; Volume 526, pp. 165–169. [Google Scholar]
- Gélinas, M.; Pinel-Alloul, B.; Ślusarczyk, M. Alternative antipredator responses of two coexisting Daphnia species to negative size selection by YOY perch. J. Plankton Res. 2007, 29, 775–789. [Google Scholar] [CrossRef]
- Hansson, L.A.; Hylander, S. Size-structured risk assessments govern Daphnia migration. Proc. R. Soc. B Biol. Sci. 2009, 276, 331–336. [Google Scholar] [CrossRef] [Green Version]
- Young, J.D.; Riessen, H.P. The interaction of Chaoborus size and vertical distribution determines predation effects on Daphnia. Freshw. Biol. 2005, 50, 993–1006. [Google Scholar] [CrossRef]
- Wood, T.C.; Moore, P.A. Big and bad: How relative predator size and dietary information influence rusty crayfish (faxonius rusticus) behavior and resource-use decisions. Can. J. Zool. 2020, 98, 62–72. [Google Scholar] [CrossRef]
- Watt, P.J.; Young, S. Effect of predator chemical cues on daphnia behaviour in both horizontal and vertical planes. Anim. Behav. 1994, 48, 861–869. [Google Scholar] [CrossRef] [Green Version]
- Taleb, H.; Lair, N.; Reyes Marchant, P.; Jamet, J.L. Observations on vertical migrations of zooplankton at four different stations of a small, eutrophic, temperate zone lake, in relation to their predators. In Ergebnisse der Limnologie ERLIA 6-Archiv für Hydrobiologie; Schweizerbart Science Publishers: Stuttgart, Germany, 1993; pp. 199–216. [Google Scholar]
- Van De Meutter, F.; Stoks, R.; De Meester, L. Behavioral linkage of pelagic prey and littoral predators: Microhabitat selection by Daphnia induced by damselfly larvae. Oikos 2004, 107, 265–272. [Google Scholar] [CrossRef]
- Lindén, E.; Lehtiniemi, M.; Viitasalo, M. Predator avoidance behaviour of Baltic littoral mysids Neomysis integer and Praunus flexuosus. Mar. Biol. 2003, 143, 845–850. [Google Scholar] [CrossRef]
- Nihan Tavşanoǧlu, Ü.; Idil Çakiroǧlu, A.; Erdoǧan, Ş.; Meerhoff, M.; Jeppesen, E.; Beklioglu, M. Sediments, not plants, offer the preferred refuge for Daphnia against fish predation in Mediterranean shallow lakes: An experimental demonstration. Freshw. Biol. 2012, 57, 795–802. [Google Scholar] [CrossRef]
- Gutierrez, M.F.; Molina, F.R.; Carvalho, D.A. Behavioural responses of freshwater zooplankton vary according to the different alarm signals of their invertebrate predators. Mar. Freshw. Behav. Physiol. 2013, 45, 317–331. [Google Scholar] [CrossRef]
- Gilbert, J.J. Morphological and behavioral responses of a rotifer to the predator Asplanchna. J. Plankton Res. 2014, 36, 1576–1584. [Google Scholar] [CrossRef]
- De Meester, L.; Pijanowska, J. On the trait-specificity of the response of Daphnia genotypes to the chemical presence of a predator. In Zooplankton: Sensory Ecology and Physiology; CRC Press: Boca Raton, FL, USA, 1996; pp. 407–417. ISBN 90-5699-022-5. [Google Scholar]
- Brewer, M. Interactive effects of fish kairomone and light on Daphnia escape behavior. J. Plankton Res. 1999, 21, 1317–1335. [Google Scholar] [CrossRef]
- Chang, K.H.; Hanazato, T. Vulnerability of cladoceran species to predation by the copepod Mesocyclops leuckarti: Laboratory observations on the behavioural interactions between predator and prey. Freshw. Biol. 2003, 48, 476–484. [Google Scholar] [CrossRef]
- Pijanowska, J.; Dawidowicz, P.; Weider, L.J. Predator-induced escape response in Daphnia. Arch. Hydrobiol. 2006, 167, 77–87. [Google Scholar] [CrossRef]
- Szulkin, M.; Dawidowicz, P.; Dodson, S.I. Behavioural uniformity as a response to cues of predation risk. Anim. Behav. 2006, 71, 1013–1019. [Google Scholar] [CrossRef]
- Rose, R.M.; Warne, M.S.J.; Lim, R.P. Exposure to chemicals exuded by fish reduces the filtration and ingestion rates of Ceriodaphnia cf. dubia. Hydrobiologia 2003, 501, 215–217. [Google Scholar] [CrossRef]
- Beckerman, A.P.; Wieski, K.; Baird, D.J. Behavioural versus physiological mediation of life history under predation risk. Oecologia 2007, 152, 335–343. [Google Scholar] [CrossRef]
- Kerfoot, W.C. Combat between predatory copepods and their prey: Cyclops, Epischura, and Bosmina. Limnol. Oceanogr. 1978, 23, 1089–1102. [Google Scholar] [CrossRef]
- La, G.H.; Choi, J.Y.; Chang, K.H.; Jang, M.H.; Joo, G.J.; Kim, H.W. Mating behavior of Daphnia: Impacts of predation risk, food quantity, and reproductive phase of females. PLoS ONE 2014, 9, e104545. [Google Scholar] [CrossRef] [Green Version]
- Sarma, S.S.S.; Dumont, H.J.; Nandini, S. Interactions between the Anomopod Cladocerans Ceriodaphnia dubia, C. cornuta, Simocephalus vetulus and S. serrulatus, the aphanoneurid worm Aeolosoma sp., and the fish Skiffia lermae: Predation or competition, or both? In Proceedings of the Hydrobiologia; Springer: Berlin/Heidelberg, Germany, 2004; Volume 526, pp. 147–156. [Google Scholar]
- Greene, C.H. Selective Predation in Freshwater Zooplankton Communities. Int. Rev. Hydrobiol. Hydrogr. 1983, 68, 297–315. [Google Scholar] [CrossRef]
- Dodson, S.I. Adaptive change in plankton morphology in response to size-selective predation: A new hypothesis of cyclomorphosis. Limnol. Oceanogr. 1974, 19, 721–729. [Google Scholar] [CrossRef]
- Dodson, S.I. Cyclomorphosis in Daphnia galeata mendotae Birge and D. retrocurva Forbes as a predator-induced response. Freshw. Biol. 1988, 19, 109–114. [Google Scholar] [CrossRef]
- Chang, K.H.; Hanazato, T. Seasonal and reciprocal succession and cyclomorphosis of two Bosmina species (Cladocera, Crustacea) co-existing in a lake: Their relationship with invertebrate predators. J. Plankton Res. 2003, 25, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Juračka, P.J.; Laforsch, C.; Petrusek, A. Neckteeth formation in two species of the Daphnia curvirostris complex (Crustacea: Cladocera). J. Limnol. 2011, 70, 359–368. [Google Scholar] [CrossRef]
- Rabus, M.; Waterkeyn, A.; Van Pottelbergh, N.; Brendonck, L.; Laforsch, C. Interclonal variation, effectiveness and long-term implications of Triops-induced morphological defences in Daphnia magna Strauss. J. Plankton Res. 2012, 34, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Miyakawa, H.; Sugimoto, N.; Kohyama, T.I.; Iguchi, T.; Miura, T. Intra-specific variations in reaction norms of predator-induced polyphenism in the water flea Daphnia pulex. Ecol. Res. 2015, 30, 705–713. [Google Scholar] [CrossRef]
- O’Brien, W.J.; Kettle, D.; Riessen, H. Helmets and Invisible Armor: Structures Reducing Predation from Tactile and Visual Planktivores. Ecology 1979, 60, 287–294. [Google Scholar] [CrossRef]
- Grant, J.W.G.; Bayly, I.A.E. Predator induction of crests in morphs of the Daphnia carinata King complex. Limnol. Oceanogr. 1981, 26, 201–218. [Google Scholar] [CrossRef]
- Tollrian, R.; Dodson, S. Inducible defenses in Cladocera: Constraints, costs, and multipredator environments. In The Ecology and Evolution of Inducible Defenses; Princeton University Press: Princeton, NJ, USA, 1999; pp. 177–202. [Google Scholar]
- Dahm, E. Zur Biologie von Notonecta glauca (Insecta, Hemiptera) unter besonderer Berücksichtigung der fischereilichen Schadwirkung. Int. Rev. Hydrobiol. Hydrogr. 1972, 57, 429–461. [Google Scholar] [CrossRef]
- Fryer, G. Studies on the functional morphology and biology of the Notostraca (Crustacea: Branchiopoda). Philos. Trans. R. Soc. London. B 1988, 321, 27–124. [Google Scholar] [CrossRef]
- Havel, J.E.; Dodson, S.I. Reproductive costs of Chaoborus-induced polymorphism in Daphnia pulex. Hydrobiologia 1987, 150, 273–281. [Google Scholar] [CrossRef]
- Dzialowski, A.R.; Lennon, J.T.; O’Brien, W.J.; Smith, V.H. Predator-induced phenotypic plasticity in the exotic cladoceran Daphnia lumholtzi. Freshw. Biol. 2003, 48, 1593–1602. [Google Scholar] [CrossRef] [Green Version]
- Amundsen, P.A.; Siwertsson, A.; Primicerio, R.; Bøhn, T. Long-term responses of zooplankton to invasion by a planktivorous fish in a subarctic watercourse. Freshw. Biol. 2009, 54, 24–34. [Google Scholar] [CrossRef] [Green Version]
- Hart, R.C.; Bychek, E.A. Body size in freshwater planktonic crustaceans: An overview of extrinsic determinants and modifying influences of biotic interactions. Hydrobiologia 2011, 668, 61–108. [Google Scholar] [CrossRef]
- Effertz, C.; Von Elert, E. Light intensity controls anti-predator defences in Daphnia: The suppression of life-history changes. Proc. R. Soc. B Biol. Sci. 2014, 281, 20133250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Effertz, C.; Von Elert, E. Coupling of anti-predator defences in Daphnia: The importance of light. Hydrobiologia 2017, 798, 5–13. [Google Scholar] [CrossRef]
- La, G.H.; Chang, K.H.; Jang, M.H.; Joo, G.J.; Kim, H.W. Comparison of morphological defences in asexually and sexually reproduced eggs of Daphnia (D. galeata and D. similis) against fish kairomones. Russ. J. Ecol. 2014, 45, 314–318. [Google Scholar] [CrossRef]
- Carter, M.J.; Vega-Retter, C.; Ramos-Jiliberto, R. Non-lethal effects of invertebrate predators on Daphnia: Morphological and life-history consequences of water mite kairomone. Freshw. Biol. 2008, 53, 1857–1867. [Google Scholar] [CrossRef]
- Ma, X.; Wolinska, J.; Petrusek, A.; Gießler, S.; Hu, W.; Yin, M. The phenotypic plasticity in Chinese populations of Daphnia similoides sinensis: Recurvate helmeted forms are associated with the presence of predators. J. Plankton Res. 2016, 38, 855–864. [Google Scholar] [CrossRef] [Green Version]
- Stoks, R.; Govaert, L.; Pauwels, K.; Jansen, B.; De Meester, L. Resurrecting complexity: The interplay of plasticity and rapid evolution in the multiple trait response to strong changes in predation pressure in the water flea Daphnia magna. Ecol. Lett. 2016, 19, 180–190. [Google Scholar] [CrossRef]
- West-Eberhard, M.J. Phenotypic accommodation: Adaptive innovation due to developmental plasticity. In Proceedings of the Journal of Experimental Zoology Part B: Molecular and Developmental Evolution; Wiley: Hoboken, NJ, USA, 2005; Volume 304, pp. 610–618. [Google Scholar]
- West-Eberhard, M.J. Developmental Plasticity and Evolution; Oxford University Press: Oxford, UK, 2003; Volume 1, pp. 1–816. [Google Scholar]
- Harvell, C.D. The ecology and evolution of inducible defenses. Q. Rev. Biol. 1990, 65, 323–340. [Google Scholar] [CrossRef]
- Tollrian, R. Fish-kairomone induced morphological changes in Daphnia lumholtzi (Sars). Arch. Hydrobiol. 1994, 130, 69–75. [Google Scholar]
- Swaffar, S.M.; O’Brien, W.J. Spines of Daphnia lumholtzi create feeding difficulties for juvenile bluegill sunfish (Lepomis macrochirus). J. Plankton Res. 1996, 18, 1055–1061. [Google Scholar] [CrossRef] [Green Version]
- Lindholm, M. Predator-induced cyclomorphosis of Daphnia laevis (Branchiopoda, Cladocera) in a tropical floodplain (Okavango Delta, Botswana). Crustaceana 2002, 75, 803–814. [Google Scholar] [CrossRef]
- Engel, K.; Schreder, T.; Tollrian, R. Morphological defences of invasive Daphnia lumholtzi protect against vertebrate and invertebrate predators. J. Plankton Res. 2014, 36, 1140–1145. [Google Scholar] [CrossRef] [Green Version]
- Engel, K.; Tollrian, R. Inducible defences as key adaptations for the successful invasion of Daphnia lumholtzi in North America? Proc. R. Soc. B Biol. Sci. 2009, 276, 1865–1873. [Google Scholar] [CrossRef] [Green Version]
- Zaret, T.M. Predators, invisible prey, and the nature of polymorphism in the Cladocera (class Crustacea). Limnol. Oceanogr. 1972, 17, 171–184. [Google Scholar] [CrossRef]
- Brönmark, C.; Hansson, L.A. The Biology of Lakes and Ponds; Oxford University Press: Oxford, UK, 2017; Volume 3, pp. 1–337. ISBN 9780198713593. [Google Scholar]
- Black, A.R.; Dodson, S.I. Demographic costs of Chaoborus-induced phenotypic plasticity in Daphnia pulex. Oecologia 1990, 83, 117–122. [Google Scholar] [CrossRef]
- Repka, S.; Walls, M.; Ketola, M. Neck spine protects Daphnia pulex from predation by Chaoborus, but individuals with longer tail spine are at a greater risk. J. Plankton Res. 1995, 17, 393–403. [Google Scholar] [CrossRef]
- Brooks, J.L. The systematics of North American Daphnia.; Memoirs of the Connecticut Academy of Arts and Sciences-Yale University Press: New Haven, CT, USA, 1957; Volume 13, pp. 1–180. [Google Scholar]
- Krueger, D.A.; Dodson, S.I. Embryological induction and predation ecology in Daphnia pulex. Limnol. Oceanogr. 1981, 26, 219–223. [Google Scholar] [CrossRef]
- Havel, J.E.; Dodson, S.I. Chaoborus predation on typical and spined morphs of Daphnia pulex: Behavioral observations. Limnol. Oceanogr. 1984, 29, 487–494. [Google Scholar] [CrossRef]
- Tollrian, R. Neckteeth formation in Daphnia pulex as an example of continuous phenotypic plasticity: Morphological effects of Chaoborus kairomone concentration and their quantification. J. Plankton Res. 1993, 15, 1309–1318. [Google Scholar] [CrossRef]
- Sell, A.F. Morphological defenses induced in situ by the invertebrate predator Chaoborus: Comparison of responses between Daphnia pulex and D. rosea. Oecologia 2000, 125, 150–160. [Google Scholar] [CrossRef]
- Tollrian, R. Chaoborus crystallinus predation on Daphnia pulex: Can induced morphological changes balance effects of body size on vulnerability? Oecologia 1995, 101, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Laforsch, C.; Tollrian, R. Embryological aspects of inducible morphological defenses in Daphnia. J. Morphol. 2004, 262, 701–707. [Google Scholar] [CrossRef] [PubMed]
- Maurone, C.; Suppa, A.; Rossi, V. Polymorphisms in predator induced defences of coexisting Daphnia pulex and D. longispina. Hydrobiologia 2018, 823, 121–133. [Google Scholar] [CrossRef]
- Sperfeld, E.; Nilssen, J.P.; Rinehart, S.; Schwenk, K.; Hessen, D.O. Ecology of predator-induced morphological defense traits in Daphnia longispina (Cladocera, Arthropoda). Oecologia 2020, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Benzie, J.A.H. Cladocera: The Genus Daphnia (including Daphniopsis) (Anomopoda: Daphniidae). Guides to the Identification of the Microinvertebrates of the Continental Waters of the World, Volume 21. Q. Rev. Biol. 2005, 80, 491. [Google Scholar] [CrossRef]
- Kotov, A.A.; Ishida, S.; Taylor, D.J. A new species in the Daphnia curvirostris (Crustacea: Cladocera) complex from the eastern Palearctic with molecular phylogenetic evidence for the independent origin of neckteeth. J. Plankton Res. 2006, 28, 1067–1079. [Google Scholar] [CrossRef]
- Imai, M.; Naraki, Y.; Tochinai, S.; Miura, T. Elaborate regulations of the predator-induced polyphenism in the water flea Daphnia pulex: Kairomone-Sensitive periods and life-history tradeoffs. J. Exp. Zool. Part A Ecol. Genet. Physiol. 2009, 311, 788–795. [Google Scholar] [CrossRef]
- Pastorok, R.A. Prey Vulnerability and Size Selection by Chaoborus Larvae. Ecology 1981, 62, 1311–1324. [Google Scholar] [CrossRef]
- Laforsch, C.; Tollrian, R. Extreme helmet formation in Daphnia cucullata induced by small-scale turbulence. J. Plankton Res. 2004, 26, 81–87. [Google Scholar] [CrossRef]
- Laforsch, C.; Ngwa, W.; Grill, W.; Tollrian, R. An acoustic microscopy technique reveals hidden morphological defenses in Daphnia. Proc. Natl. Acad. Sci. USA 2004, 101, 15911–15914. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, W.J.; Vinyard, G.L. Polymorphism and predation: The effect of invertebrate predation on the distribution of two varieties of Daphnia carinata in South India ponds. Limnol. Oceanogr. 1978, 23, 452–460. [Google Scholar] [CrossRef]
- Barry, M.J.; Bayly, I.A.E. Further studies on predator induction of crests in australian daphnia and the effects of crests on predation. Mar. Freshw. Res. 1985, 36, 519–535. [Google Scholar] [CrossRef]
- Laforsch, C.; Haas, A.; Jung, N.; Schwenk, K.; Tollrian, R.; Petrusek, A. “Crown of thorns”of Daphnia: An exceptional inducible defense discovered by DNA barcoding. Commun. Integr. Biol. 2009, 2, 379–381. [Google Scholar] [CrossRef] [PubMed]
- Petrusek, A.; Tollrian, R.; Schwenk, K.; Haas, A.; Laforsch, C. A “crown of thorns” is an inducible defense that protects Daphnia against an ancient predator. Proc. Natl. Acad. Sci. 2009, 106, 2248–2252. [Google Scholar] [CrossRef] [Green Version]
- Rabus, M.; Laforsch, C. Growing large and bulky in the presence of the enemy: Daphnia magna gradually switches the mode of inducible morphological defences. Funct. Ecol. 2011, 25, 1137–1143. [Google Scholar] [CrossRef]
- Herzog, Q.; Rabus, M.; Ribeiro, B.W.; Laforsch, C. Inducible defenses with a “twist”: Daphnia barbata abandons bilateral symmetry in response to an ancient predator. PLoS ONE 2016, 11, e0148556. [Google Scholar] [CrossRef] [Green Version]
- Rabus, M.; Söllradl, T.; Clausen-Schaumann, H.; Laforsch, C. Uncovering Ultrastructural Defences in Daphnia magna-An Interdisciplinary Approach to Assess the Predator-Induced Fortification of the Carapace. PLoS ONE 2013, 8, e67856. [Google Scholar] [CrossRef] [Green Version]
- Møller, O.S.; Olesen, J.; Høeg, J.T. SEM studies on the early larval development of Triops cancriformis (Bosc) (Crustacea: Branchiopoda, Notostraca). Acta Zool. 2003, 84, 267–284. [Google Scholar] [CrossRef]
- Laforsch, C.; Tollrian, R. A new preparation technique of daphnids for Scanning Electron Microscopy using hexamethyldisilazane. Arch. Hydrobiol. 2000, 149, 587–596. [Google Scholar] [CrossRef]
- Bungartz, B.; Branstrator, D.K. Morphological changes in Daphnia mendotae in the chemical presence of Bythotrephes longimanus. Arch. Hydrobiol. 2003, 158, 97–108. [Google Scholar] [CrossRef]
- Branstrator, D.K. Ecological Interactions Between Bythotrephes cederstroemi and Leptodora kindtii and the Implications for Species Replacement in Lake Michigan. J. Great Lakes Res. 1995, 21, 670–679. [Google Scholar] [CrossRef]
- Hellsten, M.; Lagergren, R.; Stenson, J. Can extreme morphology in Bosmina reduce predation risk from Leptodora? An experimental test. Oecologia 1999, 118, 23–28. [Google Scholar] [CrossRef]
- Chang, K.H.; Hanazato, T. Predation impact of Leptodora kindtii on population dynamics and morphology of Bosmina fatalis and B. longirostris in mesocosms. Freshw. Biol. 2004, 49, 253–264. [Google Scholar] [CrossRef]
- Lagergren, R. Chemical cues from the invertebrate predator Leptodora kindtii affect the development of cyclomorphic traits in Eubosmina coregoni gibbera. J. Plankton Res. 2000, 22, 1213–1219. [Google Scholar] [CrossRef] [Green Version]
- Chang, K.-H.; Hanazato, T. Morphological defense of Bosmina fatalis against invertebrate predators in Lake Suwa. Int. Ver. Theor. Angew. Limnol 2002, 28, 1279–1283. [Google Scholar] [CrossRef]
- Herzig, A.; Auer, B. The feeding behaviour of Leptodora kindti and its impact on the zooplankton community of Neusiedler See (Austria). Hydrobiologia 1990, 198, 107–117. [Google Scholar] [CrossRef]
- Jankowski, T. Predation of freshwater jellyfish on Bosmina: The consequences for population dynamics, body size, and morphology. In Proceedings of the Hydrobiologia; Springer: Berlin/Heidelberg, Germany, 2004; Volume 530–531, pp. 521–528. [Google Scholar]
- Kerfoot, W.C. Baltic Eubosmina morphological radiation: Sensitivity to invertebrate predators (induction) and observations on genetic differences. Arch. Hydrobiol. 2006, 167, 147–168. [Google Scholar] [CrossRef]
- Sakamoto, M.; Hanazato, T. Antennule shape and body size of Bosmina: Key factors determining its vulnerability to predacious copepoda. Limnology 2008, 9, 27–34. [Google Scholar] [CrossRef]
- Kerfoot, W.C. Implications of copepod predation. Limnol. Oceanogr. 1977, 22, 316–325. [Google Scholar] [CrossRef]
- Johnsen, G.H.; Raddum, G.G. A morphological study of two populations of Bosmina longispina exposed to different predation. J. Plankton Res. 1987, 9, 297–304. [Google Scholar] [CrossRef]
- DeBeauchamp, P. Un facteur de la variabilite chez les Rotiferes du genre Brachionus. Compt. Rend. 1952, 234, 573–575. [Google Scholar]
- Gilbert, J.J. Rotifer ecology and embryological induction. Science 1966, 151, 1234–1237. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, J.J.; Stemberger, R.S. Asplanchna-induced polymorphism in the rotifer Keratella slacki. Limnol. Oceanogr. 1984, 29, 1309–1316. [Google Scholar] [CrossRef]
- Gilbert, J.J. Predator-specific inducible defenses in the rotifer Keratella tropica. Freshw. Biol. 2009, 54, 1933–1946. [Google Scholar] [CrossRef]
- Nandini, S.; Perez-Chavez, R.; Sarma, S.S.S. The effect of prey morphology on the feeding behaviour and population growth of the predatory rotifer Asplanchna sieboldi: A case study using five species of Brachionus (Rotifera). Freshw. Biol. 2003, 48, 2131–2140. [Google Scholar] [CrossRef]
- Sarma, S.S.S.; Resendiz, R.A.L.; Nandini, S. Morphometric and demographic responses of brachionid prey (Brachionus calyciflorus Pallas and Plationus macracanthus (Daday)) in the presence of different densities of the predator Asplanchna brightwellii (Rotifera: Asplanchnidae). Hydrobiologia 2011, 662, 179–187. [Google Scholar] [CrossRef]
- Gilbert, J.J. Induction of different defences by two enemies in the rotifer Keratella tropica: Response priority and sensitivity to enemy density. Freshw. Biol. 2011, 56, 926–938. [Google Scholar] [CrossRef]
- Gilbert, J.J. Temperature, kairomones, and phenotypic plasticity in the rotifer Keratella tropica (Apstein, 1907). Hydrobiologia 2011, 678, 179–190. [Google Scholar] [CrossRef]
- Bernot, R.J.; Dodds, W.K.; Quist, M.C.; Guy, C.S. Temperature and kairomone induced life history plasticity in coexisting Daphnia. Aquat. Ecol. 2006, 40, 361–372. [Google Scholar] [CrossRef]
- Boeing, W.J.; Wissel, B.; Ramcharan, C.W. Costs and benefits of Daphnia defense against Chaoborus in nature. Can. J. Fish. Aquat. Sci. 2005, 62, 1286–1294. [Google Scholar] [CrossRef] [Green Version]
- Chakri, K.; Touati, L.; Alfarhan, A.H.; Al-Rasheid, K.A.S.; Samraoui, B. Effect of vertebrate and invertebrate kairomones on the life history of Daphnia magna Straus (Crustacea: Branchiopoda). C. R. Biol. 2010, 333, 836–840. [Google Scholar] [CrossRef] [PubMed]
- Santangelo, J.M.; Soares, B.N.; Paes, T.; Maia-Barbosa, P.; Tollrian, R.; Bozelli, R.L. Effects of vertebrate and invertebrate predators on the life history of daphnia similis and moina macrocopa (Crustacea: Cladocera). Ann. Limnol. 2018, 54, 25. [Google Scholar] [CrossRef]
- Taylor, B.E.; Gabriel, W. Optimal Adult Growth of Daphnia in a Seasonal Environment. Funct. Ecol. 1993, 7, 513. [Google Scholar] [CrossRef] [Green Version]
- Taylor, B.E.; Gabriel, W. To Grow or Not to Grow: Optimal Resource Allocation for Daphnia. Am. Nat. 1992, 139, 248–266. [Google Scholar] [CrossRef] [Green Version]
- Santangelo, J.M.; Bozelli, R.L.; De Esteves, F.A.; Tollrian, R. Predation cues do not affect the induction and termination of diapause in small-bodied cladocerans. Freshw. Biol. 2010, 55, 1577–1586. [Google Scholar] [CrossRef]
- Slusarczyk, M.; Rybicka, B. Role of temperature in diapause response to fish kairomones in crustacean Daphnia. J. Insect Physiol. 2011, 57, 676–680. [Google Scholar] [CrossRef]
- Slusarczyk, M.; Dawidowicz, P.; Rygielska, E. Hide, rest or die: A light-mediated diapause response in Daphnia magna to the threat of fish predation. Freshw. Biol. 2005, 50, 141–146. [Google Scholar] [CrossRef]
- Lass, S.; Vos, M.; Wolinska, J.; Spaak, P. Hatching with the enemy: Daphnia diapausing eggs hatch in the presence of fish kairomones. Chemoecology 2005, 15, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Waterkeyn, A.; Van Pottelbergh, N.; Vanoverbeke, J.; Vanschoenwinkel, B.; De Meester, L.; Brendonck, L. Constitutive but no Triops-induced differences in bet-hedging strategies for hatching in Daphnia. Hydrobiologia 2013, 715, 29–35. [Google Scholar] [CrossRef]
- Mikulski, A.; Lipowska, D.; Pijanowska, J. Ontogenetic changes in Daphnia responsiveness to fish kairomone. In Proceedings of the Hydrobiologia; Springer: Dordrecht, The Netherlands, 2004; Volume 526, pp. 219–224. [Google Scholar]
- Mikulski, A.; Pijanowska, J. When and how can Daphnia prepare their offspring for the threat of predation? Hydrobiologia 2010, 643, 21–26. [Google Scholar] [CrossRef]
- Ślusarczyk, M.; Rygielska, E. Fish faeces as the primary source of chemical cues inducing fish avoidance diapause in Daphnia magna. In Proceedings of the Hydrobiologia; Springer: Dordrecht, The Netherlands, 2004; Volume 526, pp. 231–234. [Google Scholar]
- Wolinska, J.; Löffler, A.; Spaak, P. Taxon-specific reaction norms to predator cues in a hybrid Daphnia complex. Freshw. Biol. 2007, 52, 1198–1209. [Google Scholar] [CrossRef]
- Beckerman, A.P.; Rodgers, G.M.; Dennis, S.R. The reaction normof size and age at maturity under multiple predator risk. J. Anim. Ecol. 2010, 79, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- Riessen, H.P. Costs of predator-induced morphological defences in Daphnia. Freshw. Biol. 2012, 57, 1422–1433. [Google Scholar] [CrossRef]
- Klintworth, S.; Von Elert, E. Risk of predation alters resource allocation in Daphnia under food limitation. J. Plankton Res. 2020, 42, 45–56. [Google Scholar] [CrossRef]
- Hülsmann, S.; Vijverberg, J.; Boersma, M.; Mooij, W.M. Effects of infochemicals released by gape-limited fish on life history traits of Daphnia: A maladaptive response? J. Plankton Res. 2004, 26, 535–543. [Google Scholar] [CrossRef]
- Relyea, R.A. How prey respond to combined predators: A review and an empirical test. Ecology 2003, 84, 1827–1839. [Google Scholar] [CrossRef]
- Tanner, C.J.; Branstrator, D.K. Generational and dual-species exposures to invertebrate predators influence relative head size in Daphnia mendotae. J. Plankton Res. 2006, 28, 793–802. [Google Scholar] [CrossRef] [Green Version]
- Laforsch, C.; Tollrian, R. Inducible defenses in multipredator environments: Cyclomorphosis in Daphnia cucullata. Ecology 2004, 85, 2302–2311. [Google Scholar] [CrossRef] [Green Version]
- Wilczynski, W.; Dynak, P.; Babkiewicz, E.; Bernatowicz, P.; Leniowski, K.; Maszczyk, P. The combined effects of hypoxia and fish kairomones on several physiological and life history traits of Daphnia. Freshw. Biol. 2019, 64, 2204–2220. [Google Scholar] [CrossRef]
- Jeyasingh, P.D.; Weider, L.J. Phosphorus availability mediates plasticity in life-history traits and predator-prey interactions in Daphnia. Ecol. Lett. 2005, 8, 1021–1028. [Google Scholar] [CrossRef]
- Maszczyk, P.; Bartosiewicz, M. Threat or treat: The role of fish exudates in the growth and life history of Daphnia. Ecosphere 2012, 3, art91. [Google Scholar] [CrossRef]
- Pauwels, K.; Stoks, R.; De Meester, L. Enhanced anti-predator defence in the presence of food stress in the water flea Daphnia magna. Funct. Ecol. 2010, 24, 322–329. [Google Scholar] [CrossRef]
- Bell, A.T.C.; Murray, D.L.; Prater, C.; Frost, P.C. Fear and food: Effects of predator-derived chemical cues and stoichiometric food quality on Daphnia. Limnol. Oceanogr. 2019, 64, 1706–1715. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Jansen, M.; De Meester, L.; Stoks, R. Energy storage and fecundity explain deviations from ecological stoichiometry predictions under global warming and size-selective predation. J. Anim. Ecol. 2016, 85, 1431–1441. [Google Scholar] [CrossRef] [PubMed]
- Preisser, E.L.; Bolnick, D.I. The many faces of fear: Comparing the pathways and impacts of nonconsumptive predator effects on prey populations. PLoS ONE 2008, 3, e2465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gliwicz, Z.M.; Maszczyk, P. Daphnia growth is hindered by chemical information on predation risk at high but not at low food levels. Oecologia 2007, 150, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Pietrzak, B.; Rabus, M.; Religa, M.; Laforsch, C.; Dańko, M.J. Phenotypic plasticity of senescence in Daphnia under predation impact: No ageing acceleration when the perceived risk decreases with age. R. Soc. Open Sci. 2020, 7, 191382. [Google Scholar] [CrossRef] [Green Version]
- Bertram, C.R.; Pinkowski, M.; Hall, S.R.; Duffy, M.A.; Cáceres, C.E. Trait-mediated indirect effects, predators, and disease: Test of a size-based model. Oecologia 2013, 173, 1023–1032. [Google Scholar] [CrossRef]
- Yin, M.; Laforsch, C.; Lohr, J.N.; Wolinska, J. Predator-induced defense makes daphnia more vulnerable to parasites. Evolution 2011, 65, 1482–1488. [Google Scholar] [CrossRef]
- Hesse, O.; Engelbrecht, W.; Laforsch, C.; Wolinska, J. Fighting parasites and predators: How to deal with multiple threats? BMC Ecol. 2012, 12, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammill, E.; Rogers, A.; Beckerman, A.P. Costs, benefits and the evolution of inducible defences: A case study with Daphnia pulex. J. Evol. Biol. 2008, 21, 705–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pohnert, G.; Steinke, M.; Tollrian, R. Chemical cues, defence metabolites and the shaping of pelagic interspecific interactions. Trends Ecol. Evol. 2007, 22, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Pohnert, G. Finding the fish factor. Elife 2019, 8. [Google Scholar] [CrossRef]
- Akkas, S.B.; Kepenek, A.O.; Beklioglu, M.; Severcan, F. Molecular approach to the chemical characterization of fish-exuded kairomone: A Fourier transform infrared spectroscopic study. Aquat. Sci. 2010, 72, 71–83. [Google Scholar] [CrossRef]
- Beklioglu, M.; Telli, M.; Gozen, A.G. Fish and mucus-dwelling bacteria interact to produce a kairomone that induces diel vertical migration in Daphnia. Freshw. Biol. 2006, 51, 2200–2206. [Google Scholar] [CrossRef]
- Castro, B.B.; Consciência, S.; Gonçalves, F. Life history responses of Daphnia longispina to mosquitofish (Gambusia holbrooki) and pumpkinseed (Lepomis gibbosus) kairomones. In Proceedings of the Hydrobiologia; Springer: Dordrecht, The Netherlands, 2007; Volume 594, pp. 165–174. [Google Scholar]
- Von Elert, E.; Stibor, H. Predator-mediated life history shifts in Daphnia: Enrichment and preliminary chemical characterisation of a kairomone exuded by fish. Arch. Hydrobiol. 2006, 167, 21–35. [Google Scholar] [CrossRef]
- Weber, A. More than one “fish kairomone”? Perch and stickleback kairomones affect Daphnia life history traits differently. Hydrobiologia 2003, 498, 143–150. [Google Scholar] [CrossRef]
- Tollrian, R.; Laforsch, C. Linking predator kairomones and turbulence: Synergistic effects and ultimate reasons for phenotypic plasticity in Daphnia cucullata. Arch. Hydrobiol. 2006, 167, 135–146. [Google Scholar] [CrossRef]
- Heuschele, J.; Selander, E. The chemical ecology of copepods. J. Plankton Res. 2014, 36, 895–913. [Google Scholar] [CrossRef] [Green Version]
- Rittschof, D.; Cohen, J.H. Crustacean peptide and peptide-like pheromones and kairomones. Peptides 2004, 25, 1503–1516. [Google Scholar] [CrossRef] [PubMed]
- Kusch, J.; Heckmann, K. Isolation of the Lembadion-factor, A morphogenetically active signal, that induces Euplotes cells to change from their ovoid form into a larger lateral winged morph. Dev. Genet. 1992, 13, 241–246. [Google Scholar] [CrossRef]
- Hahn, M.A.; Effertz, C.; Bigler, L.; Elert, E. Von 5α-cyprinol sulfate, a bile salt from fish, induces diel vertical migration in Daphnia. Elife 2019, 8, e44791. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, A.F.; Hagey, L.R.; Krasowski, M.D. Bile salts of vertebrates: Structural variation and possible evolutionary signifi cance. J. Lipid Res. 2010, 51, 226–246. [Google Scholar] [CrossRef] [Green Version]
- Yasumoto, K.; Nishigami, A.; Yasumoto, M.; Kasai, F.; Okada, Y.; Kusumi, T.; Ooi, T. Aliphatic sulfates released from Daphnia induce morphological defense of phytoplankton: Isolation and synthesis of kairomones. Tetrahedron Lett. 2005, 46, 4765–4767. [Google Scholar] [CrossRef]
- Parejko, K.; Dodson, S. Progress towards characterization of a predator/prey kairomone: Daphnia pulex and Chaoborus americanus. Hydrobiologia 1990, 198, 51–59. [Google Scholar] [CrossRef]
- Loose, C.J.; Von Elert, E.; Dawidowicz, P. Chemically-Induced Diel Vertical Migration in Daphnia-a New Bioassay for Kairomones Exuded by Fish. Arch. Hydrobiol. 1993, 126, 329–337. [Google Scholar]
- Mikulski, A.; Czernik, M.; Pijanowska, J. Induction time and reversibility of changes in Daphnia life history caused by the presence of fish. J. Plankton Res. 2005, 27, 757–762. [Google Scholar] [CrossRef] [Green Version]
- Tollrian, R.; Duggen, S.; Weiss, L.C.; Laforsch, C.; Kopp, M. Density-dependent adjustment of inducible defenses. Sci. Rep. 2015, 5, 12736. [Google Scholar] [CrossRef] [Green Version]
- Hahn, M.; Von Elert, E.; Bigler, L.; Díaz Hernández, M.D.; Schloerer, N.E. 5α-Cyprinol sulfate: Complete NMR assignment and revision of earlier published data, including the submission of a computer-readable assignment in NMReDATA format. Magn. Reson. Chem. 2018, 56, 1201–1207. [Google Scholar] [CrossRef]
- Ringelberg, J.; Van Gool, E. Do bacteria, not fish, produce “fish kairomone”? J. Plankton Res. 1998, 20, 1847–1852. [Google Scholar] [CrossRef] [Green Version]
- Pestana, J.L.T.; Loureiro, S.; Baird, D.J.; Soares, A.M.V.M. Pesticide exposure and inducible antipredator responses in the zooplankton grazer, Daphnia magna Straus. Chemosphere 2010, 78, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M.D.; Bairos-Novak, K.R.; Ferrari, M.C.O. Mechanisms underlying the control of responses to predator odours in aquatic prey. J. Exp. Biol. 2017, 220, 1937–1946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakamoto, M.; Chang, K.H.; Hanazato, T. Plastic phenotypes of antennule shape in Bosmina longirostris controlled by physical stimuli from predators. Limnol. Oceanogr. 2007, 52, 2072–2078. [Google Scholar] [CrossRef]
- Tuttle, L.J.; Robinson, H.E.; Takagi, D.; Strickler, J.R.; Lenz, P.H.; Hartline, D.K. Going with the flow: Hydrodynamic cues trigger directed escapes from a stalking predator. J. R. Soc. Interface 2019, 16, 20180776. [Google Scholar] [CrossRef] [Green Version]
- Bernatowicz, P.; Pijanowska, J. Daphnia response to biotic stress is modified by PCBs. Ecotoxicol. Environ. Saf. 2011, 74, 711–718. [Google Scholar] [CrossRef]
- Coors, A.; Hammers-Wirtz, M.; Ratte, H.T. Adaptation to environmental stress in Daphnia magna simultaneously exposed to a xenobiotic. Chemosphere 2004, 56, 395–404. [Google Scholar] [CrossRef]
- DeMille, C.M.; Arnott, S.E.; Pyle, G.G. Variation in copper effects on kairomone-mediated responses in Daphnia pulicaria. Ecotoxicol. Environ. Saf. 2016, 126, 264–272. [Google Scholar] [CrossRef]
- Hunter, K.; Pyle, G. Morphological responses of Daphnia pulex to Chaoborus americanus kairomone in the presence and absence of metals. In Proceedings of the Environmental Toxicology and Chemistry; Wiley: Hoboken, NJ, USA, 2004; Volume 23, pp. 1311–1316. [Google Scholar]
- Mirza, R.S.; Pyle, G.G. Waterborne metals impair inducible defences in Daphnia pulex: Morphology, life-history traits and encounters with predators. Freshw. Biol. 2009, 54, 1016–1027. [Google Scholar] [CrossRef]
- Weiss, L.C.; Pötter, L.; Steiger, A.; Kruppert, S.; Frost, U.; Tollrian, R. Rising pCO2 in Freshwater Ecosystems Has the Potential to Negatively Affect Predator-Induced Defenses in Daphnia. Curr. Biol. 2018, 28, 327–332.e3. [Google Scholar] [CrossRef] [Green Version]
- Bezirci, G.; Akkas, S.B.; Rinke, K.; Yildirim, F.; Kalaylioglu, Z.; Severcan, F.; Beklioglu, M. Impacts of salinity and fish-exuded kairomone on the survival and macromolecular profile of Daphnia pulex. Ecotoxicology 2012, 21, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Riessen, H.P.; Linley, R.D.; Altshuler, I.; Rabus, M.; Söllradl, T.; Clausen-Schaumann, H.; Laforsch, C.; Yan, N.D. Changes in water chemistry can disable plankton prey defenses. Proc. Natl. Acad. Sci. USA 2012, 109, 15377–15382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trotter, B.; Ramsperger, A.F.R.M.; Raab, P.; Haberstroh, J.; Laforsch, C. Plastic waste interferes with chemical communication in aquatic ecosystems. Sci. Rep. 2019, 9, 5889. [Google Scholar] [CrossRef] [PubMed]
- Relyea, R.A. Predators come and predators go: The reversibility of predator-induced traits. Ecology 2003, 84, 1840–1848. [Google Scholar] [CrossRef]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diel, P.; Kiene, M.; Martin-Creuzburg, D.; Laforsch, C. Knowing the Enemy: Inducible Defences in Freshwater Zooplankton. Diversity 2020, 12, 147. https://doi.org/10.3390/d12040147
Diel P, Kiene M, Martin-Creuzburg D, Laforsch C. Knowing the Enemy: Inducible Defences in Freshwater Zooplankton. Diversity. 2020; 12(4):147. https://doi.org/10.3390/d12040147
Chicago/Turabian StyleDiel, Patricia, Marvin Kiene, Dominik Martin-Creuzburg, and Christian Laforsch. 2020. "Knowing the Enemy: Inducible Defences in Freshwater Zooplankton" Diversity 12, no. 4: 147. https://doi.org/10.3390/d12040147
APA StyleDiel, P., Kiene, M., Martin-Creuzburg, D., & Laforsch, C. (2020). Knowing the Enemy: Inducible Defences in Freshwater Zooplankton. Diversity, 12(4), 147. https://doi.org/10.3390/d12040147