A Midsummer Night’s Diet: Snapshot on Trophic Strategy of the Alpine Salamander, Salamandra atra

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Species
2.2. Study Area
2.3. Prey Availability (Potential Trophic Niche)
2.4. Sampling Predators
2.5. Data Analysis
2.5.1. Realized Trophic Niche
2.5.2. Trophic Strategy
2.5.3. Interindividual Diet Variation
3. Results
3.1. Prey Availability (Potential Trophic Niche)
3.2. Sampling Predators
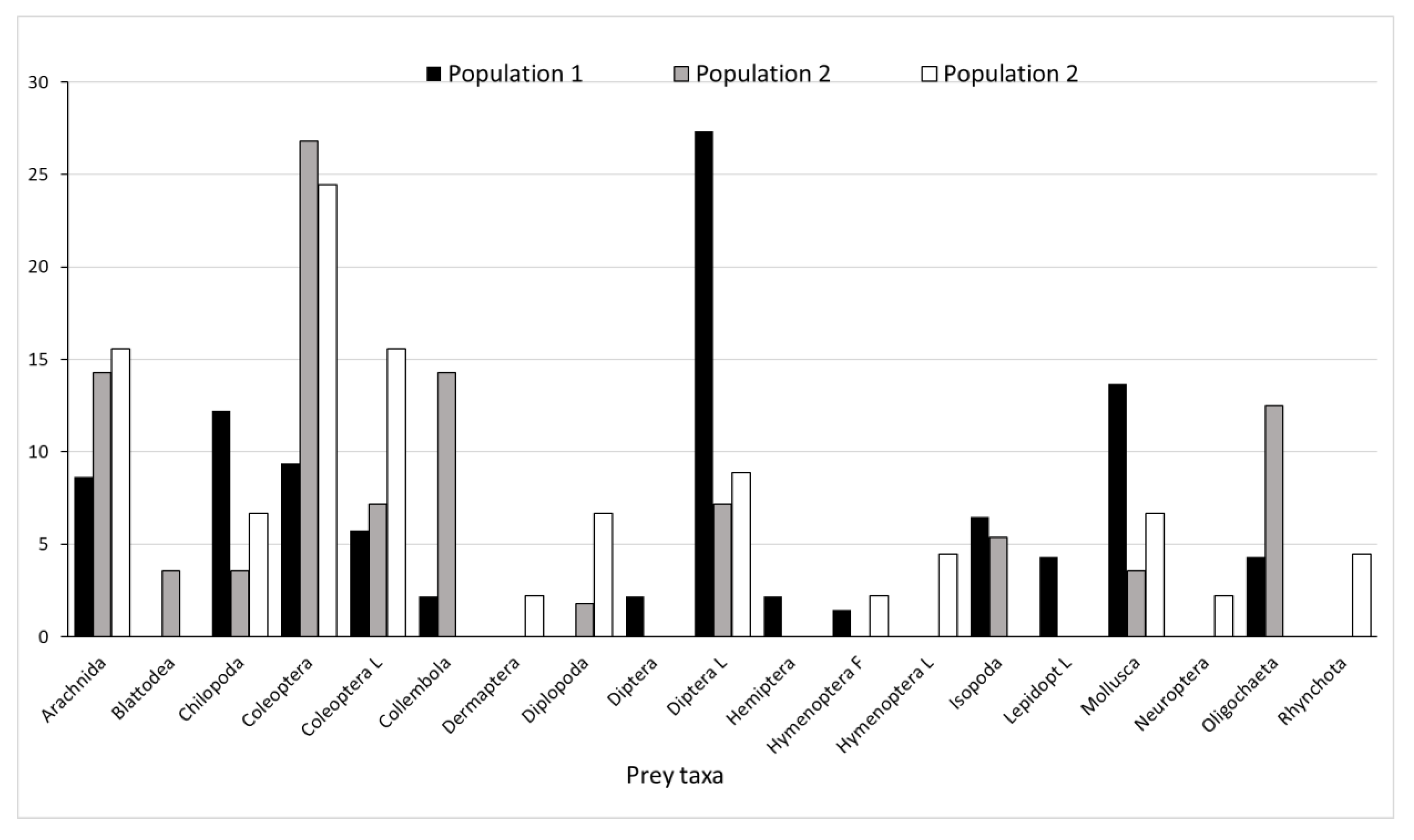
3.3. Realized Trophic Niche
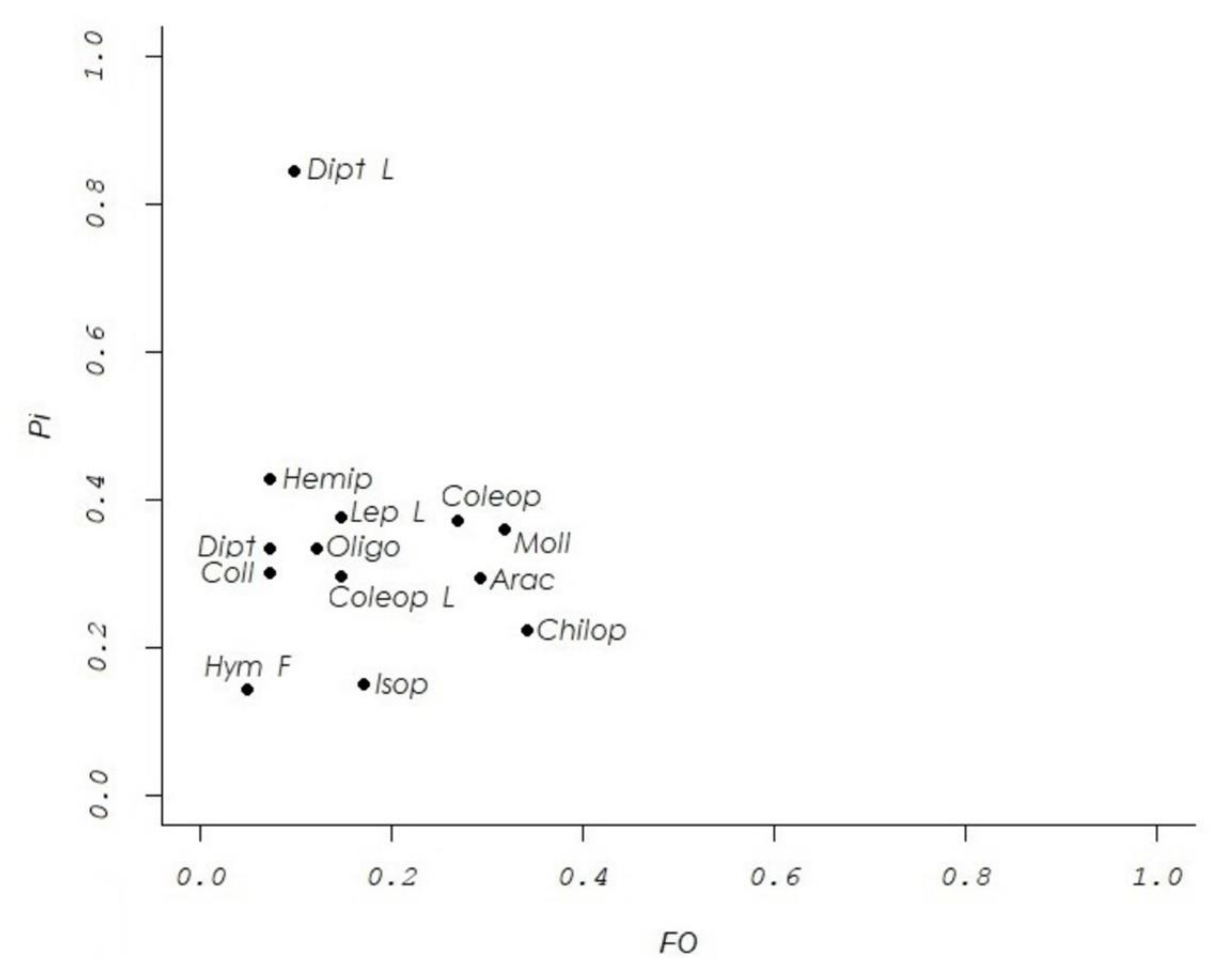
3.4. Analysis of Trophic Strategy
3.5. Analysis of Interindividual Diet Variation
4. Discussion
4.1. Prey Availability
4.2. Realized Trophic Niche
4.3. Trophic Selectivity
4.4. Interindividual Diet Variation
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Davic, R.D.; Welsh, H.H., Jr. On the ecological roles of salamanders. Ann. Rev. Ecol. Evol. System. 2004, 35, 405–434. [Google Scholar] [CrossRef] [Green Version]
- Hairston, N.G. The local distribution and ecology of the plethodontid salamanders of the southern Appalachians. Ecol. Monogr. 1949, 19, 49–73. [Google Scholar] [CrossRef]
- Petranka, J.W. Salamanders of the United States and Canada; Smithsonian Institution Press: Washington, DC, USA, 1998; pp. 1–587. [Google Scholar]
- Costa, A.; Crovetto, F.; Salvidio, S. European plethodontid salamanders on the forest floor: Local abundance is related to fine–scale environmental factors. Herpetol. Conserv. Biol. 2016, 11, 344–349. [Google Scholar]
- Romano, A.; Anderle, M.; Forti, A.; Partel, P.; Pedrini, P. Population density, sex ratio and body size in a population of Salamadra atra atra on the Dolomites. Acta Herpetol. 2018, 13, 195–199. [Google Scholar]
- Burton, T.M.; Likens, G.E. Energy flow and nutrient cycling in salamander populations in the Hubbard Brook Experimental Forest, New Hampshire. Ecology 1975, 56, 1068–1080. [Google Scholar] [CrossRef]
- Best, M.L.; Welsh, H.H., Jr. The trophic role of a forest salamander: Impacts on invertebrates, leaf litter retention, and the humification process. Ecosphere 2014, 5, 1–19. [Google Scholar] [CrossRef]
- Barton, K.A.; Zalewski, A. Winter severity limits red fox populations in Eurasia. Glob. Ecol. Biogeogr. 2007, 16, 281–289. [Google Scholar] [CrossRef]
- Isbell, L.A. Sudden short–term increase in mortality of vervet monkeys (Cercopithecus aethiops) due to leopard predation in Amboseli National Park, Kenya. Am. J. Primatol. 1990, 21, 41–52. [Google Scholar] [CrossRef]
- Berger, L.; Speare, R.; Thomas, A.; Hyatt, A. Mucocutaneous fungal disease in tadpoles of Bufo marinus in Australia. J. Herpetol. 2001, 35, 330–335. [Google Scholar] [CrossRef] [Green Version]
- Chapman, C.A.; Wasserman, M.D.; Gillespie, T.R.; Speirs, M.L.; Lawes, M.J.; Saj, T.L.; Ziegler, T.E. Do food availability, parasitism, and stress have synergistic effects on red colobus populations living in forest fragments? Am. J. Phys Anthropol. 2006, 131, 525–534. [Google Scholar] [CrossRef]
- Hanya, G.; Chapman, C.A. Linking feeding ecology and population abundance: A review of food resource limitation on primates. Ecol. Res. 2013, 28, 183–190. [Google Scholar] [CrossRef] [Green Version]
- Bolnick, D.I.; Yang, L.H.; Fordyce, J.A.; Davis, J.M.; Svanback, R. Measuring individual–level resource specialization. Ecology 2002, 83, 2936–2941. [Google Scholar] [CrossRef]
- Wells, K.D. The Ecology and Behavior of Amphibians; Chicago University Press: Chicago, IL, USA, 2007. [Google Scholar]
- Salvidio, S.; Romano, A.; Oneto, F.; Ottonello, D.; Michelon, R. Different season, different strategies: Feeding ecology of two syntopic forest dwelling salamanders. Acta. Oecol. 2012, 43, 42–50. [Google Scholar]
- Costa, A.; Salvidio, S.; Posillico, M.; Matteucci, G.; De Cinti, B.; Romano, A. Generalisation within specialization: Inter–individual diet variation in the only specialized salamander in the world. Sci. Rep. 2015, 5, 13260. [Google Scholar] [CrossRef] [PubMed]
- Bolnick, D.I.; Svanback, R.; Fordyce, J.A.; Yang, L.H.; Davis, J.M.; Hulsey, C.D.; Forister, M.L. The ecology of individuals: Incidence and implications of individual specialization. Am. Nat. 2003, 161, 1–28. [Google Scholar] [CrossRef]
- Araújo, M.S.; Martins, E.G.; Cruz, L.D.; Fernandes, F.R.; Linhares, A.X.; Dos Reis, S.F.; Guimaraes, P.R., Jr. Nested diets: A novel pattern of individual-level resource use. Oikos 2010, 119, 81–88. [Google Scholar] [CrossRef]
- Salvidio, S.; Oneto, F.; Ottonello, D.; Costa, A.; Romano, A. Trophic specialization at the individual level in a terrestrial generalist salamander. Can. J. Zool. 2015, 93, 79–83. [Google Scholar] [CrossRef]
- Salvidio, S.; Costa, A.; Crovetto, F. Individual trophic specialisation in the Alpine newt increases with increasing resource diversity. Ann. Zool. Fenn. 2019, 56, 17–24. [Google Scholar] [CrossRef]
- Rosa, G.; Costa, A.; Salvidio, S. Trophic strategies of two newt populations living in contrasting habitats on a Mediterranean island. Ethol. Ecol. Evol. 2020, 32, 96–106. [Google Scholar] [CrossRef]
- Costa, A.; Baroni, D.; Romano, A.; Salvidio, S.; Lötters, S. Individual diet variation in Salamandra salamandra larvae in a Mediterranean stream (Amphibia: Caudata). Salamandra 2017, 53, 148–152. [Google Scholar]
- Lanza, B.; Andreone, F.; Bologna, M.A.; Corti, C.; Razzetti, E. Fauna d’Italia, vol. XLII, Amphibia; Calderini Edition: Bologna, Italy, 2007; pp. 197–211. [Google Scholar]
- Sillero, N.; Campos, J.; Bonardi, A.; Corti, C.; Creemers, R.; Crochet, P.A.; Isailovic, J.C.; Denoël, M.; Ficetola, G.F.; Gonçalves, J.; et al. Updated distribution and biogeography of amphibians and reptiles of Europe. Amphib. Reptil. 2014, 35, 1–31. [Google Scholar] [CrossRef] [Green Version]
- Bedriaga, J. von. Die Lurchfauna Europa’s II. Urdodela. Schwanzlurche. Univ. typ. Moscow. Available online: http://www.europeana.eu/en/item/08711/item_23042?fbclid=IwAR368Ijnp_DRjaXfZmkOrOKIrDYic6VsgrfQ6EhAjySPxW9OcsBMZuga4dE (accessed on 16 May 2020).
- Freytag, G.E. Feuersalamander und Alpensalamander. Die Neue Brehm-Bücherei; A. Ziemsen Verlag: Wittenberg Lutherstadt, Germany, 1955; p. 142. [Google Scholar]
- Klewen, R.F. Untersuchungen zur Verbreitung, Öko-Ethologie und innerartlichen Gliederung von Salamandra atra (Laurenti 1768). Ph.D. Thesis, Universität Köln, Köln, Germany, 1986. [Google Scholar]
- Klewen, R.F. Die Landsalamander Europas 1: Die Gattungen Salamandra und Mertensiella; A. Ziemsen Verlag: Wittenberg Lutherstadt, Germany, 1988. [Google Scholar]
- Kuzmin, S.L. Feeding ecology of Salamandra and Mertensiella: A review of data and ontogenetic evolutionary trends. Mertensiella 1994, 4, 271–286. [Google Scholar]
- Fachbach, G.; Kolossau, I.; Ortner, A.; Zur Ernährungsbiologie von Salamandra, s. salamandra und Salamandra atra. Salamandra 1975, 11, 136–144. [Google Scholar]
- Scheller, H.V. Pitfall trapping as the basis for studying ground beetle predation in spring barley. Tidsskr. Planteavl. 1984, 88, 317–324. [Google Scholar]
- Woodcock, B.A. Pitfall trapping in ecological studies. In Insect Sampling in Forest Ecosystems; Leather, S.R., Ed.; Blackwell Publishing Co.: New Jersey, NJ, USA, 2005. [Google Scholar]
- Southwood, T.R.E.; Henderson, P.A. Ecological Methods, 3rd ed.; Wiley-Blackwell: Oxford, UK, 2000; p. 575. [Google Scholar]
- Ausden, M. Invertebrates. In Ecological Census Techniques: A Handbook; Sutherland, W.J., Ed.; Cambridge University Press: Cambridge, UK, 1996. [Google Scholar]
- Duellman, W.E.; Trueb, L. Biology of Amphibians; Johns Hopkins Univ. Press: Baltimore, MD, USA, 1994; p. 670. [Google Scholar]
- Solé, M.; Rödder, D. Dietary assessments of adult amphibians. In Amphibian Ecology and Conservation: A Handbook of Techniques; Dodd, J., Ed.; Oxford University Press: Oxford, UK, 2010. [Google Scholar]
- Fraser, D.F. Coexistence of salamanders in the genus Plethodon: A variation of the Santa Rosalia theme. Ecology 1976, 55, 238–251. [Google Scholar] [CrossRef]
- Salvidio, S. Diet and food utilization in a rockface population of Speleomantes ambrosii (Amphibia, Caudata, Plethodontidae). Vie et Milieu 1992, 42, 35–39. [Google Scholar]
- Schabetsberger, R. Gastric evacuation rates of adult and larval Alpine newts (Triturus alpestris) under laboratory and field conditions. Freshw. Biol. 1994, 31, 143–151. [Google Scholar] [CrossRef]
- Caldwell, J.P. The evolution of myrmecophagy and its correlates in poison frogs (Family Dendrobatidae). J. Zool. 1996, 240, 75–101. [Google Scholar] [CrossRef]
- Solé, M.; Beckmann, O.; Pelz, B.; Kwet, A.; Engels, W. Stomach–flushing for diet analysis in anurans: An improved protocol evaluated in a case study in Araucaria forests, southern Brazil. Stud. Neotrop. Fauna Environ. 2005, 40, 23–28. [Google Scholar] [CrossRef]
- Clarke, K.R. Non–parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Publishing: Oxford, UK, 2004; p. 256. [Google Scholar]
- Wu, Z.-J.; Li, Y.; Wang, Y. A comparison of stomach flush and stomach dissection in diet analysis of four frog species. Acta Zool. Sin. 2007, 53, 364–372. [Google Scholar]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Paleontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Vanderploeg, H.A.; Scavia, D. Calculation and use of selectivity coefficients of feeding: Zooplankton grazing. Ecol. Mod. 1979, 7, 135–149. [Google Scholar] [CrossRef]
- Lechowicz, M.J. The sampling characteristics of electivity indices. Oecologia 1982, 52, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Ramos–Jiliberto, R.; Valdovinos, F.S.; Arias, J.; Alcaraz, C.; García–Berthou, E. A network–based approach to the analysis of ontogenetic diet shifts: An example with an endangered, small–sized fish. Ecol. Compl. 2011, 8, 123–129. [Google Scholar] [CrossRef]
- Costello, M.J. Predator feeding strategy and prey importance: A new graphical analysis. J. Fish Biol. 1990, 36, 261–263. [Google Scholar] [CrossRef]
- Amundsen, P.A.; Gabler, H.M.; Staldvik, F.J. A new approach to graphical analysis of feeding strategy from stomach contents data—Modification of the Costello (1990) method. J. Fish Biol. 1996, 48, 607–614. [Google Scholar]
- Araújo, M.S.; Guimarães, P.R., Jr.; Svanbäck, R.; Pinheiro, A.; Guimarães, P.; Reis, S.F.D.; Bolnick, D.I. Network analysis reveals contrasting effects of intraspecific competition on individual vs. population diets. Ecology 2008, 89, 1981–1993. [Google Scholar] [CrossRef] [Green Version]
- Pires, M.M.; Guimarães, P.R., Jr.; Araújo, M.S.; Giaretta, A.A.; Costa, J.C.L.; Dos Reis, S.F. The nested assembly of individual-resource networks. J. Anim. Ecol. 2011, 80, 896–903. [Google Scholar] [CrossRef]
- Tinker, M.T.; Guimarães, P.R., Jr.; Novak, M.; Marquitt, F.M.D.; Bodkin, J.L.; Staedler, M.; Bentall, G.; Estes, J.A. Structure and mechanism of diet specialisation: Testing models of individual variation in resource use with sea otters. Ecol. Lett. 2012, 15, 475–483. [Google Scholar] [CrossRef]
- Santamaría, S.; Enoksen, C.A.; Olesen, J.M.; Tavecchia, G.; Rotger, A.; Igual, J.M.; Traveset, A. Diet composition of the lizard Podarcis lilfordi (Lacertidae) on 2 small islands: An individual–resource network approach. Curr. Zool. 2020, 66, 39–49. [Google Scholar] [CrossRef]
- Blüthgen, N.; Menzel, F.; Blüthgen, N. Measuring specialization in species interaction networks. BMC Ecol. 2006, 6, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida–Neto, M.; Guimaraes, P.; Guimaraes, P.R., Jr.; Loyola, R.D.; Ulrich, W. A consistent metric for nestedness analysis in ecological systems: Reconciling concept and quantification. Oikos 2008, 117, 1227–1239. [Google Scholar] [CrossRef]
- Beckett, S.J. Improved community detection in weighted bipartite networks. R. Soc. Open Sci. 2016, 3, 140536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aubry, M.P.; Berggren, W.A.; van Couvering, J.; McGowran, B.; Pillans, B.; Hilgen, F.J. Quaternary: Status, rank, definition, survival. Episodes 2005, 28, 118–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sfenthourakis, S.; Anastasiou, I.; Strutenschi, T. The terrestrial isopod diversity of Mt. Panachaiko (Pelopon–nisos, Greece). E. J. Soil Biol. 2005, 41, 91–98. [Google Scholar] [CrossRef]
- Lessard, S.J.; Rivas, D.A.; Stephenson, E.J.; Yaspelkis, B.B., III; Koch, L.G.; Britton, S.L.; Hawley, J.A. Exercise training reverses impaired skeletal muscle metabolism induced by artificial selection for low aerobic capacity. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R175–R182. [Google Scholar] [CrossRef] [Green Version]
- Lamb, T. The influence of sex and breeding condition on microhabitat selection and diet in the Pig Frog Rana grylio. Am. Mid. Nat. 1984, 111, 311–318. [Google Scholar] [CrossRef]
- Romano, A.; Salvidio, S.; Palozzi, R.; Sbordoni, V. Diet of the newt, Triturus carnifex (Laurenti, 1768), in a flooded karstic sinkhole ("Pozzo del Merro", Central Italy). J. Cave Karst Stud. 2012, 74, 271–277. [Google Scholar] [CrossRef]
- Duellman, W.E.; Trueb, L. Biology of Amphibians; Mc Graw-Hill Book Company: New York, NY, USA, 1986. [Google Scholar]
- Whitaker, J.O., Jr.; Rubin, D.C. Food habits of Plethodon jordani metcalfi and Plethodon jordani shermani from North Carolina. Herpetologica 1971, 27, 81–86. [Google Scholar]
- Krebs, J.R. Optimal foraging: Decision rules for predators. In Behavioural Ecology: An Evolutionary Approach; Krebs, J.R., Davies, N.B., Eds.; Backwell Scientific Publications: Oxford, UK, 1978; pp. 23–63. [Google Scholar]
- Stephens, D.W. Foraging Theory; Princeton University Press: Princeton, NJ, USA, 1986. [Google Scholar]
- Schoener, T.W. Theory of feeding strategies. Annu. Rev. Ecol. Syst. 1971, 2, 369–404. [Google Scholar] [CrossRef] [Green Version]
- Pulliam, H.R. On the theory of optimal diets. Am. Nat. 1974, 108, 59–75. [Google Scholar] [CrossRef]
- Svanbäck, R.; Bolnick, D.I. Intraspecific competition affects the strength of individual specialization: An optimal diet theory method. Evol. Ecol. Res. 2005, 7, 993–1012. [Google Scholar]
- Speybroeck, J.; Beukema, W.; Bok, B.; Van Der Voort, J. Field Guide to the Amphibians and Reptiles of Britain and Europe; Bloomsbury Natural History: London, UK, 2016. [Google Scholar]
- Razpet, A.; Šunje, E.; Kalamujić, B.; Tulić, U.; Pojskić, N.; Stanković, D.; Krizmanić5, I.; Marić, S. Genetic differentiation and population dynamics of Alpine salamanders (Salamandra atra, Laurenti 1768) in Southeastern Alps and Dinarides. Herpetol. J. 2016, 26, 111–119. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Invertebrate Taxa | Preyed Invertebrates | Environmental Availability of Invertebrates | ||
|---|---|---|---|---|
| Stomach Contents | Diurnal Traps (DT) | Nocturnal Traps (NT) | Pool captures (DT + NT) | |
| Arachnida | 12 | 37 | 17 | 54 |
| Chilopoda | 17 | 5 | 3 | 8 |
| Coleoptera | 13 | 23 | 6 | 29 |
| Coleoptera larvae | 8 | 6 | 7 | 13 |
| Collembola | 3 | 74 | 101 | 175 |
| Dermaptera | 0 | 1 | 1 | 2 |
| Diptera | 3 | 66 | 12 | 78 |
| Diptera larvae | 38 | 0 | 1 | 1 |
| Formicidae | 0 | 93 | 61 | 154 |
| Hemiptera | 0 | 13 | 6 | 19 |
| Isopoda | 9 | 6 | 14 | 20 |
| Lepidoptera | 0 | 2 | 1 | 3 |
| Lepidoptera larvae | 6 | 1 | 1 | 2 |
| Mecoptera larvae | 0 | 0 | 5 | 5 |
| Mollusca | 19 | 2 | 17 | 19 |
| Oligochaeta | 6 | 1 | 1 | 2 |
| Orthoptera | 0 | 2 | 0 | 2 |
| Rynchota | 0 | 2 | 0 | 2 |
| winged Hymenoptera | 2 | 4 | 1 | 5 |
| Simpson Diversity Index | 1-D (95% C.I.) | Taxa |
|---|---|---|
| Diurnal traps (DT) | 0.82 (0.80–0.83) | 17 |
| Nocturnal traps (NT) | 0.76 (0.72–0.80) | 17 |
| Pooled captures (i.e., DT + NT) | 0.82 (0.80–0.83) | 19 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roner, L.; Costa, A.; Pedrini, P.; Matteucci, G.; Leonardi, S.; Romano, A. A Midsummer Night’s Diet: Snapshot on Trophic Strategy of the Alpine Salamander, Salamandra atra. Diversity 2020, 12, 202. https://doi.org/10.3390/d12050202
Roner L, Costa A, Pedrini P, Matteucci G, Leonardi S, Romano A. A Midsummer Night’s Diet: Snapshot on Trophic Strategy of the Alpine Salamander, Salamandra atra. Diversity. 2020; 12(5):202. https://doi.org/10.3390/d12050202
Chicago/Turabian StyleRoner, Luca, Andrea Costa, Paolo Pedrini, Giorgio Matteucci, Stefano Leonardi, and Antonio Romano. 2020. "A Midsummer Night’s Diet: Snapshot on Trophic Strategy of the Alpine Salamander, Salamandra atra" Diversity 12, no. 5: 202. https://doi.org/10.3390/d12050202
APA StyleRoner, L., Costa, A., Pedrini, P., Matteucci, G., Leonardi, S., & Romano, A. (2020). A Midsummer Night’s Diet: Snapshot on Trophic Strategy of the Alpine Salamander, Salamandra atra. Diversity, 12(5), 202. https://doi.org/10.3390/d12050202