Spatial Segregation between Red Foxes (Vulpes vulpes), European Wildcats (Felis silvestris) and Domestic Cats (Felis catus) in Pastures in a Livestock Area of Northern Spain

 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
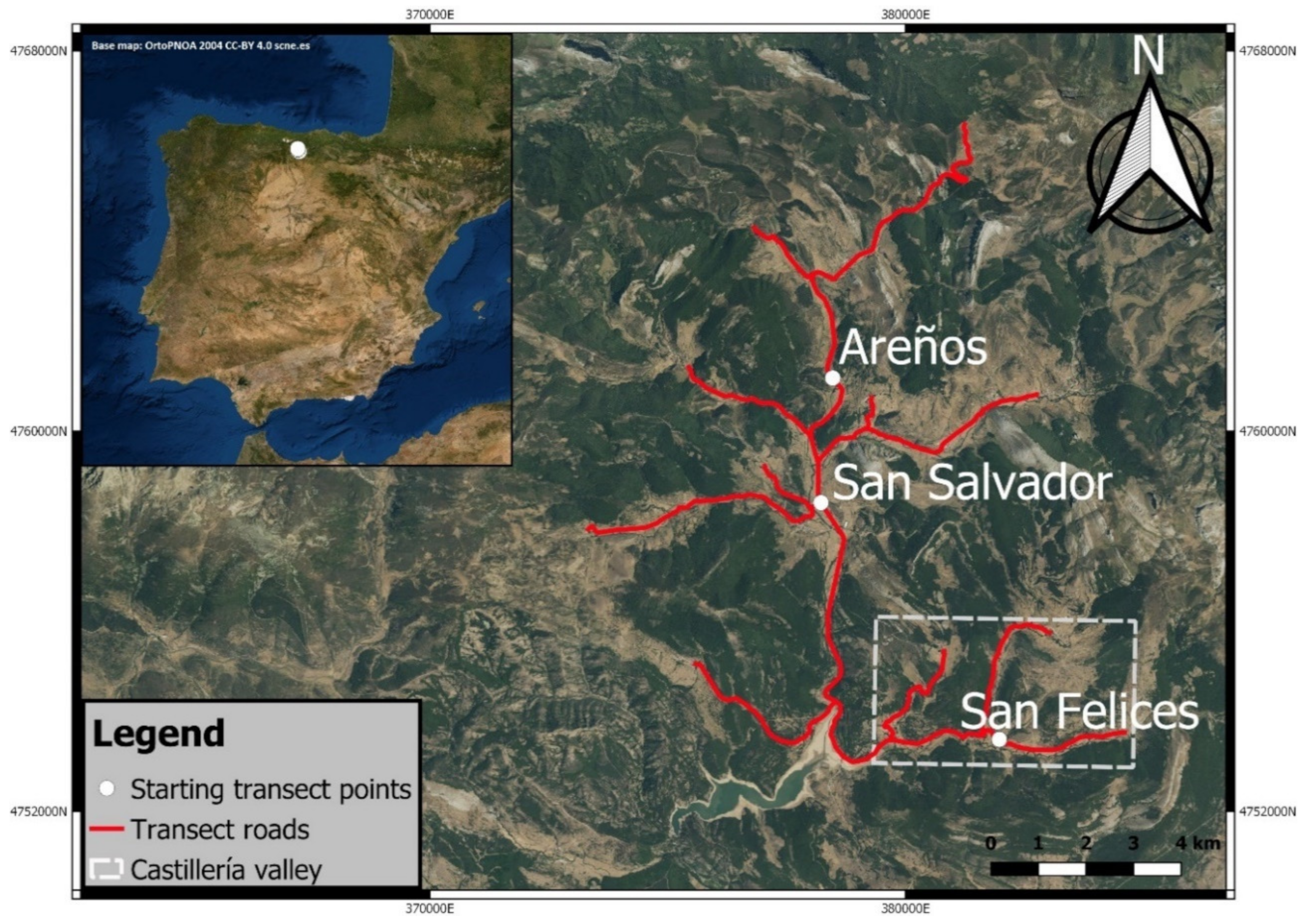
2.1. Study Area
2.2. Study Design
2.3. Selection of Sites on Pastures by Carnivores
2.4. Spatial Segregation among Carnivores
2.5. Prey Abundance in Places within Pastures Used by Carnivores
3. Results
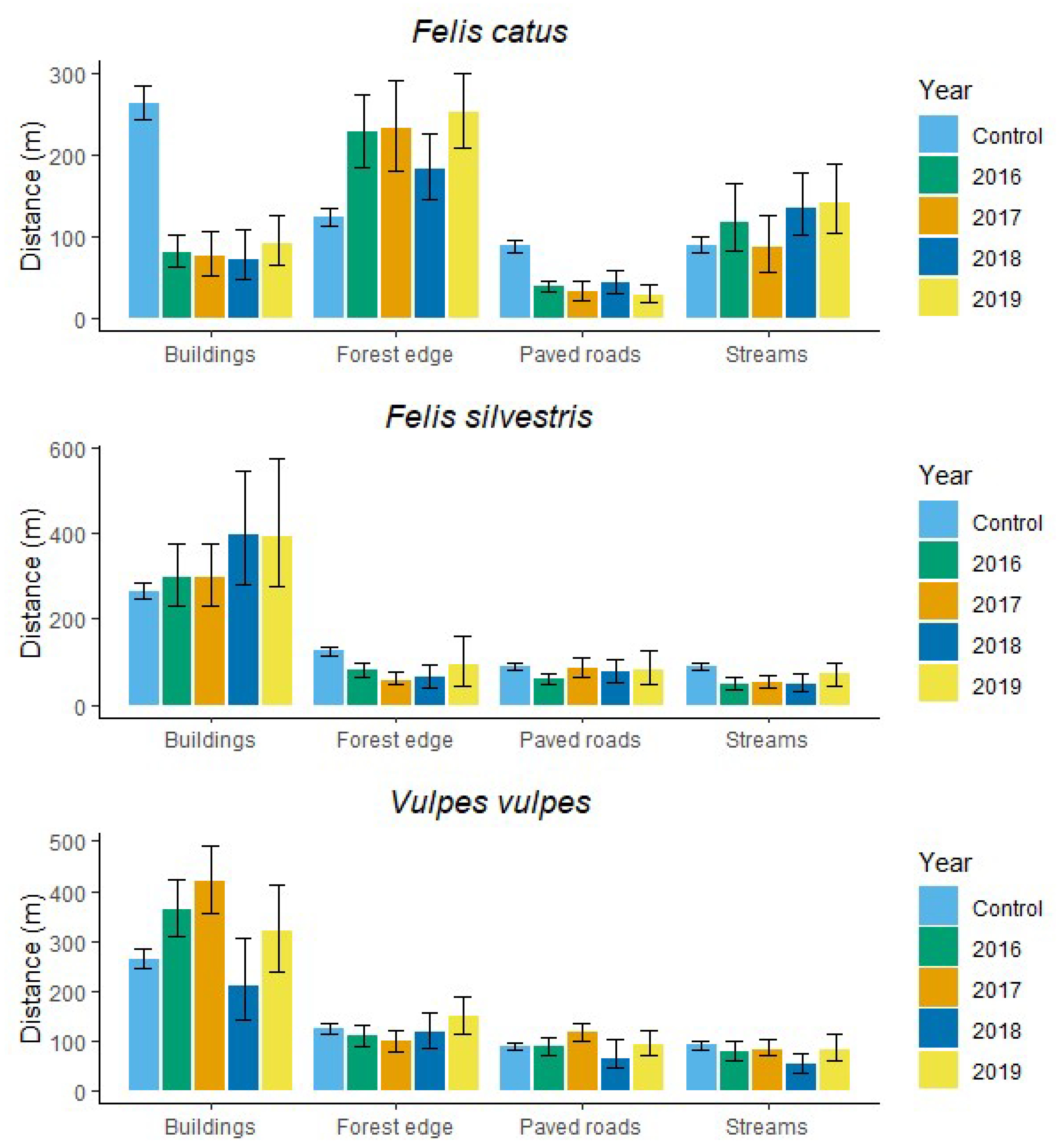
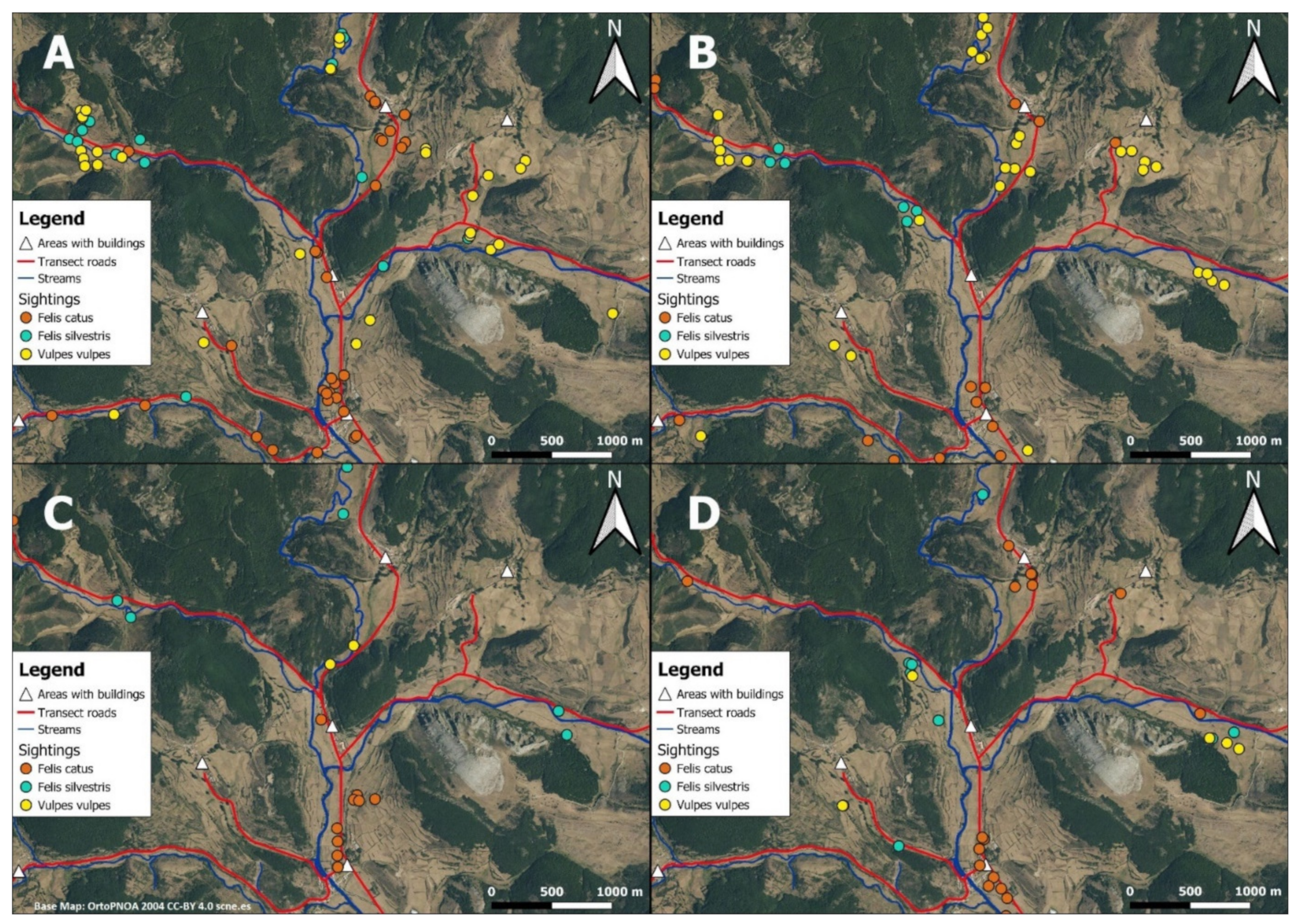
3.1. Selection of Sites within Pastures by Carnivores
3.2. Spatial Segregation among Carnivores
3.3. Prey Abundance within Pasture Areas Used by Carnivores
4. Discussion
4.1. Sites Selected in Pastures
4.2. Spatial Segregation
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Treves, A.; Karanth, K.U. Human-Carnivore Conflict and Perspectives on Carnivore Management Worldwide. Conserv. Biol. 2003, 17, 1491–1499. [Google Scholar] [CrossRef]
- Prugh, L.R.; Stoner, C.J.; Epps, C.W.; Bean, W.T.; Ripple, W.J.; Laliberte, A.S.; Brashares, J.S. The Rise of the Mesopredator. BioScience 2009, 59, 779–791. [Google Scholar] [CrossRef]
- Roemer, G.W.; Gompper, M.E.; Valkenburgh, B.V.A.N. The Ecological Role of the Mammalian Mesocarnivore. BioScience 2009, 59, 165–173. [Google Scholar] [CrossRef]
- Creel, S.; Spong, G.; Creel, N. Interspecific competition and the population biology of extinction-prone carnivores. In Carnivore Conservation; Gittleman, J.L., Funk, S.M., Macdonald, D.W., Wayne, R.K., Eds.; Cambridge University Press: Cambridge, UK, 2001; pp. 35–60. [Google Scholar]
- Polis, G.A.; Myers, C.A.; Holt, R.D. The Ecology and Evolution of Intraguild Predation: Potential Competitors that Eat Each Other. Annu. Rev. Ecol. Syst. 1989, 20, 297–330. [Google Scholar] [CrossRef]
- Palomares, F.; Caro, T.M. Interspecific killing among mammalian carnivores. Am. Nat. 1999, 153, 492–508. [Google Scholar] [CrossRef]
- Vanak, A.T.; Fortin, D.; Thaker, M.; Ogden, M.; Owen, C.; Greatwood, S.; Slotow, R. Moving to stay in place: Behavioral mechanisms for coexistence of African large carnivores. Ecology 2013, 94, 2619–2631. [Google Scholar] [CrossRef]
- Hunter, J.S.; Durant, S.M.; Caro, T.M. To flee or not to flee: Predator avoidance by cheetahs at kills. Behav. Ecol. Sociobiol. 2007, 61, 1033–1042. [Google Scholar] [CrossRef]
- Rostro-García, S.; Kamler, J.F.; Hunter, L.T.B. To kill, stay or flee: The effects of lions and landscape factors on habitat and kill site selection of cheetahs in South Africa. PLoS ONE 2015, 10, e0117743. [Google Scholar] [CrossRef] [Green Version]
- Schoener, T.W. Resource partitioning in ecological communities. Science 1974, 185, 27–39. [Google Scholar] [CrossRef]
- Karanth, K.U.; Sunquist, M.E. Prey Selection by Tiger, Leopard and Dhole in Tropical Forests. J. Anim. Ecol. 1995, 64, 439–450. [Google Scholar] [CrossRef]
- Durant, S.M. Competition refuges and coexistence: An example from Serengeti carnivores. J. Anim. Ecol. 1998, 67, 370–386. [Google Scholar] [CrossRef] [Green Version]
- Monterroso, P.; Alves, P.C.; Ferreras, P. Plasticity in circadian activity patterns of mesocarnivores in Southwestern Europe: Implications for species coexistence. Behav. Ecol. Sociobiol. 2014, 68, 1403–1417. [Google Scholar] [CrossRef] [Green Version]
- Hemami, M.R.; Watkinson, A.R.; Dolman, P.M. Habitat selection by sympatric muntjac (Muntiacus reevesi) and roe deer (Capreolus capreolus) in a lowland commercial pine forest. For. Ecol. Manag. 2004, 194, 49–60. [Google Scholar] [CrossRef]
- Soto, C.; Palomares, F. Coexistence of sympatric carnivores in relatively homogeneous Mediterranean landscapes: Functional importance of habitat segregation at the fine-scale level. Oecologia 2015, 179, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Blondel, J. The “design” of Mediterranean landscapes: A millennial story of humans and ecological systems during the historic period. Hum. Ecol. 2006, 34, 713–729. [Google Scholar] [CrossRef]
- Schuette, P.; Wagner, A.P.; Wagner, M.E.; Creel, S. Occupancy patterns and niche partitioning within a diverse carnivore community exposed to anthropogenic pressures. Biol. Conserv. 2013, 158, 301–312. [Google Scholar] [CrossRef] [Green Version]
- Marzluff, J.M. Worldwide urbanization and its effects on birds. In Avian Ecology and Conservation in an Urbanizing World; Marzluff, J.M., Bowman, R., Donnelly, R., Eds.; Springer: Boston, MA, USA, 2001; pp. 19–47. [Google Scholar]
- Fraterrigo, J.M.; Wiens, J.A. Bird communities of the Colorado Rocky Mountains along a gradient of exurban development. Landsc. Urban Plan. 2005, 71, 263–275. [Google Scholar] [CrossRef]
- Levin, S.A. Dispersion and population interactions. Am. Nat. 1974, 108, 207–228. [Google Scholar] [CrossRef]
- Rosenzweig, M.L.; Abramsky, Z.; Brand, S. Estimating species interactions in heterogeneous environments. Oikos 1984, 43, 329–340. [Google Scholar] [CrossRef]
- Pereira, P.; Alves da Silva, A.; Alves, J.; Matos, M.; Fonseca, C. Coexistence of carnivores in a heterogeneous landscape: Habitat selection and ecological niches. Ecol. Res. 2012, 27, 745–753. [Google Scholar] [CrossRef]
- Amores, F. Diet of the red fox (Vulpes vulpes) in the western Sierra Morena (south Spain). Doñana Acta Vertebr. 1975, 2, 221–229. [Google Scholar]
- Biró, Z.; Lanszki, J.; Szemethy, L.; Heltai, M.; Randi, E. Feeding habits of feral domestic cats (Felis catus), wild cats (Felis silvestris) and their hybrids: Trophic niche overlap among cat groups in Hungary. J. Zool. 2005, 266, 187–196. [Google Scholar] [CrossRef]
- Hartová-Nentvichová, M.; Šálek, M.; Červený, J.; Koubek, P. Variation in the diet of the red fox (Vulpes vulpes) in mountain habitats: Effects of altitude and season. Mamm. Biol. 2009, 75, 334–340. [Google Scholar] [CrossRef]
- Lozano, J.; Malo, A. Conservation of the European Wildcat (Felis silvestris) in Mediterranean Environments: A Reassessment of Current Threats. In Mediterranean Ecosystems: Dynamics, Management and Conservation; Nova Science Publisher’s, Inc.: Hauppauge, NY, USA, 2012; pp. 1–31. ISBN 978-1-61209-146-4. [Google Scholar]
- Széles, G.L.; Purger, J.J.; Molnár, T.; Lanszki, J. Comparative analysis of the diet of feral and house cats and wildcat in Europe. Mammal Res. 2018, 63, 43–53. [Google Scholar] [CrossRef]
- Román, J.; Fermín, U.; Jubete, F.; Revilla, E.; Palomares, F. Definiendo el Hábitat de la Rata Topera (Arvicola Scherman) en la Cordillera Cantábrica. Presented at the XIII Congreso SECEM, Guadalajara, Spain, 6–9 December 2017. [Google Scholar]
- Risbey, D.A.; Calver, M.C.; Short, J.; Bradley, J.S.; Wright, I.W. The impact of cats and foxes on the small vertebrate fauna of Heirisson Prong, Western Australia. II. A field experiment. Wildl. Res. 2000, 27, 223–235. [Google Scholar] [CrossRef] [Green Version]
- Bakaloudis, D.; Bontzorlos, V.; Vlachos, C.; Papakosta, M.; Chatzinikos, E.; Braziotis, S.; Kontsiotis, V. Factors affecting the diet of the red fox (Vulpes vulpes) in a heterogeneous Mediterranean landscape. Turk. J. Zool. 2015, 39. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, J.P.; Leitão, I.; Santos-Reis, M.; Revilla, E. Human-related factors regulate the spatial ecology of domestic cats in sensitive areas for conservation. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [Green Version]
- Molsher, R.L. The Ecology of Feral Cats, Felis catus, in Open Forest in New South Wales: Interactions with Food Resources and Foxes; School of Biological Sciences, University of Sydney: Sydney, NSW, Australia, 1999. [Google Scholar]
- Martínez, S.R.; Arregui, J.J.L. Bioclimatology of the Iberian peninsula. Itinera Geobot. 1999, 13, 41–47. [Google Scholar]
- Ortega Villazán, M.T.; Morales Rodríguez, C. El Clima de la Cordillera Cantábrica Castellano-Leonesa: Diversidad, Contrastes y Cambios. Investig. Geográficas 2015, 63, 45–67. [Google Scholar] [CrossRef] [Green Version]
- Stahl, P.R.; Léger, F. Le Chat Sauvage d’Europe:(Felis silvestris Schreber, 1777); Société Française pour L’étude et la Protection des Mammifères: Pezenas, France, 1992. [Google Scholar]
- Mead-Briggs, A.; Woods, J. An index of activity to assess the reduction in mole numbers caused by control measures. J. Appl. Ecol. 1973, 10, 837–845. [Google Scholar] [CrossRef]
- Giraudoux, P.; Pradier, B.; Delattre, P.; Deblay, S.; Salvi, D.; Defaut, R. Estimation of water vole abundance by using surface indices. Acta Theriol. 1995, 40, 77. [Google Scholar] [CrossRef] [Green Version]
- Santos, S.M.; Mira, A.P.; Mathias, M.L. Using presence signs to discriminate between similar species. Integr. Zool. 2009, 4, 258–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- QGIS Development Team. QGIS Geographic Information System; QGIS. 2009. Available online: https://www.qgis.org/en/site/ (accessed on 18 May 2020).
- Palomares, F.; Adrados, B.; Zanin, M.; Silveira, L.; Keller, C. A non-invasive faecal survey for the study of spatial ecology and kinship of solitary felids in the Viruá National Park, Amazon Basin. Mammal Res. 2017, 62, 241–249. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Jerosch, S.; Kramer-Schadt, S.; Götz, M.; Roth, M. The importance of small-scale structures in an agriculturally dominated landscape for the European wildcat (Felis silvestris silvestris) in central Europe and implications for its conservation. J. Nat. Conserv. 2018, 41, 88–96. [Google Scholar] [CrossRef]
- Klar, N.; Fernández, N.; Kramer-Schadt, S.; Herrmann, M.; Trinzen, M.; Büttner, I.; Niemitz, C. Habitat selection models for European wildcat conservation. Biol. Conserv. 2008, 141, 308–319. [Google Scholar] [CrossRef] [Green Version]
- Beutel, T.; Reineking, B.; Tiesmeyer, A.; Nowak, C.; Heurich, M. Spatial patterns of co-occurrence of the European wildcat Felis silvestris silvestris and domestic cats Felis silvestris catus in the Bavarian Forest National Park. Wildl. Biol. 2017, 2017, wlb.00284. [Google Scholar] [CrossRef] [Green Version]
- Wittmer, H.U. Home range size, movements, and habitat utilization of three male European wildcats (Felis silvestris Schreber, 1777) in Saarland and Rheinland-Pfalz (Germany). Mamm. Biol. Z. Saugetierkd. 2001, 66, 365–370. [Google Scholar]
- Jerosch, S.; Götz, M.; Klar, N.; Roth, M. Characteristics of diurnal resting sites of the endangered European wildcat (Felis silvestris silvestris): Implications for its conservation. J. Nat. Conserv. 2010, 18, 45–54. [Google Scholar] [CrossRef]
- Doyle, A.T. Use of riparian and upland habitats by small mammals. J. Mammal. 1990, 71, 14–23. [Google Scholar] [CrossRef]
- Osbourne, J.D.; Anderson, J.T.; Spurgeon, A.B. Effects of habitat on small-mammal diversity and abundance in West Virginia. Wildl. Soc. Bull. 2005, 33, 814–822. [Google Scholar] [CrossRef]
- Sullivan, T.P.; Sullivan, D.S. Plant and small mammal diversity in orchard versus non-crop habitats. Agric. Ecosyst. Environ. 2006, 116, 235–243. [Google Scholar] [CrossRef]
- Dieterlen, F. Schermaus (Ostschermaus, Große Wühlmaus) Arvicola terrestris (Linnaeus, 1758). In Die Säugetiere Baden-Württembergs; Eugen Ulmer: Stuttgart, Germany, 2005; pp. 328–341. [Google Scholar]
- Recio, M.R.; Arija, C.M.; Cabezas-Díaz, S.; Virgós, E. Changes in Mediterranean Mesocarnivore Communities along Urban and Ex-Urban Gradients. Curr. Zool. 2015, 61, 793–801. [Google Scholar] [CrossRef] [Green Version]
- Easterbee, N.; Hepburn, L.V.; Jefferies, D.J. Survey of the Status and Distribution of the Wildcat in Scotland, 1983–1987; Nature Conservancy Council for Scotland: Edinburgh, Scotland, 1991.
- Lozano, J.; Virgós, E.; Malo, A.F.; Huertas, D.L.; Casanovas, J.G. Importance of scrub-pastureland mosaics for wild-living cats occurrence in a Mediterranean area: Implications for the conservation of the wildcat (Felis silvestris). Biodivers. Conserv. 2003, 12, 921–935. [Google Scholar] [CrossRef]
- Silva, A.P.; Kilshaw, K.; Johnson, P.J.; MacDonald, D.W.; Rosalino, L.M. Wildcat occurrence in Scotland: Food really matters. Divers. Distrib. 2013, 19, 232–243. [Google Scholar] [CrossRef]
- Goszczyński, J.; Krauze, D.; Gryz, J. Activity and exploration range of house cats in rural areas of central Poland. Folia Zool. 2009, 58, 363–371. [Google Scholar]
- Krauze-Gryz, D.; Gryz, J.B.; Goszczyński, J.; Chylarecki, P.; Zmihorski, M. The good, the bad, and the ugly: Space use and intraguild interactions among three opportunistic predators—Cat (Felis catus), dog (Canis lupus familiaris), and red fox (Vulpes vulpes)—Under human pressure. Can. J. Zool. 2012, 90, 1402–1413. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, B.M. Diet of domestic cats and their impact on prey populations. In Domestic Cat: The Biology of Its Behavior; Turner, D.C., Bateson, P., Eds.; Cambridge University Press: Cambridge, UK, 1988. [Google Scholar]
- Woods, M.; McDonald, R.A.; Harris, S. Predation of wildlife by domestic cats Felis catus in Great Britain. Mammal Rev. 2003, 33, 174–188. [Google Scholar] [CrossRef] [Green Version]
- Crooks, K. Relative Sensitivities of Mammalian Carnivores to Habitat Fragmentation. Conserv. Biol. 2002, 16, 488–502. [Google Scholar] [CrossRef]
- Pyšková, K.; Kauzál, O.; Storch, D.; Horáček, I.; Pergl, J.; Pyšek, P. Carnivore distribution across habitats in a central-European landscape: A camera trap study. ZooKeys 2018, 770, 227–246. [Google Scholar] [CrossRef]
- Cavallini, P.; Lovari, S. Environmental factors influencing the use of habitat in the red fox, Vulpes vulpes. J. Zool. 1991, 223, 323–339. [Google Scholar] [CrossRef]
- Delibes-Mateos, M.; Fernandez De Simon, J.; Villafuerte, R.; Ferreras, P. Feeding responses of the red fox (Vulpes vulpes) to different wild rabbit (Oryctolagus cuniculus) densities: A regional approach. Eur. J. Wildl. Res. 2008, 54, 71–78. [Google Scholar] [CrossRef]
- Harris, S. An estimation of the number of foxes (Vulpes vulpes) in the city of Bristol, and some possible factors affecting their distribution. J. Appl. Ecol. 1981, 18, 455–465. [Google Scholar] [CrossRef]
- Sorace, A.; Gustin, M. Distribution of generalist and specialist predators along urban gradients. Landsc. Urban Plan. 2009, 90, 111–118. [Google Scholar] [CrossRef]
- Červinka, J.; Drahníková, L.; Kreisinger, J.; Šálek, M. Effect of habitat characteristics on mesocarnivore occurrence in urban environment in the Central Europe. Urban Ecosyst. 2014, 17, 893–909. [Google Scholar] [CrossRef]
- Alexandre, M.; Hipólito, D.; Ferreira, E.; Fonseca, C.; Rosalino, L.M. Humans do matter: Determinants of red fox (Vulpes vulpes) presence in a western Mediterranean landscape. Mamm. Res. 2020, 65, 203–214. [Google Scholar] [CrossRef]
- Baker, P.J.; Soulsbury, C.D.; Iossa, G.; Harris, S. Domestic cat (Felis catus) and domestic dog (Canis familiaris). In Urban Carnivores, Ecology, Conflict, and Conservation; Gehrt, S.D., Riley, S.P.D., Cypher, B.L., Eds.; John Hopkins University Press: Baltimore, MD, USA, 2010; pp. 157–172. ISBN 978-0-8018-9389-6. [Google Scholar]
- Figueiredo, A.M.; Valente, A.M.; Barros, T.; Carvalho, J.; Silva, D.A.M.; Fonseca, C.; de Carvalho, L.M.; Torres, R.T. What does the wolf eat? Assessing the diet of the endangered Iberian wolf (Canis lupus signatus) in northeast Portugal. PLoS ONE 2020, 15, 1–15. [Google Scholar] [CrossRef]
- Barja, I. Prey and Prey-Age Preference by the Iberian Wolf Canis Lupus Signatus in a Multiple-Prey Ecosystem. Wildl. Biol. 2009, 15, 147–154. [Google Scholar] [CrossRef]
- Germain, E.; Benhamou, S.; Poulle, M.L. Spatio-temporal sharing between the European wildcat, the domestic cat and their hybrids. J. Zool. 2008, 276, 195–203. [Google Scholar] [CrossRef]
- Gil-Sánchez, J.M.; Jaramillo, J.; Barea-Azcón, J.M. Strong spatial segregation between wildcats and domestic cats may explain low hybridization rates on the Iberian Peninsula. Zoology 2015, 118, 377–385. [Google Scholar] [CrossRef]
- Mori, E.; Bagnato, S.; Serroni, P.; Sangiuliano, A.; Rotondaro, F.; Marchianò, V.; Cascini, V.; Poerio, L.; Ferretti, F. Spatiotemporal mechanisms of coexistence in an European mammal community in a protected area of southern Italy. J. Zool. 2020, 310, 232–245. [Google Scholar] [CrossRef]
- Nowell, K.; Jackson, P. Wild Cats: Status Survey and Conservation Action Plan; IUCN: Gland, Switzerland, 1996; Volume 382. [Google Scholar]
- Beaumont, M.; Barratt, E.M.; Gottelli, D.; Kitchener, A.C.; Daniels, M.J.; Pritchard, J.K.; Bruford, M.W. Genetic diversity and introgression in the Scottish wildcat. Mol. Ecol. 2001, 10, 319–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecis, R.; Pierpaoli, M.; Birò, Z.S.; Szemethy, L.; Ragni, B.; Vercillo, F.; Randi, E. Bayesian analyses of admixture in wild and domestic cats (Felis silvestris) using linked microsatellite loci. Mol. Ecol. 2006, 15, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Hertwig, S.T.; Schweizer, M.; Stepanow, S.; Jungnickel, A.; Böhle, U.R.; Fischer, M.S. Regionally high rates of hybridization and introgression in German wildcat populations (Felis silvestris, Carnivora, Felidae). J. Zool. Syst. Evol. Res. 2009, 47, 283–297. [Google Scholar] [CrossRef]
- O’Brien, J.; Devillard, S.; Say, L.; Vanthomme, H.; Léger, F.; Ruette, S.; Pontier, D. Preserving genetic integrity in a hybridising world: Are European Wildcats (Felis silvestris silvestris) in eastern France distinct from sympatric feral domestic cats? Biodivers. Conserv. 2009, 18, 2351–2360. [Google Scholar] [CrossRef]
- Ruiz-García, M.; García-Perea, R.; García, F.J.; Guzmán, J. Primeros resultados sobre el análisis genético de poblaciones españolas de gato montés (Felis silvestris) y su posible hibridación con gatos domésticos (Felis catus). Presented at the V Jornadas SECEM, Vitoria, Spain, 3–5 October 2001. [Google Scholar]
- Oliveira, R.; Godinho, R.; Randi, E.; Ferrand, N.; Alves, P.C. Molecular analysis of hybridisation between wild and domestic cats (Felis silvestris) in Portugal: Implications for conservation. Conserv. Genet. 2008, 9, 1–11. [Google Scholar] [CrossRef]
- Oliveira, R.; Godinho, R.; Randi, E.; Alves, P.C. Hybridization versus conservation: Are domestic cats threatening the genetic integrity of wildcats (Felis silvestris silvestris) in Iberian Peninsula? Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 2953–2961. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
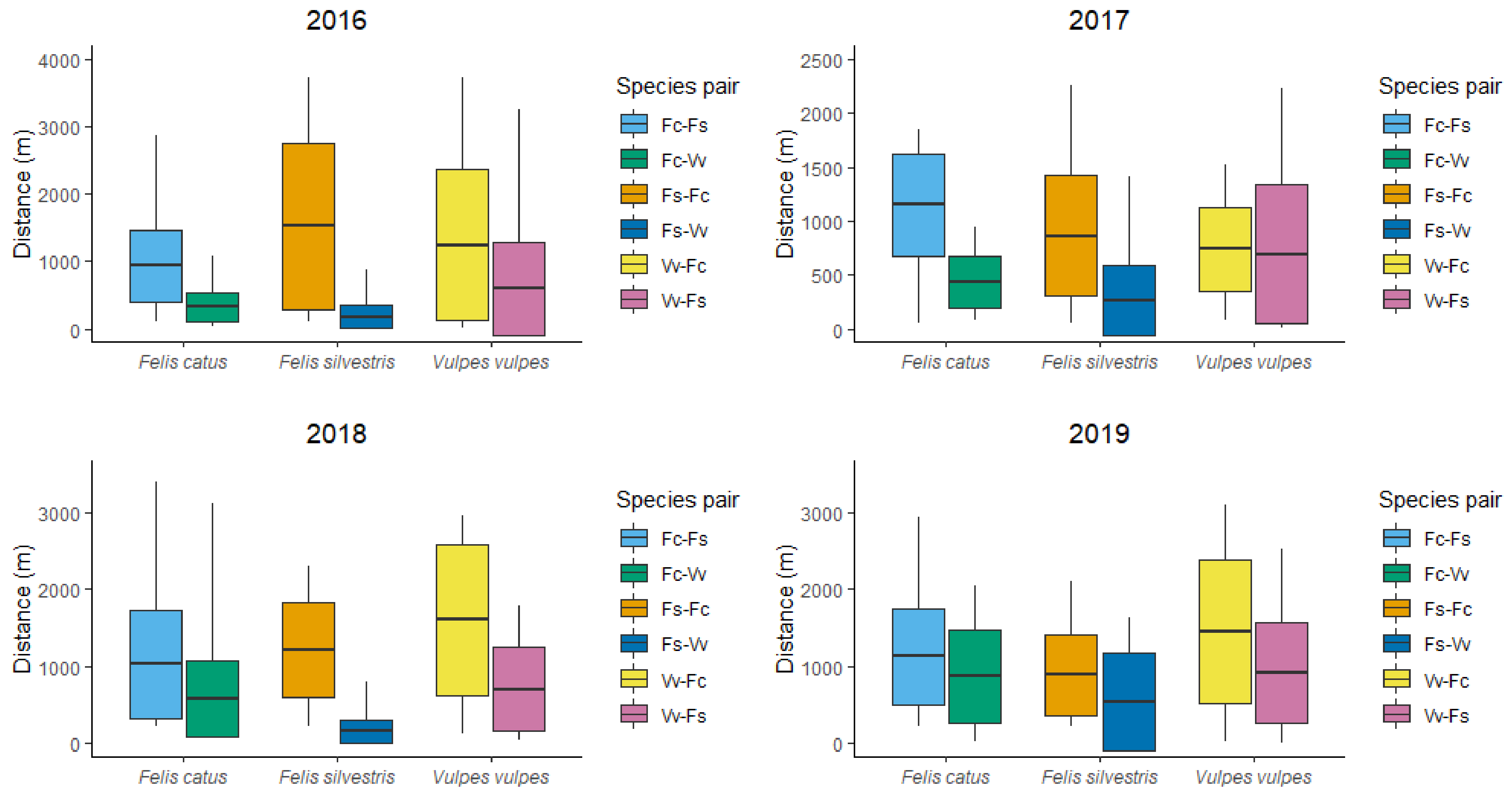{kind=link}
| Year | Period of the Day | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Species | 2016 | 2017 | 2018 | 2019 | Total | Morning | Midday | Evening | Night | Total |
| Felis catus | 63 | 33 | 34 | 46 | 176 | 72 | 9 | 90 | 5 | 176 |
| Felis silvestris | 51 | 53 | 29 | 14 | 147 | 69 | 2 | 44 | 32 | 147 |
| Vulpes vulpes | 100 | 87 | 33 | 39 | 259 | 91 | 2 | 58 | 108 | 259 |
| Total | 214 | 173 | 96 | 99 | 582 | 232 | 13 | 192 | 145 | 582 |
| Distance to | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Buildings | Forest Edge | Paved Roads | Streams | ||||||
| Years | Mean | 95% CI | Mean | 95% CI | Mean | 95% CI | Mean | 95% CI | |
| Species | Random points | 264 | 245, 285 | 125 | 115,135 | 89 | 81, 97 | 91 | 82, 100 |
| Felis catus | 2016 | 82 | 64, 104 | 229 | 186, 276 | 40 | 33, 47 | 119 | 84, 166 |
| 2017 | 77 | 54, 108 | 233 | 182, 293 | 33 | 22, 46 | 87 | 57, 128 | |
| 2018 | 73 | 49, 109 | 184 | 146, 227 | 44 | 32, 60 | 136 | 102, 180 | |
| 2019 | 93 | 67, 127 | 253 | 209, 301 | 30 | 21, 42 | 143 | 106, 191 | |
| Felis silvestris | 2016 | 297 | 232, 375 | 81 | 64, 100 | 60 | 48, 75 | 48 | 34, 67 |
| 2017 | 297 | 232, 376 | 56 | 47, 76 | 86 | 66, 110 | 54 | 41, 71 | |
| 2018 | 396 | 282, 543 | 65 | 42, 93 | 76 | 54, 106 | 47 | 31, 72 | |
| 2019 | 392 | 277, 572 | 92 | 43, 160 | 82 | 50, 125 | 73 | 43, 120 | |
| Vulpes vulpes | 2016 | 363 | 308, 425 | 110 | 89, 133 | 88 | 73, 106 | 77 | 59, 98 |
| 2017 | 421 | 357, 492 | 99 | 79, 121 | 116 | 100, 134 | 82 | 73, 104 | |
| 2018 | 210 | 141, 307 | 117 | 84, 155 | 63 | 48, 102 | 54 | 37, 76 | |
| 2019 | 319 | 240, 416 | 149 | 114, 190 | 92 | 73, 112 | 82 | 59, 113 | |
| 2016 | 2017 | |||||||
| Species | Mean ± SD | Range | N | p-Value | Mean ± SD | Range | N | p-Value |
| Fc-Fs | 932 ± 537 | 107–2861 | 63 | <0.001 | 1152 ± 472 | 57–1845 | 33 | <0.001 |
| Fc-Vv | 326 ± 216 | 27–1070 | 63 | 433 ± 235 | 83–936 | 33 | ||
| Fs-Fc | 1524 ± 1232 | 107–3736 | 51 | <0.001 | 859 ± 558 | 56–2262 | 53 | <0.001 |
| Fs-Vv | 182 ± 167 | 23–871 | 51 | 265 ± 328 | 9–1404 | 53 | ||
| Vv-Fc | 1248 ± 1128 | 10–3729 | 100 | <0.001 | 739 ± 387 | 83–1529 | 87 | 0.044 |
| Vv-Fs | 598 ± 692 | 21–3258 | 100 | 690 ± 645 | 8–2227 | 87 | ||
| 2018 | 2019 | |||||||
| Species | Mean ± SD | Range | N | p-Value | Mean ± SD | Range | N | p-Value |
| Fc-Fs | 1025 ± 698 | 223–3385 | 34 | 0.004 | 1126 ± 623 | 229–2925 | 46 | 0.067 |
| Fc-Vv | 572 ± 498 | 131–3105 | 34 | 866 ± 603 | 26–2042 | 46 | ||
| Fs-Fc | 1206 ± 613 | 223–2294 | 29 | <0.001 | 889 ± 527 | 229–2100 | 10 | 0.056 |
| Fs-Vv | 154 ± 147 | 27–796 | 29 | 533 ± 639 | 8–1627 | 10 | ||
| Vv-Fc | 1604 ± 979 | 131–2964 | 33 | <0.001 | 1450 ± 933 | 26–3083 | 39 | 0.008 |
| Vv-Fs | 703 ± 549 | 47–1778 | 33 | 913 ± 653 | 8–2512 | 39 | ||
| 2016 | 2017 | |||||
| Species Pairs | Observed Overlap (%) | Simulated Overlap (%) | Results | Observed Overlap (%) | Simulated Overlap (%) | Results |
| Felis catus-Felis silvestris | 0 | 38.7; 38.3–39.1 | Segregation | 0 | 31.9; 31.6–32.2 | Segregation |
| Felis catus-Vulpes vulpes | 2.3 | 34.2; 33.9–34.5 | Segregation | 0 | 25.5; 25.3–25.8 | Segregation |
| Felis silvestris-Vulpes vulpes | 4.8 | 39.4; 39.1–39.7 | Segregation | 5.4 | 34.2; 33.9–34.5 | Segregation |
| 2018 | 2019 | |||||
| Species Pairs | Observed Overlap (%) | Simulated Overlap (%) | Results | Observed Overlap (%) | Simulated Overlap (%) | Results |
| Felis catus-Felis silvestris | 0 | 37.1; 36.6–37.6 | Segregation | 0 | 20; 19.6–20.4 | Segregation |
| Felis catus-Vulpes vulpes | 0 | 32.9; 32.4–33.4 | Segregation | 0 | 33.6; 33.2–34 | Segregation |
| Felis silvestris-Vulpes vulpes | 1.8 | 32.6; 32.2–33 | Segregation | 6.7 | 19.4; 19–19.8 | Segregation |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez, A.; Urra, F.; Jubete, F.; Román, J.; Revilla, E.; Palomares, F. Spatial Segregation between Red Foxes (Vulpes vulpes), European Wildcats (Felis silvestris) and Domestic Cats (Felis catus) in Pastures in a Livestock Area of Northern Spain. Diversity 2020, 12, 268. https://doi.org/10.3390/d12070268
Rodríguez A, Urra F, Jubete F, Román J, Revilla E, Palomares F. Spatial Segregation between Red Foxes (Vulpes vulpes), European Wildcats (Felis silvestris) and Domestic Cats (Felis catus) in Pastures in a Livestock Area of Northern Spain. Diversity. 2020; 12(7):268. https://doi.org/10.3390/d12070268
Chicago/Turabian StyleRodríguez, Alberto, Fermín Urra, Fernando Jubete, Jacinto Román, Eloy Revilla, and Francisco Palomares. 2020. "Spatial Segregation between Red Foxes (Vulpes vulpes), European Wildcats (Felis silvestris) and Domestic Cats (Felis catus) in Pastures in a Livestock Area of Northern Spain" Diversity 12, no. 7: 268. https://doi.org/10.3390/d12070268
APA StyleRodríguez, A., Urra, F., Jubete, F., Román, J., Revilla, E., & Palomares, F. (2020). Spatial Segregation between Red Foxes (Vulpes vulpes), European Wildcats (Felis silvestris) and Domestic Cats (Felis catus) in Pastures in a Livestock Area of Northern Spain. Diversity, 12(7), 268. https://doi.org/10.3390/d12070268