Genetic Diversity of Local Greek and Bulgarian Grapevine (Vitis Vinifera L.) Varieties

 ,
, 
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling, DNA Extraction and Genotyping
2.2. Data Analysis
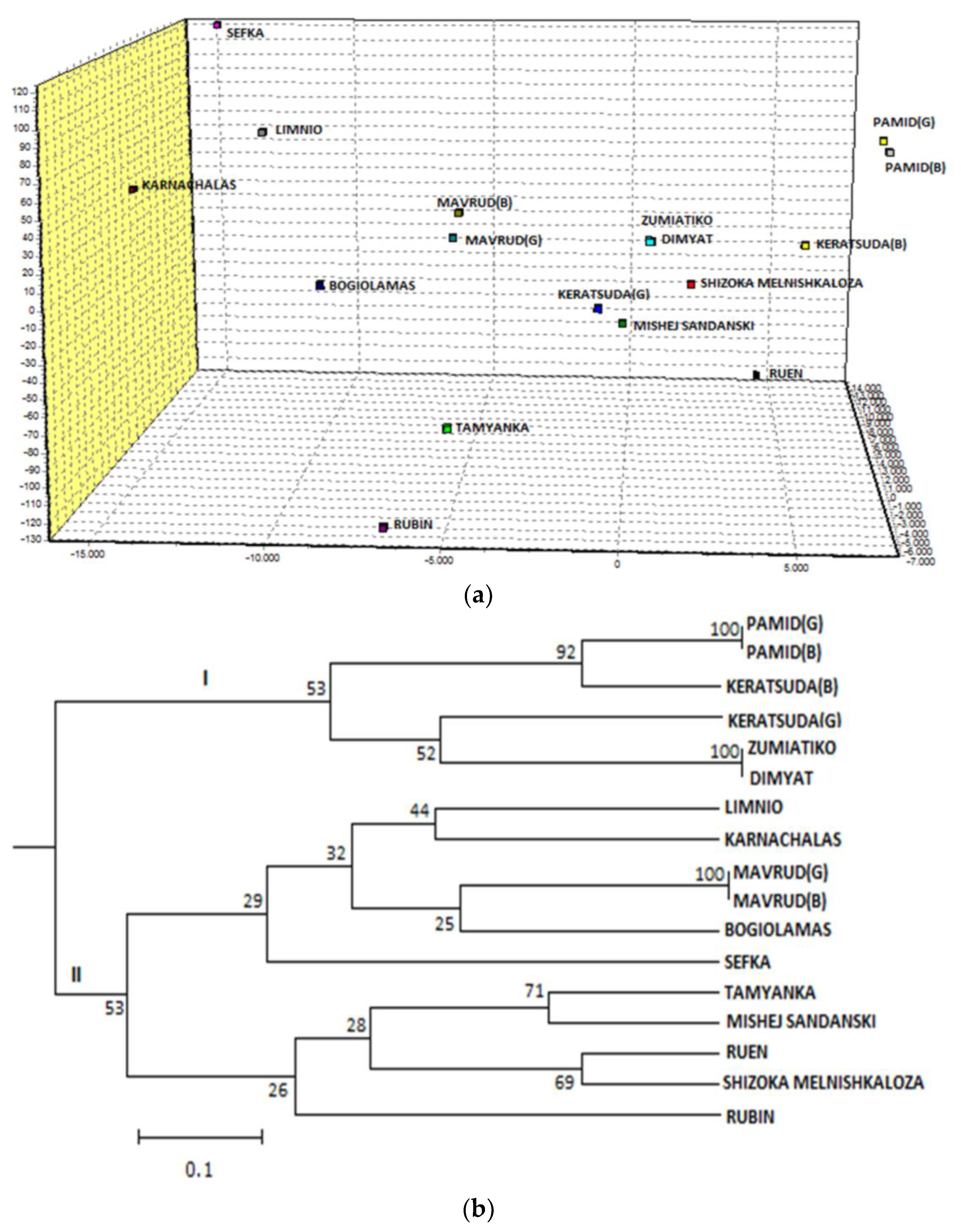
3. Results and Discussion
3.1. Genetic Variation
3.2. Genetic Differentiation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Olmo, H.P. Grapes. In Evolution of Crop Plants; Simmonds, N.W., Ed.; Longman: London, UK, 1976; pp. 294–298. [Google Scholar]
- Banilas, G.; Korkas, E.; Kaldis, P.; Hatzopoulos, P. Olive and grapevine biodiversity in Greece and Cyprus-a review. In Climate Change, Intercropping, Pest Control and Beneficial Microorganisms; Lichtfouse, E., Ed.; Springer: Berlin, Germany, 2009; pp. 401–428. [Google Scholar]
- Kotinis, X. Ampelographic Atlas of Greece (in Greek with French Summary). Athens. 1985. Available online: http://gvd.biology.uoc.gr/gvd/ (accessed on 16 March 2020).
- Hvarleva, T.; Rusanov, K.; Lefort, F.; Tsvetkov, I.; Atanassov, A.; Atanassov, I. Genotyping of Bulgarian Vitis vinifera L. cultivars by microsatellite analysis. Vitis 2004, 43, 27–34. [Google Scholar]
- This, P.; Lacombe, T.; Thomas, M.R. Historical origins and genetic diversity of wine grapes. Trends Genet. 2006, 22, 511–519. [Google Scholar] [CrossRef]
- Thomas, M.R.; Scott, N.S. Microsatellite repeats in grapevine reveal DNA polymorphisms when analysed as sequence-tagged sites (STSs). Theor. Appl. Genet. 1993, 86, 985–990. [Google Scholar] [CrossRef] [PubMed]
- Bowers, J.; Dangl, G.; Vignani, R.; Meredith, C. Isolation and characterization of new polymorphic simple sequence repeat loci in grape (Vitis vinifera L.). Genome 1996, 39, 628–633. [Google Scholar] [CrossRef] [PubMed]
- Bowers, J.; Dangl, G.; Meredith, C. Development and characterization of additional microsatellite DNA markers for grape. Am. J. Enol. Vitic. 1999, 50, 243–246. [Google Scholar]
- Laucou, V.; Boursiquot, J.M.; Lacombe, T.; Bordenave, L.; Decroocq, S.; Ollat, N. Parentage of grapevine rootstock ‘Fercal’ finally elucidated. Vitis 2008, 47, 163–167. [Google Scholar]
- Štajner, N.; Tomić, L.; Progar, V.; Pokorn, T.; Lacombe, T.; Laucou, V.; Boursiquot, J.M.; Javornik, B.; Bacilier, R. Genetic clustering and parentage analysis of Western Balkan grapevines (Vitis vinifera L.). Vitis 2015, 54, 67–72. [Google Scholar]
- Biagini, B.; Imazio, S.; Scienza, A.; Failla, O.; de Lorenzis, G. Renewal of wild grapevine (Vitis vinifera L. subsp. sylvestris (Gmelin) Hegi) populations through sexual pathway: Some Italian case studies. Flora 2016, 219, 85–93. [Google Scholar] [CrossRef]
- Lefort, F.; Roubelakis-Angelakis, K. Assessing the identity of grapevine plants from vineyards from Crete and Samos by microsatellite profiling. OENO One 2002, 36, 177–183. [Google Scholar] [CrossRef]
- Martin, J.P.; Borrego, J.; Cabello, F.; Ortiz, J.M. Characterization of Spanish grapevine cultivar diversity using sequence-tagged microsatellite site markers. Genome 2003, 46, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Vouillamoz, J.F.; McGovern, P.E.; Ergul, A.; Söylemezoglu, G.; Tevzadze, G.; Meredith, C.P.; Grando, M.S. Genetic characterization and relationships of traditional grape cultivars from Transcaucasia and Anatolia. Plant Genet. Resour. 2006, 4, 144–158. [Google Scholar] [CrossRef]
- De Lorenzis, G.; Imazio, S.; Rusjan, D.; Vouillamoz, J.F.; Nikolaou, N.; Failla, O.; Scienza, A. Genetic investigation of grapevine varieties ‘Ribolla Gialla’ (Italy), ‘Rebula’ (Slovenia) and ‘Robola’ (Ionian Islands). Sci. Hortic. 2013, 150, 425–431. [Google Scholar] [CrossRef]
- Žulj Mihalević, M.; Šimon, S.; Pejić, I.; Carka, F.; Sevo, R.; Kojic, A.; Gaši, F.; Tomić, L.; Jovanović Cvetković, T.; Maletić, E.; et al. Molecular characterization of old local grapevine varieties from South East European countries. Vitis 2013, 52, 69–76. [Google Scholar]
- Merkouropoulos, G.; Michailidou, S.; Alifragkis, A.; Zioziou, E.; Koundouras, S.; Argiriou, A.; Nikolaou, N. A combined approach involving ampelographic description, berry oenological traits and molecular analysis to study native grapevine varieties of Greece. Vitis 2015, 54, 99–103. [Google Scholar]
- Dong, Z.; Liu, W.; Li, X.; Tan, W.; Zhao, Q.; Wang, M.; Ren, R.; Ma, X.; Tang, X. Genetic relationships of 34 grapevine varieties and construction of molecular fingerprints by SSR markers. Biotechnol. Biotechnol. Equip. 2018, 32, 942–950. [Google Scholar] [CrossRef] [Green Version]
- Popescu, C.F.; Crespan, M. Combining Microsatellite Markers and Ampelography for Better Management of Romanian Grapevine Germplasm Collections. Not. Sci. Biol. 2018, 10, 193–198. [Google Scholar] [CrossRef]
- Mahmood, A.M.; Shaim, H.A.; Shabaq, M.H. Molecular characterization of autochthonous grapevine (Vitis vinifera L.) cultivars in Kurdistan using simple sequence repeats (SSRs). J. Univ. Garmian 2019, 6, 328–335. [Google Scholar] [CrossRef]
- Taheri, F.; DarziRamandi, H. Microsatellite Markers Analysis for the Genetic Characterization and Relationships among Some of Iranian Local Grapevine Accessions (Vitis vinifera L.). Int. J. Fruit Sci. 2020, 20, 1–18. [Google Scholar] [CrossRef]
- The Cooperation Programme INTERREG V-A “Greece-Bulgaria 2014–2020”. Available online: http://www.greece-bulgaria.eu/com/4_A-few-words-about-our-Programme (accessed on 13 April 2020).
- Toonen, R.J.; Hughes, S. Increased throughput for fragment analysis on an ABI Prism377 automated sequencer using a membrane comb and STRand software. Biotechniques 2001, 31, 1320–1324. [Google Scholar]
- Drábek, J.; Smolíková, M.; Kalendar, R.A.; Lopes Pinto, F.; Pavlouŝek, P.; Klepárník, K.; Frébort, I. Design and validation of an STR hexaplex assay for DNA profiling of grapevine cultivars. Electrophoresis 2016, 37, 3059–3067. [Google Scholar] [CrossRef] [Green Version]
- Sefc, K.; Regner, F.; Glössl, J.; Steinkellner, H. Identification of microsatellite sequences in Vitis riparia and their applicability for genotyping of different Vitis species. Genome 1999, 42, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Weir, B.S.; Cockerham, C.C. Estimating F-statistics for the analysis of population structure. Evolution 1984, 37, 1358–1370. [Google Scholar]
- Goudet, J. FSTAT (Version 1.2): A computer program to calculate F-statistics. J. Hered. 1995, 86, 485–486. [Google Scholar] [CrossRef]
- Belkhir, K.; Borsa, P.; Chikhi, L.; Raufaste, N.; Bonhomme, F. GENETIX 4.05, Logiciel Sous Windows TM Pour la Génétique des Populations. Laboratoire Genome; Populations, Interactions, CNRS UMR 5000, Universite’ de Montpellier II: Montpellier, France, 2004. [Google Scholar]
- Takezaki, N.; Nei, M.; Tamura, K. POPTREEW: Web version of POPTREE for constructing population trees from allele frequency data and computing some other quantities. Mol. Biol. Evol. 2004, 31, 1622–1624. [Google Scholar] [CrossRef] [Green Version]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar]
- Sefc, K.M.; Lopes, M.S.; Lefort, F.; Botta, R.; Roubelakis-Angelakis, K.A.; Ibáñez, J.; Peji’c, I.; Wagner, H.W.; Glössl, J.; Steinkellner, H. Microsatellite variability in grapevine cultivars from different European regions and evaluation of assignment testing to assess the geographic origin of cultivars. Theor. Appl. Genet. 2000, 100, 498–505. [Google Scholar] [CrossRef]
- Biniari, K.; Stavrakaki, M. Genetic study of native grapevine varieties of northern, western and central Greece with the use of ampelographic and molecular methods. Not. Bot. Horti. Agrobo. 2019, 47, 46–53. [Google Scholar] [CrossRef] [Green Version]
- Stavrakaki, M.; Biniari, K. Ampelographic and genetic characterization of grapevine varieties (Vitis vinifera L.) of the ‘Mavroudia’ group cultivated in Greece. Not. Bot. Horti. Agrobo. 2017, 45, 525–531. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Variety | A | Ho | He | Fis |
|---|---|---|---|---|
| Pamid(G) (also known as ‘Pamidi’) | 3.0000 | 0.7347 | 0.4252 | −0.7780 * |
| Keratsouda(G) | 1.5714 | 0.5714 | 0.3008 | −0.6915 * |
| Zoumiatiko | 1.8571 | 0.8571 | 0.4511 | −0.9080 * |
| Limnio | 1.8571 | 0.8571 | 0.4511 | −0.9080 * |
| Sefka | 1.8571 | 0.8571 | 0.4511 | −0.9080 * |
| Mavrud(G) (also known as ‘Mavroudi’) | 2.1429 | 1.0000 | 0.5481 | −0.9440 * |
| Bogialamas | 2.0000 | 0.7143 | 0.3931 | −0.7638 * |
| Karnachalas | 2.0000 | 1.0000 | 0.5238 | −0.9775 * |
| Τamyanka | 4.5714 | 0.8736 | 0.5930 | −0.4880 * |
| Mavrud(B) | 4.1429 | 0.9725 | 0.5952 | −0.6550 * |
| Rubin | 2.2857 | 0.8500 | 0.4533 | −0.9190 * |
| Pamid(B) | 2.8571 | 0.7143 | 0.4726 | −0.5690 * |
| Dimyat | 1.8571 | 0.8571 | 0.4403 | −0.9325 * |
| Ruen | 1.8571 | 0.7214 | 0.3734 | −0.9800 * |
| Shizoka Melnishkaloza | 2.7143 | 0.5807 | 0.3258 | −0.8020 * |
| Mishej Sandanski | 4.7143 | 0.6554 | 0.4835 | −0.3650 * |
| Keratsouda(B) | 4.5714 | 0.5946 | 0.3580 | −0.6660 * |
| Mean | 2.6975 | 0.7889 | 0.4494 | −0.7797 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papapetrou, M.; Loukovitis, D.; Papadopoulos, O.; Kazlari, Z.; Peristeraki, A.; Arsenova, S.; Bardarova, D.; Doncheva, D.; Theocharis, S.; Karagiannidis, C.; et al. Genetic Diversity of Local Greek and Bulgarian Grapevine (Vitis Vinifera L.) Varieties. Diversity 2020, 12, 273. https://doi.org/10.3390/d12070273
Papapetrou M, Loukovitis D, Papadopoulos O, Kazlari Z, Peristeraki A, Arsenova S, Bardarova D, Doncheva D, Theocharis S, Karagiannidis C, et al. Genetic Diversity of Local Greek and Bulgarian Grapevine (Vitis Vinifera L.) Varieties. Diversity. 2020; 12(7):273. https://doi.org/10.3390/d12070273
Chicago/Turabian StylePapapetrou, Maria, Dimitrios Loukovitis, Orestis Papadopoulos, Zoi Kazlari, Anastasia Peristeraki, Slavina Arsenova, Desislava Bardarova, Desislava Doncheva, Serafeim Theocharis, Constantinos Karagiannidis, and et al. 2020. "Genetic Diversity of Local Greek and Bulgarian Grapevine (Vitis Vinifera L.) Varieties" Diversity 12, no. 7: 273. https://doi.org/10.3390/d12070273
APA StylePapapetrou, M., Loukovitis, D., Papadopoulos, O., Kazlari, Z., Peristeraki, A., Arsenova, S., Bardarova, D., Doncheva, D., Theocharis, S., Karagiannidis, C., Koundouras, S., Giannakoula, A., Aggelopoulos, S., & Chatziplis, D. (2020). Genetic Diversity of Local Greek and Bulgarian Grapevine (Vitis Vinifera L.) Varieties. Diversity, 12(7), 273. https://doi.org/10.3390/d12070273