Spatial and Temporal Variation in Deep-Sea Meiofauna at the LTER Observatory HAUSGARTEN in the Fram Strait (Arctic Ocean)


Abstract
:1. Introduction
2. Materials and Methods
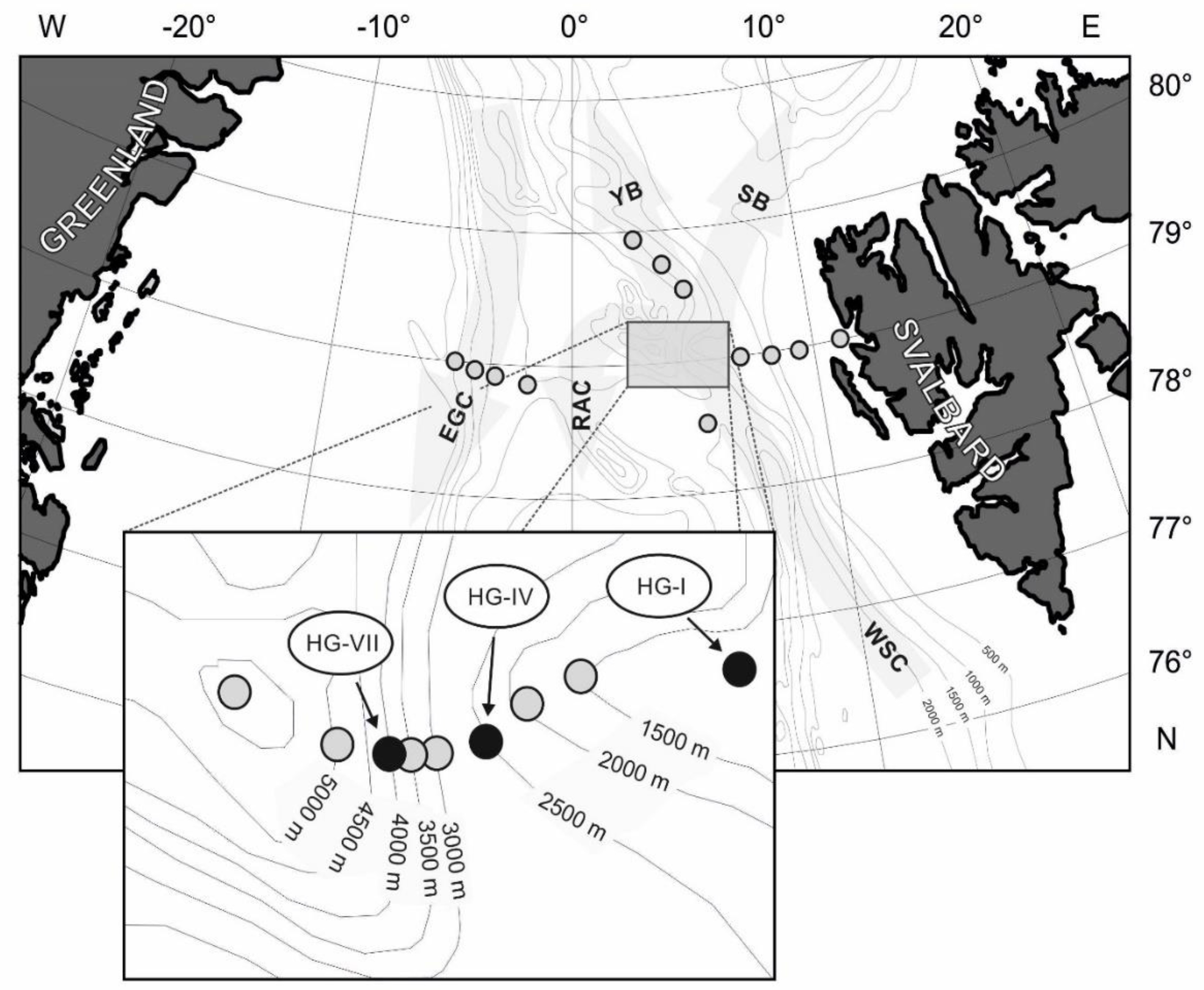
2.1. Study Site
2.2. Annual Sediment Sampling
2.3. Sample Processing for Meiofauna Organisms
2.4. Assessment of Environmental Background Data
2.5. Statistical Analyses
3. Results
3.1. Meiofaunal Data
3.2. Environmental Data
3.3. Relationships between Meiofaunal and Environmental Data
4. Discussion
4.1. Spatial and Temporal Patterns in Deep-Sea Meiofauna Communities at HAUSGARTEN
4.2. Sources of Variation in Deep-Sea Meiofauna Communities at HAUSGARTEN
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schratzberger, M.; Ingels, J. Meiofauna matters: The roles of meiofauna in benthic ecosystems. J. Exp. Mar. Biol. Ecol. 2017, 502, 12–25. [Google Scholar] [CrossRef]
- Coull, B.C. Are members of the meiofauna food for higher trophic levels? Trans. Am. Microsc. Soc. 1990, 109, 233–246. [Google Scholar] [CrossRef]
- Nehring, S. Tube-dwelling Meiofauna in Marine Sediments. Int. Rev. ges. Hydrobiol. Hydrogr. 1993, 78, 521–534. [Google Scholar] [CrossRef]
- Meysman, F.J.; Middelburg, J.J.; Heip, C.H. Bioturbation: A fresh look at Darwin’s last idea. Trends Ecol. Evol. 2006, 21, 688–695. [Google Scholar] [CrossRef]
- Näslund, J.; Nascimento, F.J.; Gunnarsson, J.S. Meiofauna reduces bacterial mineralization of naphthalene in marine sediment. ISME J. 2010, 4, 1421–1430. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, N.L.; Aarnio, K.; Mäensivu, M.; Bonsdorff, E. Drifting filamentous algal mats disturb sediment fauna: Impacts on macro-meiofaunal interactions. J. Exp. Mar. Biol. Ecol. 2012, 420, 77–90. [Google Scholar] [CrossRef]
- Thiel, H. Meiobenthos and nanobenthos of the deep-sea. In Deep-Sea Biology, The Sea; Rowe, G.T., Ed.; John Wiley and Sons: New York, NY, USA, 1983; Volume 8, pp. 167–230. [Google Scholar]
- Tietjen, J.H. Abundance and biomass of metazoan meiobenthos in the deep-sea. In Deep-Sea Food Chains and the Global Carbon Cycle; Rowe, G.T., Pariente, V., Eds.; Kluwer Academic Publ.: Dordrecht, The Netherlands, 1992; Volume 360, pp. 45–62. [Google Scholar]
- Soltwedel, T. Metazoan meiobenthos along continental margins: A review. Prog. Oceanogr. 2000, 46, 59–84. [Google Scholar] [CrossRef]
- Rosli, N.; Leduc, D.; Rowden, A.A.; Probert, P.K. Review of recent trends in ecological studies of deep-sea meiofauna, with focus on patterns and processes at small to regional spatial scales. Mar. Biodiv. 2018, 48, 13–34. [Google Scholar] [CrossRef]
- Neira, C.; Sellanes, J.; Levin, L.A.; Arntz, W.E. Meiofaunal distributions on the Peru margin: Relationship to oxygen and organic matter availability. Deep Sea Res. I 2001, 48, 2453–2472. [Google Scholar]
- Van Gaever, S.; Vanreusel, A.; Hughes, J.A.; Bett, B.; Kiriakoulakis, K. The macro- and micro-scale patchiness of meiobenthos associated with the Darwin Mounds (north-east Atlantic). J. Mar. Biol. Assoc. UK 2004, 84, 547–556. [Google Scholar] [CrossRef]
- Vanreusel, A.; Fonseca, G.; Danovaro, R.; Da Silva, M.C.; Esteves, A.M.; Ferrero, T.; Gad, G.; Galtsova, V.; Gambi, C.; Genevois, V.D.F.; et al. The contribution of deep-sea macrohabitat heterogeneity to global nematode diversity. Mar. Ecol. 2010, 31, 6–20. [Google Scholar] [CrossRef] [Green Version]
- Pusceddu, A.; Bianchelli, S.; Martin, J.; Puig, P.; Palanques, A.; Masque, P.; Danovaro, R. Chronic and intensive bottom trawling impairs deep-sea biodiversity and ecosystem functioning. Proc. Natl. Acad. Sci. USA 2014, 111, 8861–8866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soltwedel, T.; Bauerfeind, E.; Bergmann, M.; Budaeva, N.; Hoste, E.; Jaeckisch, N.; Juterzenka, K.V.; Matthießen, J.; Mokievsky, V.; Nöthig, E.-M.; et al. HAUSGARTEN: Multidisciplinary investigations at a deep-sea, long-term observatory in the Arctic Ocean. Oceanography 2005, 18, 46–61. [Google Scholar] [CrossRef] [Green Version]
- Jacob, M.; Soltwedel, T.; Boetius, A.; Ramette, A. Biogeography of deep-sea benthic bacteria at regional scale (LTER HAUSGARTEN, Fram Strait, Arctic). PLoS ONE 2013, 8, e72779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacob, M. Influence of Global Change on Microbial Communities in Arctic Sediments. Ph.D. Thesis, University of Bremen, Bremen, Germany, 2014; 178p. [Google Scholar]
- Soltwedel, T.; Jaeckisch, N.; Ritter, N.; Hasemann, C.; Bergmann, M.; Klages, M. Bathymetric patterns of megafaunal assemblages from the arctic deep-sea observatory HAUSGARTEN. Deep Sea Res. I 2009, 56, 1856–1872. [Google Scholar] [CrossRef]
- Bergmann, M.; Soltwedel, T.; Klages, M. The interannual variability of megafaunal assemblages in the Arctic deep sea: Preliminary results from the HAUSGARTEN observatory (79°N). Deep Sea Res. I 2011, 58, 711–723. [Google Scholar] [CrossRef]
- Taylor, J.; Krumpen, T.; Soltwedel, T.; Gutt, J.; Bergmann, M. Dynamic benthic communities: Assessing temporal variations in benthic community structure, megafaunal composition and diversity at the Arctic deep sea observatory HAUSGARTEN between 2004 and 2015. Deep Sea Res. I 2017, 122, 81–94. [Google Scholar] [CrossRef] [Green Version]
- Taylor, J.; Staufenbiel, B.; Soltwedel, T.; Bergmann, M. Temporal trends in the biomass of three epibenthic invertebrates from the deep-sea observatory HAUSGARTEN (Fram Strait, Arctic Ocean). Mar. Ecol. Prog. Ser. 2018, 602, 15–29. [Google Scholar] [CrossRef]
- Hoste, E. Temporal and Spatial Variability in Deep-Sea Meiobenthic Communities from the Arctic Marginal Ice Zone. Ph.D. Thesis, Ghent University, Ghent, Belgium, 2015; 185p. [Google Scholar]
- Grzelak, K. Structural and Functional Diversity of Nematoda at the Arctic Deep-Sea Long-Term Observatory Hausgarten (Fram Strait). Ph.D. Thesis, Institute of Oceanology Polish Academy of Sciences, Sopot, Poland, 2015; 206p. [Google Scholar]
- Soltwedel, T.; Bauerfeind, E.; Bergmann, M.; Bracher, A.; Budaeva, N.; Busch, K.; Cherkasheva, A.; Fahl, K.; Grzelak, K.; Hasemann, C.; et al. Natural variability or anthropogenically-induced variation? Insights from 15 years of multidisciplinary observations at the arctic marine LTER site HAUSGARTEN. Ecol. Ind. 2016, 65, 89–102. [Google Scholar] [CrossRef]
- Beszczynska-Möller, A.; Fahrbach, E.; Schauer, U.; Hansen, E. Variability in Atlantic water temperature and transport at the entrance to the Arctic Ocean, 1997–2010. ICES J. Mar. Sci. 2012, 69, 852–863. [Google Scholar] [CrossRef] [Green Version]
- De Steur, L.; Hansen, E.; Gerdes, R.; Karcher, M.; Fahrbach, E.; Holfort, J. Freshwater fluxes in the East Greenland Current: A decade of observations. Geophys. Res. Lett. 2009, 36, L23611. [Google Scholar] [CrossRef] [Green Version]
- Paquette, R.G.; Bourke, R.H.; Newton, J.F.; Perdue, W.F. The East Greenland Polar Front in autumn. J. Geophys. Res. 2012, 90, 4866–4882. [Google Scholar] [CrossRef]
- Pfannkuche, O.; Thiel, H. Sample Processing. In Introduction to the Study of Meiofauna; Higgens, R.P., Thiel., H., Eds.; Smithsonian Institute Press: Washington, DC, USA; London, UK, 1988; pp. 134–145. [Google Scholar]
- Buchanan, J.B. Sediment analysis. In Methods for the Study of Marine Benthos; Holme, N.A., McIntyre, A.D., Eds.; Blackwell Scientific Publications: Oxford, UK, 1984; pp. 41–65. [Google Scholar]
- Hebbeln, D.; Wefer, G. Late Quaternary paleoceanography in the Fram Strait. Paleoceanography 1997, 12, 65–78. [Google Scholar] [CrossRef]
- Soltwedel, T.; Hasemann, C.; Vedenin, A.; Bergmann, M.; Taylor, J.; Krauß, F. Bioturbation rates in the deep Fram Strait: Results from long-term in situ experiments at the arctic LTER Observatory HAUSGARTEN. J. Exp. Mar. Biol. Ecol. 2019, 511, 1–9. [Google Scholar] [CrossRef]
- Greiser, N.; Faubel, A. Biotic factors. In Introduction to the Study of Meiofauna; Higgens, R.P., Thiel, H., Eds.; Smithsonian Institution Press: Washington, DC, USA; London, UK, 1988; pp. 79–114. [Google Scholar]
- Shuman, F.R.; Lorenzen, C.F. Quantitative degradation of chlorophyll by a marine herbivore. Limnol. Oceanogr. 1975, 20, 580–586. [Google Scholar] [CrossRef] [Green Version]
- Thiel, H. Benthos in upwelling regions. In Upwelling Ecosystems; Boje, R., Tomczak, M., Eds.; Springer Verlag: Berlin/Heidelberg, Germany, 1978; pp. 124–138. [Google Scholar]
- Findlay, R.H.; King, G.M.; Watling, L. Efficiency of phospholipid analysis in determining microbial biomass in sediments. Appl. Environ. Microbiol. 1989, 55, 2888–2893. [Google Scholar] [CrossRef] [Green Version]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 2nd ed.; PRIMER-E Ltd.: Dhaka, Bangladesh; Plymouth Marine Laboratory: Plymouth, UK, 2001. [Google Scholar]
- Clarke, K.; Somerfield, P.; Gorley, R. Testing of null hypotheses in exploratory community analyses: Similarity profiles and biota-environment linkage. J. Exp. Mar. Biol. Ecol. 2008, 366, 56–69. [Google Scholar] [CrossRef]
- McArdle, B.H.; Anderson, M.J. Fitting Multivariate Models to Community Data: A Comment on Distance-Based Redundancy Analysis. Ecology 2001, 82, 290–297. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008; 214p. [Google Scholar]
- Borcard, D.; Legendre, P.; Drapeau, P. Partialling out the spatial component of ecological variation. Ecology 1992, 73, 1045–1055. [Google Scholar] [CrossRef] [Green Version]
- Leduc, D.; Rowden, A.A.; Bowden, D.A.; Nodder, S.D.; Probert, P.K.; Pilditch, C.A.; Duineveld, G.C.A.; Witbaard, R. Nematode beta diversity on the continental slope of New Zealand: Spatial patterns and environmental drivers. Mar. Ecol. Prog. Ser. 2012, 454, 37–52. [Google Scholar] [CrossRef] [Green Version]
- Hoste, E.; Vanhove, S.; Schewe, I.; Soltwedel, T.; Vanreusel, A. Spatial and temporal variations in deep-sea meiofauna assemblages in the Marginal Ice Zone of the Arctic Ocean. Deep Sea Res. I 2007, 54, 109–129. [Google Scholar] [CrossRef]
- Gorska, B.; Grzelak, K.; Kotwicki, L.; Hasemann, C.; Schewe, I.; Soltwedel, T.; Wlodarska Kowalczuk, M. Bathymetric variations in vertical distribution patterns of meiofauna in the surface sediments of the deep Arctic Ocean (HAUSGARTEN, Fram Strait). Deep Sea Res. I 2014, 91, 36–49. [Google Scholar] [CrossRef]
- Grzelak, K.; Kotwicki, L.; Hasemann, C.; Soltwedel, T. Bathymetric patterns in standing stock and diversity of deep-sea nematodes at the long-term ecological research observatory HAUSGARTEN (Fram Strait). J. Mar. Sys. 2017, 172, 160–177. [Google Scholar] [CrossRef]
- Kalogeropoulou, V.; Bett, B.J.; Gooday, A.J.; Lampadariou, N.; Martinez Arbizu, P.; Vanreusel, A. Temporal changes (1989–1999) in deep-sea metazoan meiofaunal assemblages on the Porcupine Abyssal Plain, NE Atlantic. Deep Sea Res. II 2010, 57, 1383–1395. [Google Scholar] [CrossRef] [Green Version]
- Käß, M.; Vedenin, A.; Hasemann, C.; Brandt, A.; Soltwedel, T. Community structure of macrofauna in the deep Fram Strait: A comparison between two bathymetric gradients in ice covered and ice-free areas. Deep Sea Res. I 2019, 152, 103102. [Google Scholar] [CrossRef]
- Smith, C.R.; DeMaster, D.J.; Thomas, C.; Sršen, P.; Grange, L.; Evrard, V.; DeLeo, F. Pelagic-benthic coupling, food banks, and climate change on the West Antarctic Peninsula Shelf. Oceanography 2012, 25, 188–201. [Google Scholar] [CrossRef] [Green Version]
- Larsen, K.M.H.; Gonzalez-Pola, C.; Fratantoni, P.; Beszczynska-Möller, A.; Hughes, S.L. (Eds.) ICES Report on Ocean Climate 2015. ICES Coop. Res. Rep. 2016, 331, 79. [Google Scholar]
- Nöthig, E.-M.; Bracher, A.; Engel, A.; Metfies, K.; Niehoff, B.; Peeken, I.; Bauerfeind, E.; Cherkasheva, A.; Gäbler-Schwarz, S.; Hardge, K.; et al. Summertime plankton ecology in Fram Strait—A compilation of long- and short-term observations. Polar Res. 2015, 34, 23349. [Google Scholar] [CrossRef]
- Kraft, A.; Bauerfeind, E.; Nöthig, E.-M.; Bathmann, U. Size structure and lifecycle patterns of dominant pelagic amphipods collected as swimmers in sediment traps in the eastern Fram Strait. J. Mar. Syst. 2012, 95, 1–15. [Google Scholar] [CrossRef]
- Kraft, A.; Nöthig, E.-M.; Bauerfeind, E.; Wildish, D.J.; Pohle, G.W.; Bathmann, U.; Beszczynska-Möller, A.; Klages, M. First evidence of reproductive success in a southern invader indicates possible community shifts among Arctic zooplankton. Mar. Ecol. Prog. Ser. 2013, 493, 291–296. [Google Scholar] [CrossRef] [Green Version]
- Bauerfeind, E.; Nöthig, E.-M.; Pauls, B.; Kraft, A.; Beszczynska-Möller, A. Variability in pteropod sedimentation and corresponding aragonite flux at the Arctic deep-sea long-term observatory HAUSGARTEN in the eastern Fram Strait from 2000 to 2009. J. Mar. Syst. 2014, 132, 95–105. [Google Scholar] [CrossRef]
- Weydmann, A.; Carstensen, J.; Goszczko, I.; Dmoch, K.; Olszewska, A.; Kwasniewski, S. Shift towards the dominance of boreal species in the Arctic: Inter-annual and spatial zooplankton variability in the West Spitsbergen Current. Mar. Ecol. Prog. Ser. 2014, 501, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Busch, K.; Bauerfeind, E.; Nöthig, E.-M. Pteropod sedimentation patterns in different water depths observed with moored sediment traps over a 4-year period at the LTER station HAUSGARTEN in eastern Fram Strait. Polar Biol. 2015, 38, 845–859. [Google Scholar] [CrossRef]
- Saiz, E.; Calbet, A.; Isari, S.; Anto, M.; Velasco, E.M.; Almeda, R.; Movilla, J.; Alcaraz, M. Zooplankton distribution and feeding in the Arctic Ocean during a Phaeocystis pouchetii bloom. Deep Sea Res. I 2013, 72, 17–33. [Google Scholar] [CrossRef] [Green Version]
- Veit-Köhler, G.; Guilini, K.; Peeken, I.; Quillfeldt, P.; Mayr, C. Carbon and nitrogen stable isotope signatures of deep-sea meiofauna follow oceanographical gradients across the Southern Ocean. Prog. Oceanogr. 2013, 110, 69–79. [Google Scholar] [CrossRef]
- Vanhove, S.; Beghyn, M.; Van Gansbeke, D.; Bullough, L.W.; Vincx, M. A seasonally varying biotope at Signy Island, Antarctic: Implications for meiofaunal structure. Mar. Ecol. Prog. Ser. 2000, 202, 13–25. [Google Scholar] [CrossRef]
- Zeppilli, D.; Leduc, D.; Fernandes, D. Characteristics of meiofauna in extreme marine ecosystems: A review. Mar. Biodiv. 2018, 48, 35–71. [Google Scholar] [CrossRef] [Green Version]
- Morrisey, D.J.; Howitt, L.; Underwood, A.J.; Stark, J.S. Spatial variation in soft-sediment benthos. Mar. Ecol. Prog. Ser. 1992, 81, 197–204. [Google Scholar] [CrossRef]
- Underwood, A.J. Spatial patterns of variance in densities of intertidal populations. In Frontiers of Population Ecology; Floyd, R.B., Sheppard, A.W., De Barro, P.J., Eds.; CSIRO Publishing: Melbourne, Australia, 1996; pp. 369–389. [Google Scholar]
- Underwood, A.J.; Chapman, M.G.; Connell, S.D. Observations in ecology: You can’t make progress on processes without understanding the patterns. J. Exp. Mar. Biol. Ecol. 2000, 250, 97–115. [Google Scholar] [CrossRef]
- Gooday, A.J.; Pfannkuche, O.; Lambshead, P.J.D. An apparent lack of response by metazoan meiofauna to phytodetritus deposition in the bathyal north-eastern Atlantic. J. Mar. Biol. Assoc. UK 1996, 76, 297–310. [Google Scholar] [CrossRef]
- Soetaert, K.; Vanaverbeke, J.; Heip, C.; Herman, P.M.J.; Middelburg, J.J.; Duineveld, G.; Sandee, A. Nematode distribution in ocean margin sediments of the Goban Spur (north-east Atlantic) in relation to sediment geochemistry. Deep Sea Res. I 1997, 44, 1671–1683. [Google Scholar] [CrossRef]
- Ingels, J.; Billett, D.S.M.; Kiriakoulakis, K.; Wolff, G.A.; Vanreusel, A. Structural and functional diversity of Nematoda in relation with environmental variables in the Setúbal and Cascais canyons, Western Iberian Margin. Deep Sea Res. II 2011, 58, 2354–2368. [Google Scholar] [CrossRef]
- Ingels, J.; Tchesunov, A.V.; Vanreusel, A. Meiofauna in the Gollum Channels and the Whittard Canyon, Celtic Margin—How local environmental conditions shape nematode structure and function. PLoS ONE 2011, 6, e20094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeppilli, D.; Pusceddu, A.; Trincardi, F.; Danovaro, R. Seafloor heterogeneity influences the biodiversity-ecosystem functioning relationships in the deep sea. Sci. Rep. 2016, 6, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, C.; Martínez Arbizu, P. Unexpectedly higher metazoan meiofauna abundances in the Kuril–Kamchatka Trench compared to the adjacent abyssal plains. Deep Sea Res. II 2015, 111, 60–75. [Google Scholar] [CrossRef]
- Vanreusel, A.; Vincx, M.; Schramm, D.; Van Gansbeke, D. On the vertical distribution of the metazoan meiofauna in shelf break and upper slope habitats of the NE Atlantic. Int. Rev. Ges. Hydrobiol. 1995, 80, 313–326. [Google Scholar] [CrossRef]
- Vanaverbeke, J.; Soetaert, K.; Heip, C.; Vanreusel, A. The meiobenthos along the continental slope of the Goban Spur (NE Atlantic). J. Sea. Res. 1997, 38, 93–108. [Google Scholar] [CrossRef]
- Giere, O. Meiobenthology. The Microscopic Motile Fauna of Aquatic Sediments, 2nd ed.; Springer-Verlag: Berlin/Heidelberg, Germany, 2009; 527p. [Google Scholar]
- Moens, T.; Braeckman, U.; Derycke, S.; Fonseca, G.; Gallucci, F.; Ingels, J.; Leduc, D.; Vanaverbeke, J.; van Colen, C.; Vanreusel, A.; et al. Ecology of free-living marine nematodes. In Handbook of Zoology: Gastrotricha, Cycloneuralia and Gnathifera, Vol. 2: Nematoda; Schmidt-Rhaesa, A., Ed.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2014; pp. 109–152. [Google Scholar]
- Romano, C.; Coenjaerts, J.; Flexas, M.M.; Zuniga, D.; Vanreusel, A.; Company, J.B.; Martin, D. Spatial and temporal variability of meiobenthic density in the Blanes submarine canyon (NW Mediterranean). Prog. Oceanogr. 2013, 118, 144–158. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Cruise | Station-ID | Site | Lat N | Long E | Depth (m) |
|---|---|---|---|---|---|---|
| 19 August 2000 | ARK-XVI/2 | PS57/272 | HG-I | 79°08.28′ | 06°06.19′ | 1246 |
| 05 August 2000 | ARK-XVI/2 | PS57/178 | HG-IV | 79°04.10′ | 04°11.20′ | 2385 |
| 06 August 2000 | ARK-XVI/2 | PS57/183 | HG-VII | 79°03.60′ | 03°28.80′ | 4020 |
| 12 July 2001 | ARK-XVII | PS59/091 | HG-I | 79°08.00′ | 06°04.50′ | 1284 |
| 13 July 2001 | ARK-XVII | PS59/094 | HG-IV | 79°04.00′ | 04°10.40′ | 2468 |
| 15 July 2001 | ARK-XVII | PS59/108 | HG-VII | 79°04.00′ | 03°29.20′ | 3997 |
| 06 August 2002 | ARK-XVIII/1b | PS62/171 | HG-I | 79°08.44′ | 06°05.49′ | 1292 |
| 02 August 2002 | ARK-XVIII/1b | PS62/161 | HG-IV | 79°03.90′ | 04°10.93′ | 2469 |
| 08 August 2002 | ARK-XVIII/1b | PS62/183 | HG-VII | 79°03.60′ | 03°28.87′ | 4039 |
| 21 July 2003 | ARK-XIX/3c | PS64/402-1 | HG-I | 79°08.00′ | 06°05.54′ | 1277 |
| 26 July 2003 | ARK-XIX/3c | PS64/429-1 | HG-IV | 79°04.31′ | 04°07.57′ | 2501 |
| 02 August 2003 | ARK-XIX/3c | PS64/464-1 | HG-VII | 79°03.57′ | 03°28.49′ | 4098 |
| 07 July 2004 | ARK-XX/1 | PS66/104-1 | HG-I | 79°07.99′ | 06°05.46′ | 1281 |
| 09 July 2004 | ARK-XX/1 | PS66/117-1 | HG-IV | 79°05.00′ | 04°04.98′ | 2508 |
| 10 July 2004 | ARK-XX/1 | PS66/122-2 | HG-VII | 79°03.56′ | 03°28.57′ | 4090 |
| 26 August 2005 | ARK-XXI/1b | PS68/277-2 | HG-I | 79°08.00′ | 06°05.57′ | 1279 |
| 19 August 2005 | ARK-XXI/1b | PS68/238-3 | HG-IV | 79°03.91′ | 04°10.81′ | 2462 |
| 24 August 2005 | ARK-XXI/1b | PS68/267-2 | HG-VII | 79°03.61′ | 03°28.55′ | 4008 |
| 22 August 2006 | MSM02-4 | MSM2/773-1 | HG-I | 79°07.99′ | 06°05.49′ | 1266 |
| 24 August 2006 | MSM02-4 | MSM2/780-4 | HG-IV | 79°03.93′ | 04°10.84′ | 2411 |
| 06 September 2006 | MSM02-4 | MSM2/877-1 | HG-VII | 79°03.91′ | 03°29.56′ | 3923 |
| 12 July 2007 | ARK-XXII/1c | PS70/163-1 | HG-I | 79°08.07′ | 05°59.45′ | 1304 |
| 10 July 2007 | ARK-XXII/1c | PS70/147-1 | HG-IV | 79°03.92′ | 04°10.55′ | 2477 |
| 19 July 2007 | ARK-XXII/1c | PS70/211-1 | HG-VII | 79°03.59′ | 03°28.50′ | 4065 |
| 12 July 2008 | ARK-XXVIII/1b | PS72/137-2 | HG-I | 79°08.00′ | 06°05.51′ | 1287 |
| 09 July 2008 | ARK-XXVIII/1b | PS72/122-2 | HG-IV | 79°03.92′ | 04°11.01′ | 2417 |
| 17 July 2008 | ARK-XXVIII/1b | PS72/160-1 | HG-VII | 79°03.50′ | 03°28.83′ | 4070 |
| 13 July 2009 | ARK-XXIV/2 | PS74/109-2 | HG-I | 79°08.07′ | 06°05.79′ | 1285 |
| 16 July 2009 | ARK-XXIV/2 | PS74/121-1 | HG-IV | 79°03.89′ | 04°10.92′ | 2464 |
| 06 July 2010 | ARK-XXV/2 | PS76/132-2 | HG-I | 79°08.16′ | 06°06.35′ | 1283 |
| 07 July 2010 | ARK-XXV/2 | PS76/142-3 | HG-IV | 79°03.87′ | 04°10.38′ | 2471 |
| 13 July 2010 | ARK-XXV/2 | PS76/176-3 | HG-VII | 79°03.51′ | 03°28.81′ | 4085 |
| 14 July 2011 | ARK-XXVI/2 | PS78/140-6 | HG-I | 79°08.11′ | 06°06.27′ | 1283 |
| 17 July 2011 | ARK-XXVI/2 | PS78/143-7 | HG-IV | 79°03.86′ | 04°10.58′ | 2468 |
| 21 July 2011 | ARK-XXVI/2 | PS78/159-4 | HG-VII | 79°03.50′ | 03°28.87′ | 3988 |
| 17 July 2012 | ARK-XXVII/2 | PS80/168-7 | HG-I | 79°08.11′ | 06°06.12′ | 1283 |
| 16 July 2012 | ARK-XXVII/2 | PS80/165-8 | HG-IV | 79°03.86′ | 04°10.85′ | 2467 |
| 22 July 2012 | ARK-XXVII/2 | PS80/182-2 | HG-VII | 79°03.60′ | 03°28.46′ | 4042 |
| 26 June 2013 | MSM29 | MSM29/425-3 | HG-I | 79°08.00′ | 06°05.55′ | 1254 |
| 09 July 2013 | MSM29 | MSM29/453-1 | HG-IV | 79°04.82′ | 04°04.74′ | 2464 |
| 24 June 2014 | ARK-XXVIII/2 | PS85/470-3 | HG-I | 79°08.01′ | 06°06.39′ | 1244 |
| 22 June 2014 | ARK-XXVIII/2 | PS85/460-4 | HG-IV | 79°03.91′ | 04°10.98′ | 2403 |
| Year | Site | Nematoda | Copepoda | Ostracoda | Kinorhyncha | Polychaeta | Tardigrada | Bivalvia | Gastrotricha | Platyhelminthes | Others | Total |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2000 | HG-I | 2486 | 88 | 6 | 0 | 16 | 0 | 2 | 0 | 0 | 2 | 2600 |
| 2000 | HG-IV | 557 | 6 | 0 | 0 | 5 | 0 | 0 | 0 | 0 | 0 | 568 |
| 2000 | HG-VII | 535 | 17 | 4 | 0 | 8 | 0 | 1 | 0 | 0 | 0 | 565 |
| 2001 | HG-I | 3093 | 120 | 4 | 3 | 16 | 0 | 1 | 1 | 0 | 5 | 3243 |
| 2001 | HG-IV | 1769 | 29 | 10 | 3 | 1 | 3 | 2 | 0 | 0 | 3 | 1820 |
| 2001 | HG-VII | 784 | 14 | 1 | 0 | 5 | 0 | 2 | 0 | 0 | 2 | 808 |
| 2002 | HG-I | 2516 | 108 | 4 | 0 | 13 | 0 | 2 | 2 | 0 | 6 | 2651 |
| 2002 | HG-IV | 2015 | 25 | 3 | 1 | 3 | 3 | 1 | 1 | 0 | 3 | 2055 |
| 2002 | HG-VII | 590 | 43 | 8 | 4 | 3 | 1 | 4 | 0 | 0 | 11 | 664 |
| 2003 | HG-I | 2392 | 110 | 4 | 0 | 3 | 0 | 0 | 0 | 0 | 11 | 2520 |
| 2003 | HG-IV | 1563 | 32 | 0 | 1 | 3 | 8 | 0 | 0 | 0 | 5 | 1612 |
| 2003 | HG-VII | 387 | 5 | 1 | 0 | 9 | 0 | 0 | 0 | 0 | 4 | 406 |
| 2004 | HG-I | 3113 | 141 | 11 | 2 | 8 | 1 | 4 | 0 | 0 | 18 | 3298 |
| 2004 | HG-IV | 827 | 22 | 2 | 9 | 5 | 6 | 0 | 0 | 0 | 12 | 883 |
| 2004 | HG-VII | 134 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 13 | 148 |
| 2005 | HG-I | 2050 | 109 | 6 | 5 | 6 | 0 | 0 | 6 | 18 | 6 | 2206 |
| 2005 | HG-IV | 774 | 33 | 2 | 1 | 4 | 2 | 0 | 1 | 0 | 1 | 818 |
| 2005 | HG-VII | 299 | 8 | 1 | 0 | 3 | 0 | 0 | 0 | 1 | 0 | 312 |
| 2006 | HG-I | 1813 | 95 | 6 | 0 | 7 | 0 | 2 | 0 | 1 | 7 | 1931 |
| 2006 | HG-IV | 1371 | 15 | 2 | 0 | 4 | 4 | 0 | 0 | 1 | 0 | 1397 |
| 2006 | HG-VII | 333 | 9 | 3 | 2 | 4 | 0 | 0 | 1 | 0 | 0 | 352 |
| 2007 | HG-I | 2251 | 195 | 9 | 2 | 4 | 0 | 4 | 0 | 0 | 16 | 2481 |
| 2007 | HG-IV | 2854 | 85 | 5 | 4 | 9 | 0 | 2 | 0 | 0 | 6 | 2965 |
| 2007 | HG-VII | 338 | 4 | 0 | 0 | 9 | 0 | 0 | 0 | 0 | 8 | 359 |
| 2008 | HG-I | 2678 | 130 | 6 | 2 | 5 | 2 | 15 | 0 | 0 | 23 | 2861 |
| 2008 | HG-IV | 1239 | 38 | 3 | 1 | 4 | 4 | 2 | 1 | 0 | 10 | 1302 |
| 2008 | HG-VII | 214 | 25 | 0 | 0 | 8 | 0 | 1 | 0 | 0 | 1 | 249 |
| 2009 | HG-I | 2556 | 166 | 16 | 4 | 9 | 0 | 4 | 1 | 3 | 20 | 2779 |
| 2009 | HG-IV | 1097 | 26 | 2 | 1 | 3 | 3 | 0 | 2 | 2 | 4 | 1140 |
| 2010 | HG-I | 1340 | 19 | 0 | 0 | 5 | 0 | 0 | 0 | 0 | 3 | 1367 |
| 2010 | HG-IV | 467 | 27 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 496 |
| 2010 | HG-VII | 281 | 2 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 287 |
| 2011 | HG-I | 607 | 44 | 1 | 0 | 4 | 0 | 7 | 0 | 0 | 2 | 665 |
| 2011 | HG-IV | 536 | 25 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 563 |
| 2011 | HG-VII | 187 | 5 | 0 | 0 | 5 | 0 | 1 | 0 | 0 | 0 | 198 |
| 2012 | HG-I | 1704 | 31 | 5 | 0 | 5 | 0 | 0 | 0 | 0 | 1 | 1746 |
| 2012 | HG-IV | 889 | 14 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 905 |
| 2012 | HG-VII | 523 | 3 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 528 |
| 2013 | HG-I | 1753 | 33 | 0 | 1 | 5 | 0 | 0 | 0 | 0 | 0 | 1792 |
| 2013 | HG-IV | 1054 | 13 | 2 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 1071 |
| 2014 | HG-I | 1979 | 43 | 2 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 2025 |
| 2014 | HG-IV | 626 | 9 | 1 | 0 | 2 | 0 | 0 | 0 | 0 | 1 | 639 |
| Main Test | Pairwise Comparisons | |
|---|---|---|
| Years | p–Values * | |
| HG-I | 2011 < 2001 | 0.0005 |
| H (14, n = 225) = 43.39; p = 0.0001 | 2011 < 2002 | 0.0199 |
| 2011 < 2003 | 0.0390 | |
| 2011 < 2004 | 0.0002 | |
| 2011 < 2008 | 0.0052 | |
| 2011 < 2009 | 0.0069 | |
| HG-IV | 2002 > 2000 | 0.0190 |
| H (14, n = 225) = 58.34; p = 0.0000 | 2002 > 2010 | 0.0084 |
| 2002 > 2011 | 0.0153 | |
| 2002 > 2014 | 0.0410 | |
| 2007 > 2001 | 0.0003 | |
| 2007 > 2004 | 0.0446 | |
| 2007 > 2005 | 0.0082 | |
| 2007 > 2010 | 0.0001 | |
| 2007 > 2011 | 0.0002 | |
| 2007 > 2014 | 0.0006 | |
| HG-VII | no pairwise significant differences | |
| H (11, n = 180) = 21.41; p = 0.0294 | ||
| Sediment Depth (cm) | HG-I | HG-IV | HG-VII |
|---|---|---|---|
| 0–1 | 5.62 | 17.74 | 30.75 |
| 1–2 | 5.09 | 20.89 | 30.75 |
| 2–3 | 6.55 | 24.45 | 40.82 |
| 3–4 | 14.33 | 22.22 | 42.42 |
| 4–5 | 9.08 | 21.30 | 41.55 |
| Year | Site | H2O | AFDW | CHLA | PHAEO | CPE | %CHLA | LIPIDS |
|---|---|---|---|---|---|---|---|---|
| 2000 | HG-I | 65.97 | 138.70 | 3.69 | 8.49 | 12.19 | 30.30 | 23.65 |
| 2000 | HG-IV | 52.90 | 141.67 | 0.68 | 5.35 | 6.03 | 11.24 | 14.77 |
| 2000 | HG-VII | 32.67 | 121.41 | 0.36 | 3.78 | 4.14 | 8.59 | 11.20 |
| 2001 | HG-I | 65.32 | 130.42 | 1.93 | 10.61 | 12.54 | 15.39 | 12.40 |
| 2001 | HG-IV | 51.97 | 117.73 | 0.40 | 5.55 | 5.95 | 6.68 | 11.50 |
| 2001 | HG-VII | 38.35 | 124.07 | 0.01 | 1.44 | 1.45 | 0.76 | 15.88 |
| 2002 | HG-I | 66.31 | 117.00 | 1.74 | 10.12 | 11.86 | 14.69 | 7.60 |
| 2002 | HG-IV | 50.26 | 113.03 | 0.35 | 4.98 | 5.33 | 6.48 | 3.57 |
| 2002 | HG-VII | 42.91 | 102.06 | 0.27 | 4.58 | 4.85 | 5.49 | 16.13 |
| 2003 | HG-I | 65.69 | 110.89 | 2.46 | 12.36 | 14.82 | 16.59 | 14.77 |
| 2003 | HG-IV | 52.39 | 99.85 | 0.98 | 9.40 | 10.39 | 9.48 | 9.29 |
| 2003 | HG-VII | 35.58 | 92.77 | 0.23 | 7.85 | 8.07 | 2.81 | 11.29 |
| 2004 | HG-I | 66.31 | 100.69 | 1.77 | 9.82 | 11.59 | 15.31 | 11.93 |
| 2004 | HG-IV | 52.54 | 105.14 | 0.80 | 5.26 | 6.06 | 13.26 | 3.88 |
| 2004 | HG-VII | 37.59 | 91.11 | 0.23 | 2.23 | 2.46 | 9.44 | 3.55 |
| 2005 | HG-I | 65.30 | 110.70 | 1.28 | 9.10 | 10.38 | 12.34 | 11.71 |
| 2005 | HG-IV | 49.10 | 95.24 | 1.31 | 9.74 | 11.05 | 11.90 | 4.20 |
| 2005 | HG-VII | 39.55 | 97.83 | 0.48 | 3.65 | 4.13 | 11.59 | 5.43 |
| 2006 | HG-I | 52.76 | 69.28 | 1.13 | 7.14 | 8.28 | 13.70 | 13.91 |
| 2006 | HG-IV | 49.03 | 67.23 | 0.52 | 3.69 | 4.21 | 12.37 | 8.05 |
| 2006 | HG-VII | 34.84 | 56.60 | 0.18 | 1.90 | 2.07 | 8.52 | 6.43 |
| 2007 | HG-I | 66.62 | 135.38 | 3.39 | 21.96 | 25.35 | 13.38 | 13.65 |
| 2007 | HG-IV | 49.83 | 100.45 | 1.94 | 11.78 | 13.73 | 14.16 | 9.36 |
| 2007 | HG-VII | 33.66 | 82.15 | 0.88 | 6.24 | 7.11 | 12.31 | 5.99 |
| 2008 | HG-I | 65.49 | 162.94 | 2.57 | 28.22 | 30.79 | 8.35 | 36.39 |
| 2008 | HG-IV | 49.59 | 102.66 | 0.70 | 11.78 | 12.48 | 5.64 | 17.76 |
| 2008 | HG-VII | 31.50 | 83.44 | 0.49 | 6.06 | 6.55 | 7.46 | 20.34 |
| 2009 | HG-I | 66.11 | 125.18 | 5.73 | 31.10 | 36.83 | 15.56 | 11.69 |
| 2009 | HG-IV | 50.34 | 119.75 | 1.89 | 11.01 | 12.89 | 14.63 | 6.70 |
| 2010 | HG-I | 63.93 | 107.02 | 2.27 | 20.52 | 22.79 | 9.97 | 9.45 |
| 2010 | HG-IV | 50.63 | 101.34 | 1.83 | 7.59 | 9.42 | 19.46 | 7.97 |
| 2010 | HG-VII | 35.72 | 90.37 | 1.68 | 5.37 | 7.05 | 23.83 | 6.82 |
| 2011 | HG-I | n.d. | n.d. | 1.78 | 15.89 | 17.67 | 10.05 | 24.25 |
| 2011 | HG-IV | 50.34 | 103.45 | 0.94 | 8.54 | 9.48 | 9.93 | 12.79 |
| 2011 | HG-VII | 34.85 | 87.92 | 0.59 | 5.53 | 6.12 | 9.66 | 11.72 |
| 2012 | HG-I | 67.37 | 88.70 | 2.11 | 21.20 | 23.30 | 9.04 | 7.02 |
| 2012 | HG-IV | 49.92 | 88.70 | 1.12 | 11.20 | 12.33 | 9.12 | 6.33 |
| 2012 | HG-VII | 47.47 | 81.86 | 1.07 | 12.38 | 13.45 | 7.92 | 9.39 |
| 2013 | HG-I | 64.78 | 125.57 | 4.99 | 32.95 | 37.94 | 13.16 | 25.64 |
| 2013 | HG-IV | 52.21 | 120.79 | 1.92 | 16.08 | 18.00 | 10.67 | 6.50 |
| 2014 | HG-I | 64.99 | 83.36 | 3.29 | 27.24 | 30.54 | 10.79 | 6.13 |
| 2014 | HG-IV | 48.85 | 137.11 | 1.04 | 9.46 | 10.50 | 9.93 | 4.10 |
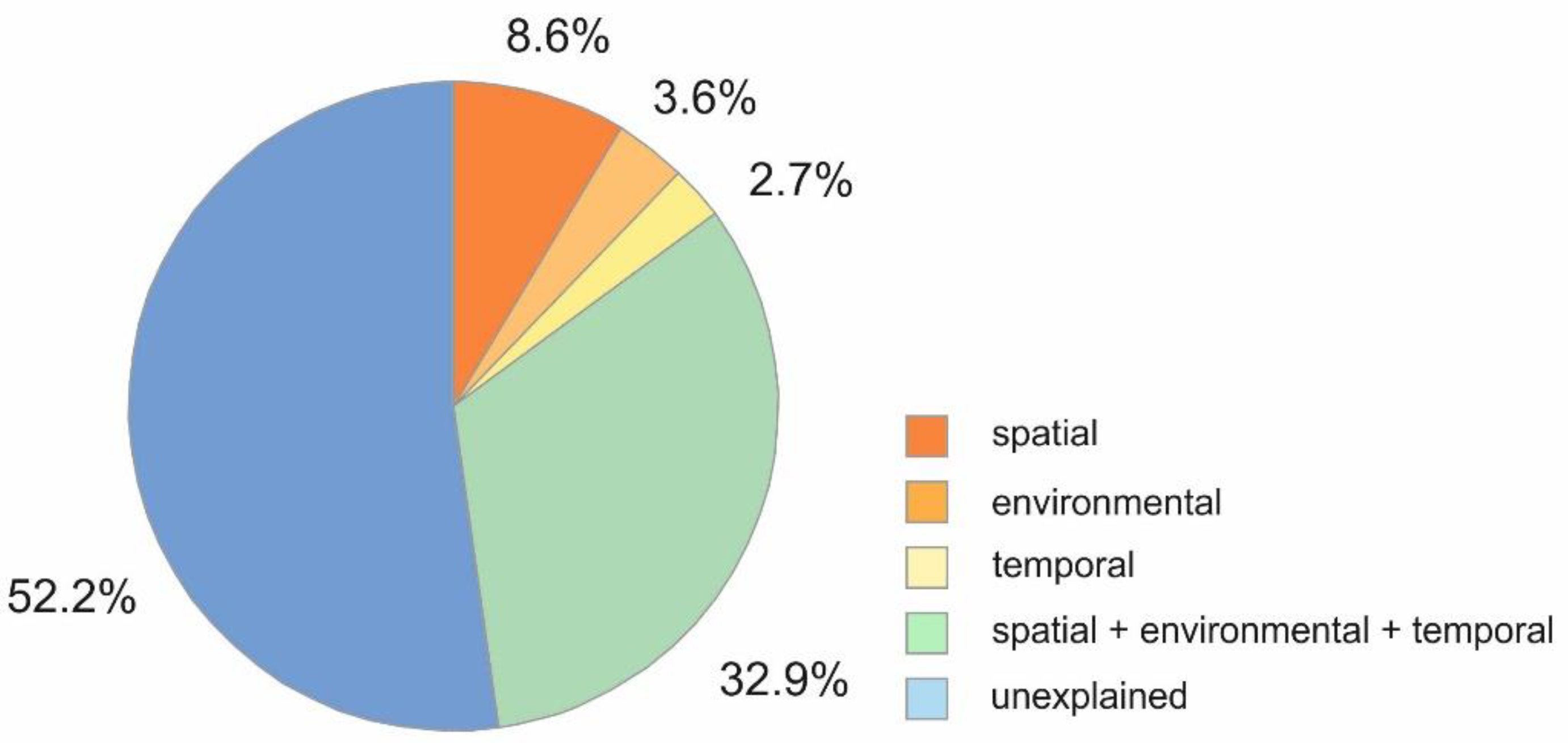
| Parameter | Prop. | Prop. (cumul.) | R2adj (cumul.) |
|---|---|---|---|
| Marginal Test | |||
| Environmental ** | |||
| Water content ** | 0.31 | - | - |
| Total organic matter ** | 0.01 | - | - |
| Chlorophyll a | 0.15 | - | - |
| Phospholipid concentrations ** | 0.05 | - | - |
| Spatial ** | |||
| Water depth ** | 0.19 | - | - |
| Sediment depth ** | 0.22 | - | - |
| Temporal * | |||
| Year * | 0.02 | - | - |
| Sequential Test | |||
| Environmental first | |||
| Environmental ** | 0.36 | 0.36 | 0.34 |
| Spatial ** | 0.10 | 0.45 | 0.43 |
| Temporal ** | 0.03 | 0.48 | 0.46 |
| Spatial first | |||
| Spatial ** | 0.42 | 0.42 | 0.41 |
| Temporal ** | 0.02 | 0.44 | 0.43 |
| Environmental ** | 0.04 | 0.48 | 0.46 |
| Temporal first | |||
| Temporal * | 0.02 | 0.02 | 0.01 |
| Environmental ** | 0.38 | 0.39 | 0.38 |
| Spatial ** | 0.09 | 0.48 | 0.46 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soltwedel, T.; Grzelak, K.; Hasemann, C. Spatial and Temporal Variation in Deep-Sea Meiofauna at the LTER Observatory HAUSGARTEN in the Fram Strait (Arctic Ocean). Diversity 2020, 12, 279. https://doi.org/10.3390/d12070279
Soltwedel T, Grzelak K, Hasemann C. Spatial and Temporal Variation in Deep-Sea Meiofauna at the LTER Observatory HAUSGARTEN in the Fram Strait (Arctic Ocean). Diversity. 2020; 12(7):279. https://doi.org/10.3390/d12070279
Chicago/Turabian StyleSoltwedel, Thomas, Katarzyna Grzelak, and Christiane Hasemann. 2020. "Spatial and Temporal Variation in Deep-Sea Meiofauna at the LTER Observatory HAUSGARTEN in the Fram Strait (Arctic Ocean)" Diversity 12, no. 7: 279. https://doi.org/10.3390/d12070279
APA StyleSoltwedel, T., Grzelak, K., & Hasemann, C. (2020). Spatial and Temporal Variation in Deep-Sea Meiofauna at the LTER Observatory HAUSGARTEN in the Fram Strait (Arctic Ocean). Diversity, 12(7), 279. https://doi.org/10.3390/d12070279