Assessing the Diversity of the Form of Age-Specific Changes in Adult Mortality from Captive Mammalian Populations


Abstract
:1. Introduction
2. Materials and Methods
2.1. Dataset
2.2. Age-Dependent Models of Mortality Changes during the Adult Stage
- (1)
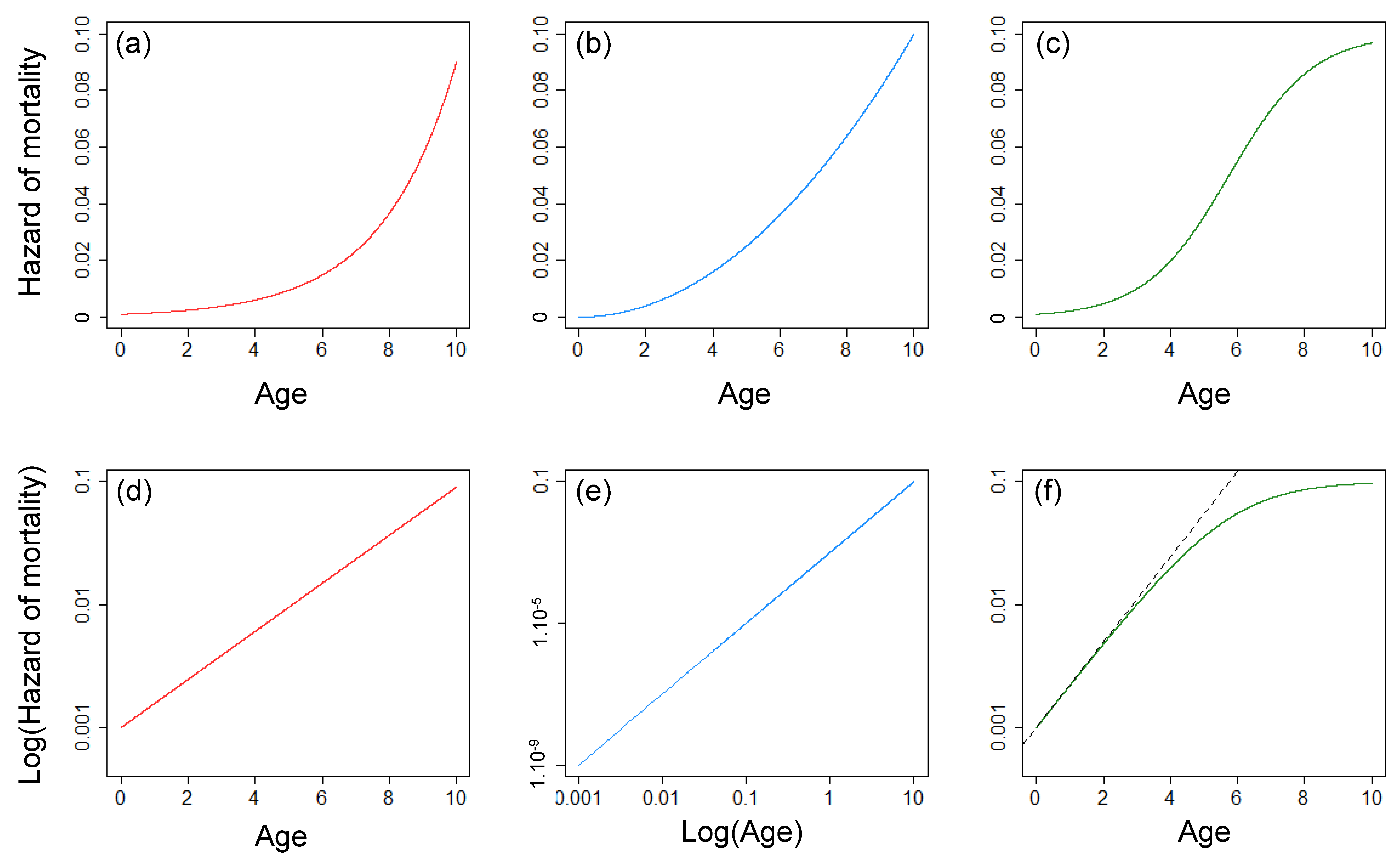
- The Gompertz model [1] is a two-parameter model in which the hazard of mortality follows an exponential increase with age (see Equation (1) and Figure 1a,d).where µ(x) is the hazard of mortality for the Gompertz model with representing the basal mortality rate and representing the exponential rate of increase.
- (2)
- The Weibull model [21] is also a two-parameter model, but it is characterized by an exponential increase with the logarithm of age (see Equation (2) and Figure 1b,e).where µ(x) is the hazard of mortality for the Weibull model with b1 representing the scale parameter and b0 representing the shape parameter.µ(x) = b0b1b0xb0−1
- (3)
- The logistic model [26] is a three-parameter model that is equivalent to the Gompertz model at young age but with a rate of increase that decreases with age (see Equation (3) and Figure 1c,f). This model is often used to model a Gompertz increase associated with a strong selective disappearance occurring in a population (i.e., populations with heterogeneous mortality rates among individuals resulting in an increase of the proportion of high-quality individuals with increasing age [27]).where µ(x) is the hazard of mortality for the logistic model with b0 and b1 corresponding to the parameters a and b of the Gompertz model, respectively, and b2 describing the rate of deceleration of the speed of mortality increase.
2.3. Statistical Analysis
3. Results
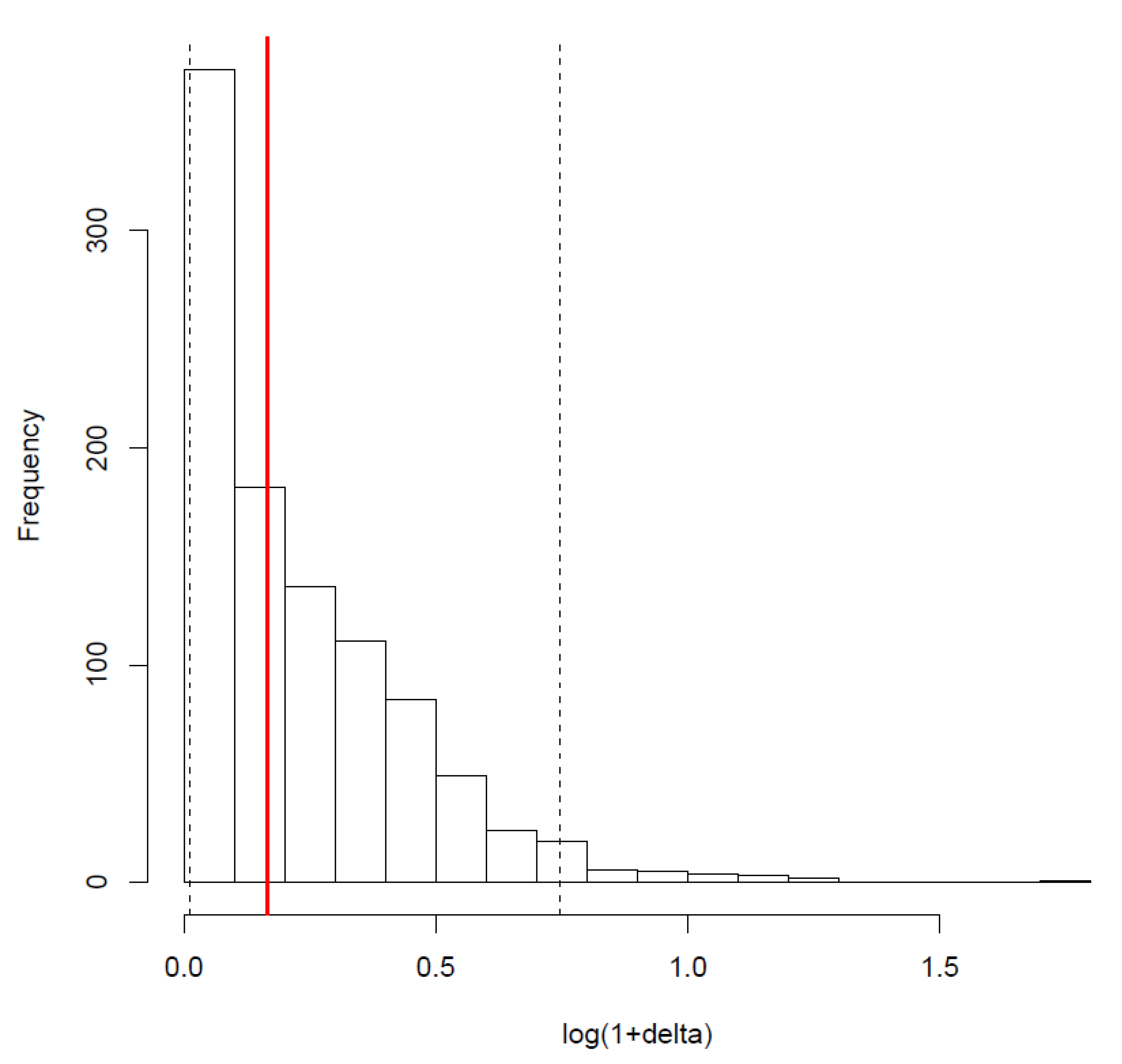
3.1. Form of the Senescent Phase
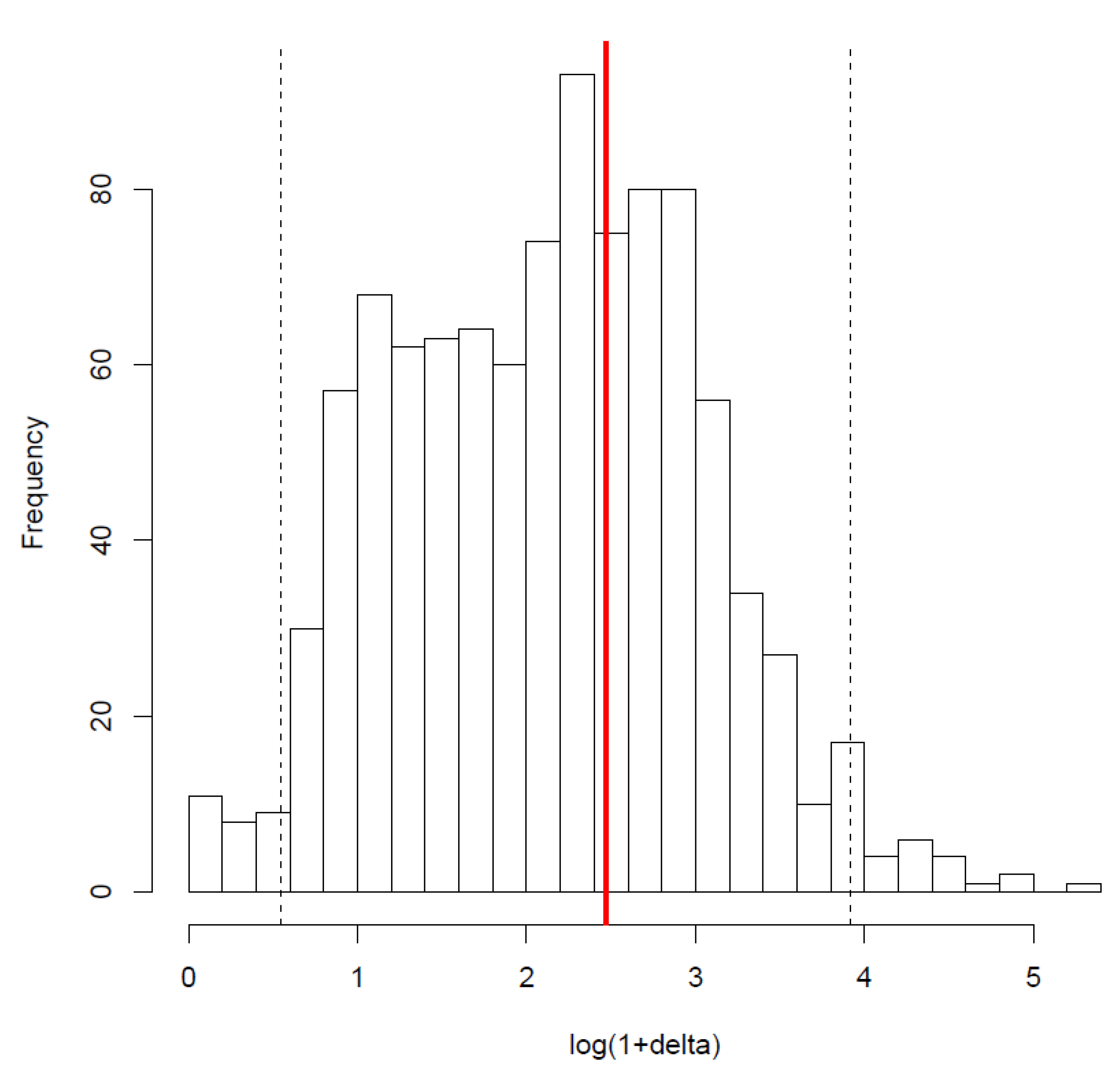
3.2. Structure of the Model
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A. Equations of the Mortality Hazard for the Nine Different Mortality Models Fitted Sequentially for Mammal Species (see the Main Text for More Details on the Meaning of Each Parameter)
| Gompertz models |
| Weibull models |
| Logistic models |
Appendix B. Phylogenetic Signal Associated with the Form of Age Dependence during the Senescent Stage and the Model Structure


References
- Gompertz, B., XXIV. On the nature of the function expressive of the law of human mortality, and on a new mode of determining the value of life contingencies. In a letter to Francis Baily, Esq. FRS &c. Philos. Trans. R. Soc. Lond. 1825, 115, 513–583. [Google Scholar] [CrossRef]
- Nussey, D.H.; Froy, H.; Lemaitre, J.-F.; Gaillard, J.-M.; Austad, S.N. Senescence in natural populations of animals: Widespread evidence and its implications for bio-gerontology. Ageing Res. Rev. 2013, 12, 214–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caughley, G. Mortality Patterns in Mammals. Ecology 1966, 47, 906–918. [Google Scholar] [CrossRef]
- Gimenez, O.; Viallefont, A.; Charmantier, A.; Pradel, R.; Cam, E.; Brown, C.R.; Anderson, M.D.; Brown, M.B.; Covas, R.; Gaillard, J. The Risk of Flawed Inference in Evolutionary Studies When Detectability Is Less than One. Am. Nat. 2008, 172, 441–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebreton, J.-D.; Burnham, K.P.; Clobert, J.; Anderson, D.R. Modeling Survival and Testing Biological Hypotheses Using Marked Animals: A Unified Approach with Case Studies. Ecol. Monogr. 1992, 62, 67–118. [Google Scholar] [CrossRef] [Green Version]
- Clutton-Brock, T.; Sheldon, B.C. Individuals and populations: The role of long-term, individual-based studies of animals in ecology and evolutionary biology. Trends Ecol. Evol. 2010, 25, 562–573. [Google Scholar] [CrossRef]
- Choquet, R.; Reboulet, A.-M.; Pradel, R.; Giménez, O.; Lebreton, J.-D. M–SURGE: New software specifically designed for multistate capture–recapture models. Anim. Biodivers. Conserv. 2004, 27, 207–215. [Google Scholar]
- Colchero, F.; Jones, O.R.; Rebke, M. BaSTA: An R package for Bayesian estimation of age-specific survival from incomplete mark–recapture/recovery data with covariates. Methods Ecol. Evol. 2012, 3, 466–470. [Google Scholar] [CrossRef]
- Gaillard, J.-M.; Lemaître, J.-F. An integrative view of senescence in nature. Funct. Ecol. 2020, 34, 4–16. [Google Scholar] [CrossRef] [Green Version]
- Garrott, R.A.; Eberhardt, L.L.; Otton, J.K.; White, P.J.; Chaffee, M.A. A Geochemical Trophic Cascade in Yellowstone’s Geothermal Environments. Ecosystems 2002, 5, 0659–0666. [Google Scholar] [CrossRef]
- Tidière, M.; Gaillard, J.-M.; Berger, V.; Müller, D.W.H.; Bingaman Lackey, L.; Gimenez, O.; Clauss, M.; Lemaître, J.-F. Comparative analyses of longevity and senescence reveal variable survival benefits of living in zoos across mammals. Sci. Rep. 2016, 6, 36361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lahdenperä, M.; Mar, K.U.; Courtiol, A.; Lummaa, V. Differences in age-specific mortality between wild-caught and captive-born Asian elephants. Nat. Commun. 2018, 9, 1–10. [Google Scholar]
- Jones, O.R.; Scheuerlein, A.; Salguero-Gómez, R.; Camarda, C.G.; Schaible, R.; Casper, B.B.; Dahlgren, J.P.; Ehrlén, J.; García, M.B.; Menges, E.S.; et al. Diversity of ageing across the tree of life. Nature 2014, 505, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Gaillard, J.-M.; Lemaître, J.-F. The Williams’ legacy: A critical reappraisal of his nine predictions about the evolution of senescence. Evolution 2017, 71, 2768–2785. [Google Scholar] [CrossRef] [PubMed]
- Arbuthnott, D.; Promislow, D.E.; Moorad, J. Evolutionary theory and aging. In Handbook of Theories of Aging, 3rd ed.; Springer Publishing Company: New York, NY, USA, 2016. [Google Scholar]
- Kirkwood, T.B. Deciphering death: A commentary on Gompertz (1825) ‘On the nature of the function expressive of the law of human mortality, and on a new mode of determining the value of life contingencies’. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaillard, J.-M.; Garratt, M.; Lemaître, J.-F. Senescence in mammalian life history traits. Evol. Senescence Tree Life 2017, 126–155. [Google Scholar]
- Siler, W. A Competing-Risk Model for Animal Mortality. Ecology 1979, 60, 750–757. [Google Scholar] [CrossRef]
- Tidière, M.; Gaillard, J.-M.; Müller, D.W.H.; Lackey, L.B.; Gimenez, O.; Clauss, M.; Lemaître, J.-F. Does sexual selection shape sex differences in longevity and senescence patterns across vertebrates? A review and new insights from captive ruminants. Evolution 2015, 69, 3123–3140. [Google Scholar] [CrossRef]
- Lemaître, J.-F.; Ronget, V.; Tidière, M.; Allainé, D.; Berger, V.; Cohas, A.; Colchero, F.; Conde, D.A.; Garratt, M.; Liker, A. Sex differences in adult lifespan and aging rates of mortality across wild mammals. Proc. Natl. Acad. Sci. USA 2020, 117, 8546–8553. [Google Scholar] [CrossRef]
- Pinder, J.E., III; Wiener, J.G.; Smith, M.H. The Weibull distribution: A new method of summarizing survivorship data. Ecology 1978, 59, 175–179. [Google Scholar] [CrossRef]
- Ricklefs, R.E.; Scheuerlein, A. Comparison of aging-related mortality among birds and mammals. Exp. Gerontol. 2001, 36, 845–857. [Google Scholar] [CrossRef]
- Juckett, D.A.; Rosenberg, B. Comparison of the Gompertz and Weibull functions as descriptors for human mortality distributions and their intersections. Mech. Ageing Dev. 1993, 69, 1–31. [Google Scholar] [CrossRef]
- Wilson, D.L. The analysis of survival (mortality) data: Fitting Gompertz, Weibull, and logistic functions. Mech. Ageing Dev. 1994, 74, 15–33. [Google Scholar] [CrossRef]
- Péron, G.; Lemaître, J.-F.; Ronget, V.; Tidière, M.; Gaillard, J.-M. Variation in actuarial senescence does not reflect life span variation across mammals. PLoS Biol. 2019, 17, e3000432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pletcher, S.D. Model fitting and hypothesis testing for age-specific mortality data. J. Evol. Biol. 1999, 12, 430–439. [Google Scholar] [CrossRef]
- Vaupel, J.W.; Yashin, A.I. Heterogeneity’s ruses: Some surprising effects of selection on population dynamics. Am. Stat. 1985, 39, 176–185. [Google Scholar]
- R Core Team. R: A language and environment for statistical computing. In R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2020; Available online: http://www.R-project.org/ (accessed on 1 July 2020).
- Spiegelhalter, D.J.; Best, N.G.; Carlin, B.P.; Linde, A.V.D. Bayesian measures of model complexity and fit. J. R. Stat. Soc. Ser. B (Stat. Methodol.) 2002, 64, 583–639. [Google Scholar] [CrossRef] [Green Version]
- Borges, R.; Machado, J.P.; Gomes, C.; Rocha, A.P.; Antunes, A. Measuring phylogenetic signal between categorical traits and phylogenies. Bioinformatics 2019, 35, 1862–1869. [Google Scholar] [CrossRef]
- Upham, N.S.; Esselstyn, J.A.; Jetz, W. Inferring the mammal tree: Species-level sets of phylogenies for questions in ecology, evolution, and conservation. PLoS Biol. 2019, 17, e3000494. [Google Scholar] [CrossRef]
- Barbi, E.; Lagona, F.; Marsili, M.; Vaupel, J.W.; Wachter, K.W. The plateau of human mortality: Demography of longevity pioneers. Science 2018, 360, 1459–1461. [Google Scholar] [CrossRef] [Green Version]
- Gaillard, J.-M.; Yoccoz, N.G.; Lebreton, J.-D.; Bonenfant, C.; Devillard, S.; Loison, A.; Pontier, D.; Allaine, D. Generation Time: A Reliable Metric to Measure Life-History Variation among Mammalian Populations. Am. Nat. 2005, 166, 119–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, G.C. Pleiotropy, Natural Selection, and the Evolution of Senescence. Evolution 1957, 11, 398–411. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002; ISBN 978-0-387-95364-9. [Google Scholar]
- Hamilton, W.D. The moulding of senescence by natural selection. J. Theor. Biol. 1966, 12, 12–45. [Google Scholar] [CrossRef]
- McGilchrist, C.A.; Aisbett, C.W. Regression with Frailty in Survival Analysis. Biometrics 1991, 47, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Aalen, O.O. Heterogeneity in survival analysis. Stat. Med. 1988, 7, 1121–1137. [Google Scholar] [CrossRef]
- Lynch, H.J.; Fagan, W.F. Survivorship curves and their impact on the estimation of maximum population growth rates. Ecology 2009, 90, 1116–1124. [Google Scholar] [CrossRef] [PubMed]
- Conde, D.A.; Staerk, J.; Colchero, F.; da Silva, R.; Schöley, J.; Baden, H.M.; Jouvet, L.; Fa, J.E.; Syed, H.; Jongejans, E. Data gaps and opportunities for comparative and conservation biology. Proc. Natl. Acad. Sci. USA 2019, 116, 9658–9664. [Google Scholar] [CrossRef] [Green Version]
- Jones, O.R.; Gaillard, J.-M.; Tuljapurkar, S.; Alho, J.S.; Armitage, K.B.; Becker, P.H.; Bize, P.; Brommer, J.; Charmantier, A.; Charpentier, M.; et al. Senescence rates are determined by ranking on the fast–slow life-history continuum. Ecol. Lett. 2008, 11, 664–673. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) Form of the Senescent Phase | |||
|---|---|---|---|
| Model | k | AIC | ΔAIC |
| Log(E) + Log(N) | 3 | 84.9497 | 0 |
| NULL | 1 | 85.1474 | 0.1977 |
| Log(E) | 2 | 85.2748 | 0.3251 |
| Log(N) | 2 | 86.0621 | 1.1124 |
| Log(E) * Log(N) | 4 | 87.2107 | 2.2610 |
| (b) Structure of the Model | |||
| Model | k | AIC | ΔAIC |
| NULL | 1 | 129.6015 | 0 |
| Log(E) | 2 | 130.9753 | 1.3738 |
| Log(N) | 2 | 131.0201 | 1.4186 |
| Log(E) + Log(N) | 3 | 132.1048 | 2.5033 |
| Log(E) * Log(N) | 4 | 135.4106 | 5.8091 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ronget, V.; Lemaître, J.-F.; Tidière, M.; Gaillard, J.-M. Assessing the Diversity of the Form of Age-Specific Changes in Adult Mortality from Captive Mammalian Populations. Diversity 2020, 12, 354. https://doi.org/10.3390/d12090354
Ronget V, Lemaître J-F, Tidière M, Gaillard J-M. Assessing the Diversity of the Form of Age-Specific Changes in Adult Mortality from Captive Mammalian Populations. Diversity. 2020; 12(9):354. https://doi.org/10.3390/d12090354
Chicago/Turabian StyleRonget, Victor, Jean-François Lemaître, Morgane Tidière, and Jean-Michel Gaillard. 2020. "Assessing the Diversity of the Form of Age-Specific Changes in Adult Mortality from Captive Mammalian Populations" Diversity 12, no. 9: 354. https://doi.org/10.3390/d12090354
APA StyleRonget, V., Lemaître, J. -F., Tidière, M., & Gaillard, J. -M. (2020). Assessing the Diversity of the Form of Age-Specific Changes in Adult Mortality from Captive Mammalian Populations. Diversity, 12(9), 354. https://doi.org/10.3390/d12090354