Incidence of Galls on Sympatric California Oaks: Ecological and Physiological Perspectives



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Collection
2.2.1. Gall Incidence Surveys
2.2.2. Diversity of Galls on Oaks
2.2.3. Leaf Physiological Responses to Galling
2.2.4. Statistical Analyses
3. Results
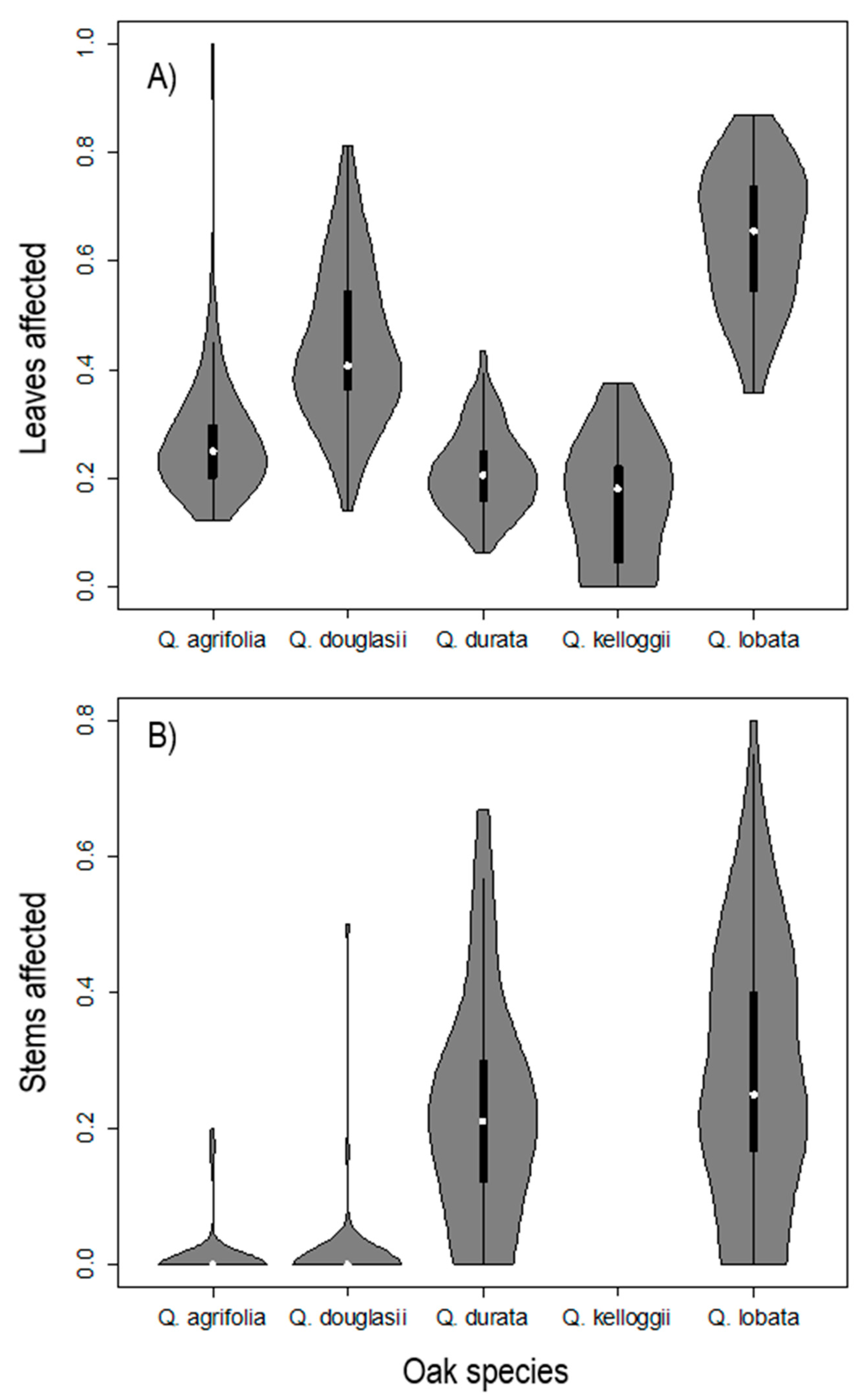
3.1. Gall Incidence Surveys
3.2. Diversity of Oak Galls
3.3. Leaf Physiological Responses to Galling
4. Discussion
4.1. Gall Incidence Surveys
4.2. Diversity of Oak Galls
4.3. Leaf Physiological Responses to Galling
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Price, P.W.; Fernandes, G.W.; Waring, G.L. Adaptive nature of insect galls. Environ. Entomol. 1987, 16, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Stone, G.N.; Schönrogge, K. The adaptive significance of insect gall morphology. Trends Ecol. Evol. 2003, 18, 512–522. [Google Scholar] [CrossRef]
- Shorthouse, J.D.; Rohfritsche, O. Biology of Insect-Induced Galls; Oxford University Press: New York, NY, USA, 1992. [Google Scholar]
- Fernandes, G.W.; Price, P.W. The adaptive significance of insect gall distribution: Survivorship of species in xeric and mesic habitats. Oecologia 1992, 90, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Betancourt, E.K.; Soto, P.H.; Cortés, N.C.; Anaya, M.R.; Estrella, A.H.; Oyama, K. Ecological Genomics of Plant-Insect Interactions: The Case of Wasp-Induced Galls. In Evolutionary Ecology of Plant-Herbivore Interaction; Núñez-Farfán, J., Valverde, P., Eds.; Springer: Cham, Switzerland, 2020; pp. 315–341. [Google Scholar]
- Espírito-Santo, M.M.; Fernandes, G.W. How many species of gall-inducing insects are there on Earth, and where are they? Ann. Entomol. Soc. Am. 2007, 100, 95–99. [Google Scholar]
- Price, P.W. Adaptive radiation of gall-inducing insects. Bas. Appl. Ecol. 2005, 6, 413–421. [Google Scholar] [CrossRef]
- Schultz, J.C.; Edger, P.P.; Body, M.J.A.; Appel, H.M. A galling insect activates plant reproductive programs during gall development. Sci. Rep. 2019, 9, 1833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shorthouse, J.D.; Wool, D.; Raman, A. Gall-inducing insects—Nature’s most sophisticated herbivores. Bas. Appl. Ecol 2005, 6, 407–411. [Google Scholar] [CrossRef]
- Julião, G.R.; Venticinque, E.M.; Fernandes, G.W.; Price, P.W. Unexpected high diversity of galling insects in the Amazonian upper canopy: The savanna out there. PLoS ONE 2014, 9, e114986. [Google Scholar] [CrossRef]
- Fernandes, G.W.; Price, P.W. Biogeographical gradients in galling species richness. Oecologia 1988, 76, 161–167. [Google Scholar] [CrossRef]
- Goncalves-Alvim, S.J.; Fernandes, G.W. Biodiversity of galling insects: Historical, community, and habitat effects in four neotropical savannas. Biod. Conserv. 2001, 10, 79–98. [Google Scholar] [CrossRef]
- Abrahamson, W.G.; Melika, G.; Scrafford, R.; Csóka, G. Gall-inducing insects provide insights into plant systematic relationships. Am. J. Bot. 1998, 85, 1159–1165. [Google Scholar] [CrossRef]
- Tyler, C.M.; Kuhn, B.; Davis, F.W. Demography and recruitment limitations of three oak species in California. Q. Rev. Biol. 2006, 81, 127–152. [Google Scholar] [CrossRef] [PubMed]
- Davis, F.W.; Baldocchi, D.D.; Tyler, C.M. Oak Woodlands. In Ecosystems of California; Mooney, H.A., Zavaleta, E., Eds.; University of California Press: Berkeley, CA, USA, 2016; pp. 509–534. [Google Scholar]
- Brown, L.R. Insects feeding on California oak trees. In Proceedings of the Symposium on the Ecology, Management, and Utilization of California Oaks, Claremont, CA, USA, 26–28 June 1979; Pacific Southwest Forest and Range Experiment Station: Claremont, CA, USA, 1981; Volume 44, p. 184. [Google Scholar]
- Swiecki, T.J.; Bernhardt, E.A. A Field Guide to Insects and Diseases of California Oaks; Gen. Tech Rep. PSW-GTR-197; Pacific Southwest Research Station, Forest Service, U.S. Department of Agriculture: Albany, CA, USA, 2006.
- Fürjes-Mikó, Á.; Csősz, S.; Csóka, G. Ants inhabiting oak Cynipid galls in Hungary. North Western J. Zool. 2006, 16, 95–98. [Google Scholar]
- Wetzel, W.C.; Screen, R.M.; Li, I.; McKenzie, J.; Phillips, K.A.; Cruz, M.; Zhang, W.; Greene, A.; Lee, E.; Singh, N.; et al. Ecosystem engineering by a gall-forming wasp indirectly suppresses diversity and density of herbivores on oak trees. Ecology 2016, 97, 427–438. [Google Scholar] [CrossRef] [Green Version]
- Price, P. The Plant Vigor Hypothesis and Herbivore Attack. Oikos 1991, 62, 244–251. [Google Scholar] [CrossRef] [Green Version]
- Koricheva, J.; Larsson, S. Insect performance on experimentally stressed woody plants: A meta-analysis. Ann. Rev. Entomol. 1998, 43, 195–216. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, G.W. Hypersensitivity as a phenotypic basis of plant-induced resistance against a galling insect (Diptera: Cecidomyiidae). Environ. Entomol. 1998, 27, 260–267. [Google Scholar] [CrossRef]
- Cornelissen, T.; Fernandes, G.W.; Vasconcellos-Neto, J. Size does matter: Variation in herbivory between and within plants and the plant vigor hypothesis. Oikos 2008, 117, 1121–1130. [Google Scholar] [CrossRef]
- Pires, C.S.; Price, P.W. Patterns of host plant growth and attack and establishment of gall-inducing wasp (Hymenoptera: Cynipidae). Environ. Entomol. 2000, 29, 49–54. [Google Scholar] [CrossRef]
- Kirst, G.O.; Rapp, H. Physiology of gall of Mikiola fagi HTG on leaves of Fagus silvatica: Translocation of C-14 labeled assimilates from host leaf and adjacent leaves onto gall. Biochem. Physiol. Pflanz. 1974, 165, 445–455. [Google Scholar] [CrossRef]
- Larson, K.C.; Whitham, T.G. Manipulation of food resources by a gall-forming aphid: The physiology of sink-source interactions. Oecologia 1991, 88, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Wool, D.; Aloni, R.; Ben-Zvi, O.; Wollberg, M. A galling aphid furnishes its home with a built-in pipeline to the host food supply. Entomol. Exp. Appl. 1999, 91, 183–186. [Google Scholar] [CrossRef]
- Giron, D.; Huguet, E.; Stone, G.N.; Body, M. Insect-induced effects on plants and possible effectors used by galling and leaf-mining insects to manipulate their host plant. J. Insect. Physiol. 2016, 84, 70–89. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.O.; de Carvalho Junior, A.R.; da Silva, T.M.; Castro, R.N.; da Silva, T.M.; de Carvalho, M.G. Distribution of metabolites in galled and non-galled leaves of Clusia lanceolata and its antioxidant activity. Rev. Bras. Farmacogn. 2014, 24, 617–625. [Google Scholar] [CrossRef] [Green Version]
- Leege, L.M. The relationship between psyllid leaf galls and redbay (Persea borbonia) fitness traits in sun and shade. Plant Ecol. 2006, 184, 203–212. [Google Scholar] [CrossRef]
- Fernandes, G.W.; Santos, J.C.; Castro, F.M.C.; Castilho, A. Impact of gall midge Parkiamyia paraensis (Diptera, Cecidomyiidae) on the Amazon plant Parkia pendula (Fabacaeae). Rev. Bras. Entomol. 2007, 51, 471–475. [Google Scholar] [CrossRef]
- Marini-Filho, O.J.; Fernandes, G.W. Stem galls drain nutrients and decrease shoot performance in Diplusodon orbicularis (Lythraceae). Arthropod-Plant Interact. 2012, 6, 121–128. [Google Scholar] [CrossRef]
- Vuorisalo, T.; Walls, M.; Kuitunen, H. Gall mite (Eriophyes laevis) infestation and leaf removal affect growth of leaf area in black alder (Alnus glutinosa) short shoots. Oecologia 1990, 84, 122–125. [Google Scholar] [CrossRef]
- Hartley, S.E. The chemical composition of plant galls: Are levels of nutrients and secondary compounds controlled by the gall-former? Oecologia 1998, 113, 492–501. [Google Scholar] [CrossRef]
- Chalker-Scott, L. Environmental significance of anthocyanins in plant stress responses. Photochem. Photobiol. 1999, 70, 1–9. [Google Scholar] [CrossRef]
- Perea, R.; Fernandes, G.W.; Dirzo, R. Embryo size as a tolerance trait against seed predation: Contribution of embryo-damaged seeds to plant regeneration. Perspect. Plant Ecol. Evol. Syst. 2018, 31, 7–16. [Google Scholar] [CrossRef]
- López-Sánchez, A.; Johnson, I.; Dirzo, R.; Perea, R. Matching species traits and microsites improves regeneration in mixed oak woodlands. Appl. Veg. Sci. 2020. [Google Scholar] [CrossRef]
- Bryant, J.P.; Chapin, F.S.; Klein, D.R. Carbon/nutrient balance of boreal plants in relation to vertebrate herbivory. Oikos 1983, 40, 357–368. [Google Scholar] [CrossRef] [Green Version]
- Zavaleta, E.; Kettley, L. Ecosystem change along a woody invasion chronosequence in a California grassland. J. Arid Environ. 2006, 66, 290–306. [Google Scholar] [CrossRef]
- Coleman, R.G. Geologic nature of the Jasper Ridge Biological Preserve, San Francisco Peninsula, California. Internat. Geol. Rev. 2004, 46, 629–637. [Google Scholar] [CrossRef]
- Perea, R.; López-Sánchez, A.; Dirzo, R. Differential tree recruitment in California oak savannas: Are evergreen oaks replacing deciduous oaks? For. Ecol. Manag. 2017, 399, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Koenig, W.D. The effects of tannins and lipids on digestion of acorns by acorn woodpeckers. Auk 1991, 108, 79–88. [Google Scholar]
- Koenig, W.D.; Faeth, S.H. Effects of storage on tannin and protein content of cached acorns. Southwest. Nat. 1998, 43, 170–175. [Google Scholar]
- Steinberg, P.D. Quercus agrifolia. In Fire Effects Information System; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer): Fort Collins, CO, USA, 2002. [Google Scholar]
- Little, R.; Swiecki, T.J.; Tietje, W. Oak Woodland Invertebrates: The Little Things Count; University of California Division of Agriculture and Natural Resources: Oakland, CA, USA, 2001. [Google Scholar]
- Koehler, C.S. Control of Asterolecanium and cynipid leaf galls on oak in northern California. J. Econ. Entomol. 1964, 57, 579–581. [Google Scholar] [CrossRef]
- Goulas, Y.; Cerovic, Z.G.; Cartelat, A.; Moya, I. Dualex: A new instrument for field measurements of epidermal UV-absorbance by chlorophyll fluorescence. Appl. Optics 2004, 43, 4488–4496. [Google Scholar] [CrossRef]
- Pfündel, E.E.; Ben Ghozlen, N.; Meyer, S.; Cerovic, Z.G. Investigating UV screening in leaves by two different types of portable UV fluorimeters reveals in vivo screening by anthocyanins and carotenoids. Photosynth. Res. 2007, 93, 205–221. [Google Scholar] [CrossRef] [PubMed]
- Cerovic, Z.G.; Masdoumier, G.; Ben Ghozlen, N.; Latouche, G. A new optical leaf-clip meter for simultaneous non-destructive assessment of leaf chlorophyll and epidermal flavonoids. Physiol. Plant. 2012, 3, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Cartelat, A.; Cerovic, Z.G.; Goulas, Y.; Meyer, S.; Lelarge, C.; Prioul, J.L.; Barbottin, A.; Jeuffroy, M.H.; Gate, P.; Agati, G.; et al. Optically assessed contents of leaf polyphenolics and chlorophyll as indicators of nitrogen deficiency in wheat (Triticum aestivum L.). Field Crops Res. 2005, 91, 35–49. [Google Scholar] [CrossRef]
- Agati, G.; Tuccio, L.; Kusznierewicz, B.; Chmiel, T.; Bartoszek, A.; Kowalski, A.; Grzegorzewska, M.; Kosson, R.; Kaniszewski, S. Nondestructive optical sensing of flavonols and chlorophyll in white head cabbage (Brassica oleracea L. var. capitata subvar. alba) grown under different nitrogen regimens. J. Agric. Food Chem. 2016, 64, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Casa, R.; Castaldi, F.; Pascucci, S.; Pignatti, S. Chlorophyll estimation in field crops: An assessment of handheld leaf meters and spectral reflectance measurements. J. Agric. Sci. 2015, 153, 876. [Google Scholar] [CrossRef]
- Pringle, E.G.; Adams, R.I.; Broadbent, E.; Busby, P.E.; Donatti, C.I.; Kurten, E.L.; Renton, K.; Dirzo, R. Distinct Leaf-Trait Syndromes of Evergreen and Deciduous Trees in a Seasonally Dry Tropical Forest. Biotropica 2011, 43, 299–308. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.r-project.org (accessed on 15 September 2020).
- Cribari-Neto, F.; Zeileis, A. Beta Regression in R. J. Stat. Softw. 2010, 34, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.S.S. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef]
- Smart, J.; Sutherland, W.J.; Watkinson, A.R.; Gill, J.A. A new means of presenting the results of logistic regression. Bull. Ecol. Soc. Am. 2004, 85, 100–102. [Google Scholar] [CrossRef] [Green Version]
- Leckey, E.H.; Smith, D.M.; Nufio, C.R.; Fornash, K.F. Oak-insect herbivore interactions along a temperature and precipitation gradient. Acta Oecologica 2014, 61, 1–8. [Google Scholar] [CrossRef]
- Swiecki, T.J.; Bernhardt, E.A.; Arnold, R.A. Monitoring Insect and Disease Impacts on Rangeland Oaks in California. In Proceedings of the Symposium on Oak Woodlands and Hardwood Rangeland Management, Davis, CA, USA, 31 October–2 November 1990; Gen. Tech. Rep. PSW-GTR-126. Pacific Southwest: Berkeley, CA, USA, 1991; pp. 208–213. [Google Scholar]
- Kinsey, A.C. Phylogeny of cynipid genera and biological characteristics. Bull. Am. Mus. Nat. Hist. 1990, 42, 307–402. [Google Scholar]
- Evans, D. The bisexual and agamaic generations of Besbicus mirabilis (Hymenoptera: Cynipidae), and their associated insects. Can. Entomol. 1967, 99, 187–196. [Google Scholar] [CrossRef]
- Stone, G.N.; Schönrogge, K.; Atkinson, R.J.; Bellido, D.; Pujade-Villar, J. The population biology of oak gall wasps (Hymenoptera: Cynipidae). Ann. Rev. Entomol. 2002, 47, 633–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, P.W.; Fernandes, G.W.; Lara, A.C.F.; Brawn, J.; Barrios, H.; Wright, M.G.; Ribeiro, S.P.; Rothcliff, N. Global patterns in local number of insect galling species. J. Biogeogr. 1998, 25, 581–591. [Google Scholar] [CrossRef] [Green Version]
- Allen-Diaz, B.; Standiford, R.; Jackson, R.D. Oak woodlands and forests. In Terrestrial vegetation of California; University of California Press: Berkeley, CA, USA, 2007; pp. 313–338. [Google Scholar]
- Russo, R. Plant Galls of the California Region; Boxwood Press: Pacific Grove, CA, USA, 1979. [Google Scholar]
- Zargaran, M.R.; Erbilgin, N.; Ghosta, Y. Changes in Oak Gall Wasps Species Diversity(Hymenoptera: Cynipidae) in Relation to the Presence of Oak Powdery Mildew(Erysiphe alphitoides). Zool. Stud. 2012, 51, 175–184. [Google Scholar]
- Ronquist, F.; Liljeblad, J. Evolution of the gall wasp-host plant association. Evolution 2001, 55, 2503–2522. [Google Scholar]
- Keator, G.; Bazell, S. The Life of an Oak: An Intimate Portrait; Heyday Books and Oakland: California Oak Foundation: Berkeley, CA, USA, 1998. [Google Scholar]
- Abrahamson, W.G.; Hunter, M.D.; Melika, G.; Price, P.W. Cynipid gall-wasp communities correlate with oak chemistry. J. Chem. Ecol. 2003, 29, 209–223. [Google Scholar] [CrossRef]
- Cuevas-Reyes, P.; Quesada, M.; Hanson, P.; Dirzo, R.; Oyama, K.E.N. Diversity of gall-inducing insects in a Mexican tropical dry forest: The importance of plant species richness, life-forms, host plant age and plant density. J. Ecol. 2004, 92, 707–716. [Google Scholar] [CrossRef]
- Pascual-Alvarado, E.; Nieves-Aldrey, J.L.; Castillejos-Lemus, D.E.; Cuevas-Reyes, P.; Oyama, K. Diversity of galls induced by wasps (Hymenoptera: Cynipidae, Cynipini) associated with oaks (Fagaceae: Quercus) in Mexico. Bot. Sci. 2017, 95, 461–472. [Google Scholar] [CrossRef] [Green Version]
- Csóka, G.; Stone, G.N.; Melika, G. The biology, ecology and evolution of Gall-inducing Cynipidae. In Biology, Ecology and Evolution of Gall Inducing Insects; Raman, C., Schaefer, W., Withers, T.M., Eds.; CRC Press: Boca Raton, FL, USA, 2005; pp. 573–642. [Google Scholar]
- Veldtman, R.; Mcgeoch, M.A. Gall-forming insect species richness along a non-scleromorphic vegetation rainfall gradient in South Africa: The importance of plant community composition. Austral. Ecol. 2003, 28, 1–13. [Google Scholar] [CrossRef]
- Bargali, K.; Joshi, B.; Bargali, S.S.; Singh, S.P. Oaks and the biodiversity they sustain. Int. Oaks 2015, 26, 65–76. [Google Scholar]
- Maschinski, J.; Whitham, T.G. The continuum of plant responses to herbivory: The influence of plant association, nutrient availability, and timing. Am. Nat. 1989, 134, 1–19. [Google Scholar] [CrossRef]
- Thomson, V.P.; Cunningham, S.A.; Ball, M.C.; Nicotra, A.B. Compensation for herbivory by Cucumis sativus through increased photosynthetic capacity and efficiency. Oecologia 2003, 134, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Dirzo, R. Herbivory a Phytocentric Overview. In Perspectives in Plant Population Ecology; Dirzo, R., Sarikhan, J., Eds.; Sinauer Associates, Inc.: Sunderland, MA, USA, 1984; pp. 141–165. [Google Scholar]
- Fay, P.A.; Hartnett, D.C.; Knapp, A.K. Plant tolerance of gall-insect attack and gall-insect performance. Ecology 1996, 77, 521–534. [Google Scholar] [CrossRef]
- Mani, M.S. The Ecology of Plant Galls; Dr. W. Junk Publisher: The Hague, The Netherlands, 1964. [Google Scholar]
- Roychoudhury, N.; Chandra, S.; Singh, R.B. Pest status of gall making insect, Leptocybe invasa Fisher & Lasalle, in nurseries and plantations of Eucalyptus in Madhya Pradesh. Indian For. 2016, 142, 601–606. [Google Scholar]
- Fonseca, C.R.; Fleck, T.; Fernandes, G.W. Processes Driving Ontogenetic Succession of Galls in a Canopy Tree 1. Biotropica 2006, 38, 514–521. [Google Scholar] [CrossRef]
- Santos, J.C.; Fernandes, G.W. Mediation of herbivore attack and induced resistance by plant vigor and ontogeny. Acta Oecologica 2010, 36, 617–625. [Google Scholar] [CrossRef]
- Barton, K.E.; Edwards, K.F.; Koricheva, J. Shifts in woody plant defence syndromes during leaf development. Fun. Ecol. 2019, 33, 2095–2104. [Google Scholar] [CrossRef] [Green Version]
- Weis, A.E.; Walton, R.; Crego, C.L. Reactive plant tissue sites and the population biology of gall makers. Ann. Rev. Entomol. 1988, 33, 467–486. [Google Scholar] [CrossRef]
- Dawkins, R. The Extended Phenotype; Oxford University Press: Oxford, UK, 1982. [Google Scholar]
- Oliveira, D.C.; Isaias, R.M.S.; Fernandes, G.W.; Ferreira, B.G.; Carneiro, R.G.S.; Fuzaro, L. Manipulation of host plant cells and tissues by gall-inducing insects and adaptive strategies used by different feeding guilds. J. Ins. Physiol. 2016, 84, 103–113. [Google Scholar] [CrossRef]
- Florentine, S.K.; Raman, A.; Dhileepan, K. Effects of Gall Induction by Epiblema Strenuana on Gas Exchange, Nutrients, and Energetics in Parthenium Hysterophorus. BioControl 2005, 50, 787–801. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, G.W.; Castro, F.M.C.; Marques, E.S.A. Leaflet abscission caused by a gall induced by Melaphis rhois (Homoptera: Aphididae) on Rhus glabra (Anacardiaceae). Internat. J. Ecol. Environ. Sci. 1999, 25, 63–69. [Google Scholar]
- Tooker, J.F.; Rohr, J.R.; Abrahamson, W.G.; De Moraes, C.M. Gall insects can avoid and alter indirect plant defenses. New Phytol. 2008, 178, 657–671. [Google Scholar] [CrossRef] [PubMed]
- Traveset, A. Reproductive biology of Phyllyrea angustifolia. L. (Oleaceae) and effect of galling insects on its reproductive output. Bot. J. Linn. Soc. 2008, 114, 153–166. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Galling Agent | Family | Host Species | Host Organ | Incidence (%) |
|---|---|---|---|---|
| Callirhytis congregata | Cynipidae | Q. agrifolia | Catkin | 83% |
| Eriophyes mackiei | Eriophyidae | Q. agrifolia | Leaf | 83% |
| Callirhytis flora | Cynipidae | Q. agrifolia | Leaf | 83% |
| Callirhytis quercuspomiformis | Cynipidae | Q. agrifolia | Leaf | 17% |
| Callirhytis sp. | Cynipidae | Q. agrifolia | Stem | 33% |
| Heteroecus sp1 | Cynipidae | Q. agrifolia | Leaf | 50% |
| Unknown | – | Q. agrifolia | Leaf | 17% |
| Heteroecus sp2 | Cynipidae | Q. agrifolia | Leaf | 33% |
| Contarinia sp. | Cecidomyiidae | Q. agrifolia | Leaf | 33% |
| Callirhytes cf. perdens | Cynipidae | Q. agrifolia | Stem | 17% |
| Neuroterus fragilis | Cynipidae | Q. lobata | Leaf | 33% |
| Neuroterus saltatorius | Cynipidae | Q. lobata | Leaf | 83% |
| Andricus quercuscalifornicus | Cynipidae | Q. lobata | Stem | 33% |
| Disholcaspis eldoradensis | Cynipidae | Q. lobata | Stem | 33% |
| Antron douglasii | Cynipidae | Q. lobata and Q. douglasii | Leaf | 67% and 83% |
| Andricus crystallinus | Cynipidae | Q. douglasii | Leaf | 83% |
| Neuroterus saltatorius | Cynipidae | Q. douglasii | Leaf | 83% |
| Andricus gigas | Cynipidae | Q. douglasii | Leaf | 83% |
| Disholcaspis washingtonensis | Cynipidae | Q. douglasii | Stem | 67% |
| Liodora pattersonae | Cynipidae | Q. douglasii | Leaf | 33% |
| Besbiscus multipunctatus | Cynipidae | Q. douglasii | Leaf | 50% |
| Predictors | Relative Importance | Averaged Estimate | Standard Error | z-Value | p-Value |
|---|---|---|---|---|---|
| Flavonol Content | 0.68 | −0.2208919 | 0.1464645 | 1.506 | 0.132 |
| Chlorophyll | 0.51 | −0.1083698 | 0.3353858 | 0.323 | 0.747 |
| Anthocyanin Content | 0.50 | 0.1558205 | 0.1143836 | 1.360 | 0.174 |
| Nitrogen Balance Index | 0.50 | −0.1979788 | 0.3764416 | 0.525 | 0.599 |
| Specific Leaf Area | 0.43 | 0.1161805 | 0.0998047 | 1.162 | 0.245 |
| Water Content | 0.30 | −0.0457975 | 0.0928121 | 0.493 | 0.622 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perea, R.; Dirzo, R.; Bieler, S.; Wilson Fernandes, G. Incidence of Galls on Sympatric California Oaks: Ecological and Physiological Perspectives. Diversity 2021, 13, 20. https://doi.org/10.3390/d13010020
Perea R, Dirzo R, Bieler S, Wilson Fernandes G. Incidence of Galls on Sympatric California Oaks: Ecological and Physiological Perspectives. Diversity. 2021; 13(1):20. https://doi.org/10.3390/d13010020
Chicago/Turabian StylePerea, Ramón, Rodolfo Dirzo, Stephanie Bieler, and Geraldo Wilson Fernandes. 2021. "Incidence of Galls on Sympatric California Oaks: Ecological and Physiological Perspectives" Diversity 13, no. 1: 20. https://doi.org/10.3390/d13010020
APA StylePerea, R., Dirzo, R., Bieler, S., & Wilson Fernandes, G. (2021). Incidence of Galls on Sympatric California Oaks: Ecological and Physiological Perspectives. Diversity, 13(1), 20. https://doi.org/10.3390/d13010020