The Role of Carrion in the Landscapes of Fear and Disgust: A Review and Prospects


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
3. Results and Discussion
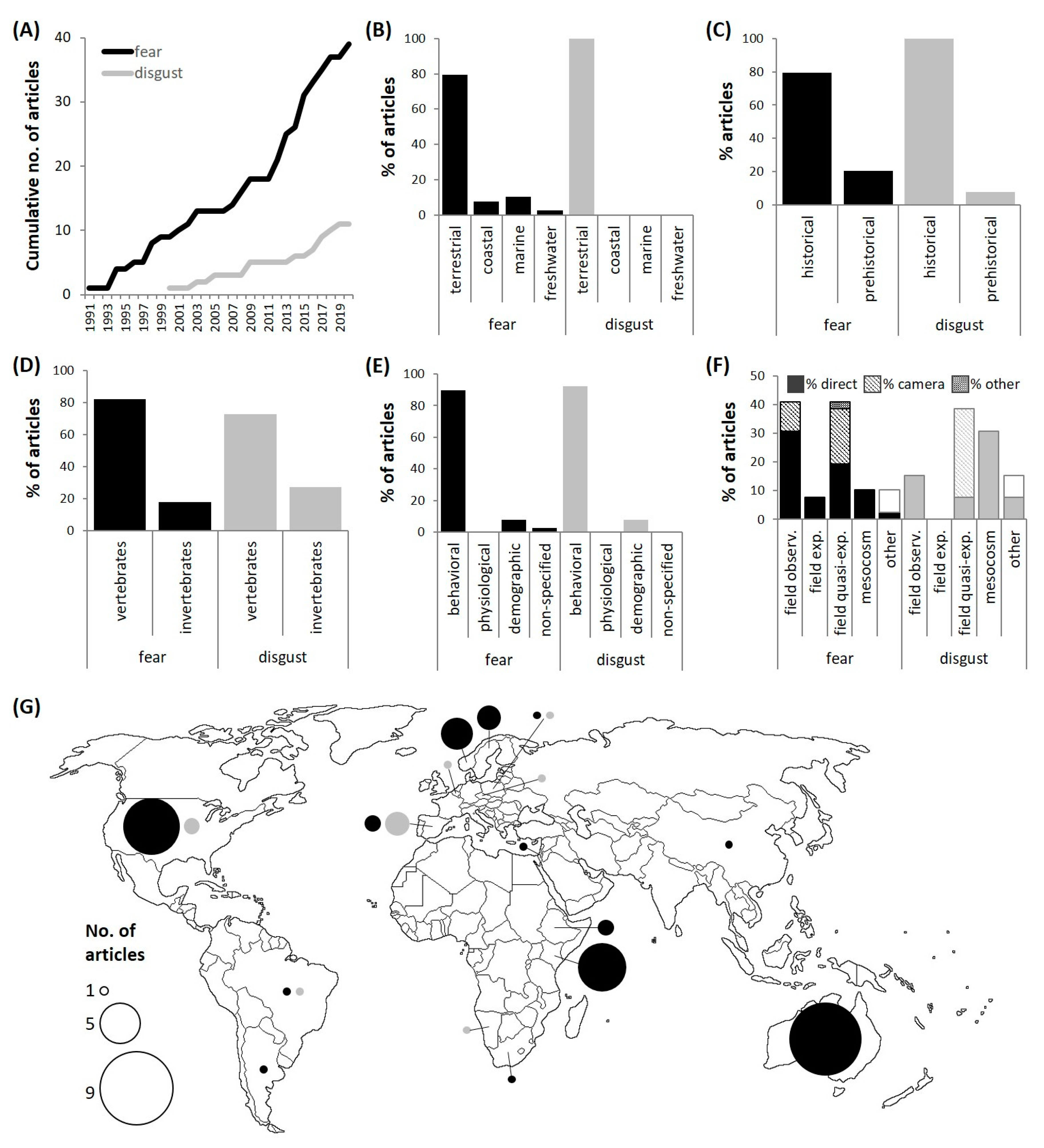
3.1. General Results
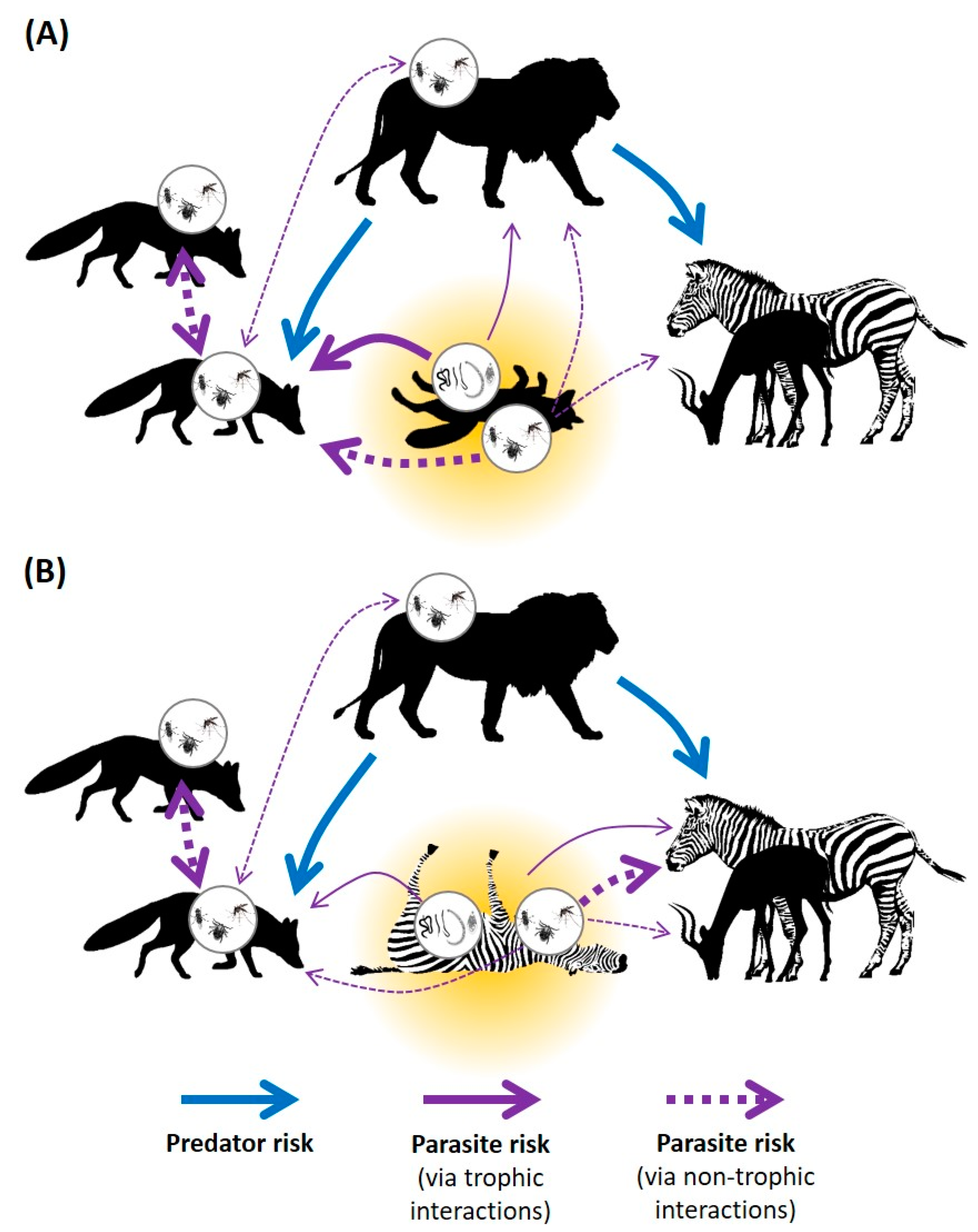
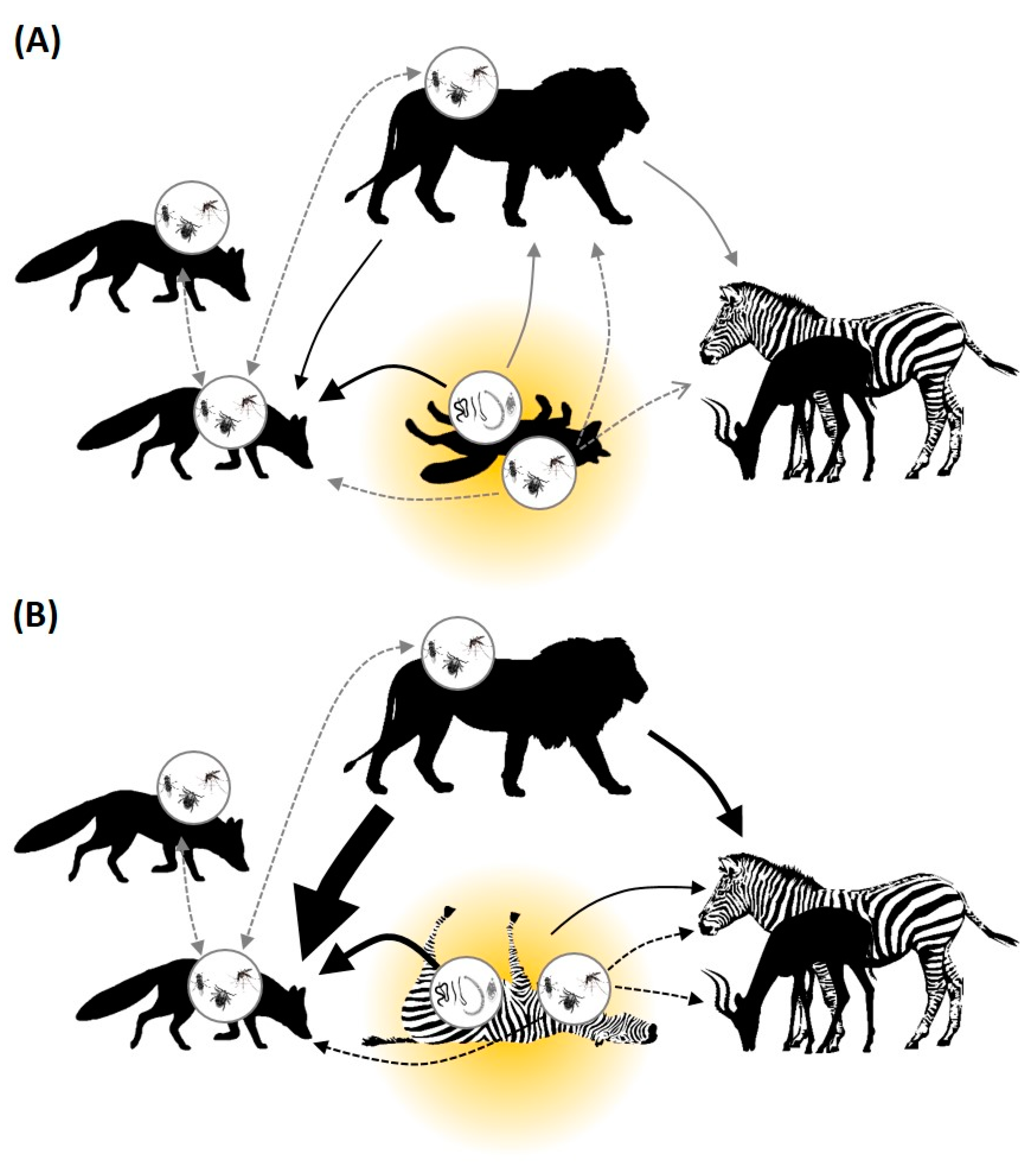
3.2. Interaction Pathways and Research Effort
3.3. Carrion Is Disgusting
3.4. Carrion Is Scary
3.5. Conclusions and Directions for Further Research
Author Contributions
Funding
Conflicts of Interest
Appendix A. References A1. Articles Reviewed. Articles Identified in the Google Scholar Search Only Are Marked with an Asterisk
- *Allen, M.L.; Elbroch, L.M.; Wilmers, C.C.; Witmer, H.U. The comparative effects of large carnivores on the acquisition of carrion by scavengers. Am. Nat. 2015, 185, 822–833.
- Allen, M.L.; Wilmers, C.C.; Elbroch, L.M.; Golla, J.M.; Witmer, H.U. The importance of motivation, weapons, and foul odors in driving encounter competition in carnivores. Ecology 2016, 97, 1905–1912.
- *Archer, M.S.; Elgar, M.A. Effects of decomposition on carcass attendance in a guild of carrion-breeding flies. Med. Vet. Entomol. 2003, 17, 263–271.
- *Archer, M.S.; Elgar, M.A. Female breeding-site preferences and larval feeding strategies of carrion-breeding Calliphoridae and Sarcophagidae (Diptera): A quantitative analysis. Aust. J. Zool. 2003, 51, 165–174.
- Atwood, T.C.; Gese, E.M. Coyotes and recolonizing wolves: Social rank mediates risk-conditional behaviour at ungulate carcasses. Anim. Behav. 2008, 75, 753–762.
- *Blanco, G.; Cardells, J.; Garijo-Toledo, M.M. Supplementary feeding and endoparasites in threatened avian scavengers: Coprologic evidence from red kites in their wintering stronghold. Environ. Res. 2017, 155, 22–30.
- Blumenschine, R.J. Hominid carnivory and foraging strategies, and the socio-economic function of early archaeological sites. Philos. Trans. R. Soc. Lond. B 1991, 334, 211–221.
- *Blumenschine, R.J.; Cavallo, J.A.; Capaldo, S.D. Competition for carcasses and early hominid behavioral ecology: A case study and conceptual framework. J. Hum. Evol. 1994, 27, 197–213.
- Blumenschine, R.J.; Stanistreet, I.G.; Njau, J.K.; Bamford, M.K.; Masao, F.T.; Albert, R.M.; Stollhofen, H.; Andrews, P.; Prassack, K.A.; McHenry, L.J.; et al. Environments and hominin activities across the FLK Peninsula during Zinjanthropus times (1.84 Ma), Olduvai Gorge, Tanzania. J. Hum. Evol. 2012, 63, 364–383.
- *Cortés-Avizanda, A.; Selva, N.; Carrete, M.; Donázar, J.A. Effects of carrion resources on herbivore spatial distribution are mediated by facultative scavengers. Basic Appl. Ecol. 2009, 10, 265–272.
- Cortés-Avizanda, A.; Carrete, M.; Serrano, D.; Donázar, J.A. Carcasses increase the probability of predation of ground-nesting birds: A caveat regarding the conservation value of vulture restaurants. Anim. Conserv. 2009, 12, 85–88.
- Cunningham, C.X.; Johnson, C.N.; Barmuta, L.A.; Hollings, T.; Woehler, E.J.; Jones, M.E. Top carnivore decline has cascading effects on scavengers and carrion persistence. Proc. R. Soc. B 2018, 285, 20181582.
- Daleo, P.; Alberti, J.; Avaca, M.S.; Narvarte, M.; Martinetto, M.; Iribarne, O. Avoidance of feeding opportunities by the whelk Buccinanops globulosum in the presence of damaged conspecifics. Mar. Biol. 2012, 159, 2359–2365.
- *DeVault, T.L.; Rhodes, O.E., Jr. Identification of vertebrate scavengers of small mammal carcasses in a forested landscape. Acta Theriol. 2002, 47, 185–192.
- Fialho, V.S.; Rodrigues, V.B.; Elliot, S.L. Nesting strategies and disease risk in necrophagous beetles. Ecol. Evol. 2018, 8, 3296–3310.
- Fodrie, F.J.; Brodeur, M.C.; Toscano, B.J.; Powers, S.P. Friend or foe: Conflicting demands and conditional risk taking by opportunistic scavengers. J. Exp. Mar. Biol. Ecol. 2012, 422–423, 114–121.
- *Foltan, P.; Puza, V. To complete their life cycle, pathogenic nematode-bacteria complexes deter scavengers from feeding on their host cadaver. Behav. Process. 2009, 80, 76–79.
- Frank, S.C.; Blaalid, R.; Mayer, M.; Zedrosser, A.; Steyaert, S.M.J.G. Fear the reaper: Ungulate carcasses may generate an ephemeral landscape of fear for rodents. R. Soc. Open Sci. 2020, 7, 191644.
- García-García, F.J.; Reyes-Martínez, M.J.; Ruiz-Delgado, M.C.; Sánchez-Moyano, J.E.; Castro-Casas, M.; Pérez-Hurtado, A. Does the gathering of shellfish affect the behavior of gastropod scavengers on sandy beaches? A field experiment. J. Exp. Mar. Biol. Ecol. 2015, 467, 1–6.
- Halley, D.J. Interspecific dominance and risk-taking in three species of corvid scavenger. J. Yamashina Inst. Ornithol. 2001, 33, 44–50.
- *Hunter, J.S.; Durant, S.M.; Caro, T.M. To flee or not to flee: Predator avoidance by cheetahs at kills. Behav. Ecol. Sociobiol. 2007, 61, 1033–1042.
- *Jennelle, C.S.; Samuel, M.D.; Nolden, C.A.; Berkley, E.A. Deer carcass decomposition and potential scavenger exposure to chronic wasting disease. J. Wildl. Manage. 2009, 73, 655–662.
- Jones, M.E. The function of vigilance in sympatric marsupial carnivores: The eastern quoll and the Tasmanian devil. Anim. Behav. 1998, 56, 1279–1284.
- *Lev-Yadun, S.; Gutman, M. Carrion odor and cattle grazing. Comm. Integr. Biol. 2013, 6, e26111.
- *Lupo, K.D. Experimentally derived extraction rates for marrow: Implications for body part exploitation strategies of Plio-Pleistocene hominid scavengers. J. Archaeol. Sci. 1998, 25, 657–675.
- *Mattisson, J.; Rauset, G.R.; Odden, J.; Andrén, H.; Linnell, J.D.C.; Persson, J. Predation or scavenging? Prey body condition influences decision-making in a facultative predator, the wolverine. Ecosphere 2016, 7, e01407.
- McKillup, S.C.; McKillup, R.V. The decision to feed by a scavenger in relation to the risks of predation and starvation. Oecologia 1994, 97, 41–48.
- *Moleón, M.; Sánchez-Zapata, J.A.; Sebastián-González, E.; Owen-Smith, N. Carcass size shapes the structure and functioning of an African scavenging assemblage. Oikos 2015, 124, 1391–1403.
- Moleón, M.; Martínez-Carrasco, C.; Muellerklein, O.C.; Getz, W.M.; Muñoz-Lozano, C.; Sánchez-Zapata, J.A. Carnivore carcasses are avoided by carnivores. J. Anim. Ecol. 2017, 86, 1179–1191.
- Monahan, C.M. Tha Hadza carcass transport debate revisited and its archaeological implications. J. Archaeol. Sci. 1998, 25, 405–424.
- Morton, B.; Chan, K. Hunger rapidly overrides the risk of predation in the subtidal scavenger Nassarius siquijorensis (Gastropoda: Nassariidae): An energy budget and a comparison with the intertidal Nassarius festivus in Hong Kong. J. Exp. Mar. Biol. Ecol. 1999, 240, 213–228.
- *Muñoz-Lozano, C.; Martín-Vega, D.; Martínez-Carrasco, C.; Sánchez-Zapata, J.A.; Morales-Reyes, Z.; Gonzálvez, M.; Moleón, M. Avoidance of carnivore carcasses by vertebrate scavengers enables colonization by a diverse community of carrion insects. PLoS ONE 2019, 14, e0221890.
- *Navarro, F.K.S.P.; Rezende, R. de S.; Gonçalves, J.F., Jr. Experimental assessment of temperature increase and presence of predator carcass changing the response of invertebrate shredders. Biota Neotrop. 2013, 13.
- Oliver, J.S. Estimates of hominid and carnivore involvement in the FLK Zinjanthropus fossil assemblage: Some socioecological implications. J. Hum. Evol. 1994, 27, 267–294.
- *Olson, Z.H.; Beasley, J.C.; Rhodes, O.E., Jr. Carcass type affects local scavenger guilds more than habitat connectivity. PLoS ONE 2016, 11, e0147798.
- O’Malley, C.; Elbroch, L.M.; Lendrum, P.E.; Quigley, H. Motion-triggered video cameras reveal spatial and temporal patterns of red fox foraging on carrion provided by mountain lions. PeerJ 2018, 6, e5324.
- *Pereira, H.; Detrain, C. Pathogen avoidance and prey discrimination in ants. R. Soc. Open Sci. 2020, 7, 191705.
- *Ragir, S.; Rosenberg, M.; Tierno, P. Gut morphology and the avoidance of carrion among chimpanzees, baboons, and early hominids. J. Anthropol. Res. 2000, 56, 477–512.
- *Rees, J.D.; Webb, J.K.; Crowther, M.S.; Letnic, M. Carrion subsidies provided by fishermen increase predation of beach-nesting bird nests by facultative scavengers. Anim. Conserv. 2015, 18, 44–49.
- Rees, J.D., Crowther, M.S., Kingsford, R.T., Letnic, M. Direct and indirect effects of carrion subsidies in an arid rangeland: Carrion has positive effects on facultative scavengers and negative effects on a small songbird. J. Arid Environ. 2020, 179, 104174.
- Rose, L.; Marshall, F. Meat eating, hominid sociality, and home bases revisited. Curr. Anthropol. 1996, 37, 307–338.
- *Schlacher, T.A.; Strydom, S.; Connolly, R.M. Multiple scavengers respond rapidly to pulsed carrion resources at the land-ocean interface. Acta Oecol. 2013, 48, 7–12.
- Schlacher, T.A.; Weston, M.A.; Lynn, D.; Schoeman, D.S.; Huijbers, C.M.; Olds, A.D.; Masters, S.; Connolly, R.M. Conservation gone to the dogs: When canids rule the beach in small coastal reserves. Biodivers. Conserv. 2015, 24, 493–509.
- *Selva, N.; Jędrzejewska, B.; Jędrzejewski, W.; Wajrak, A. Factors affecting carcass use by a guild of scavengers in European temperate woodland. Can. J. Zool. 2005, 83, 1590–1601.
- *Siva-Jothy, J.A.; Monteith, K.M.; Vale, P.F. Navigating infection risk during oviposition and cannibalistic foraging in a holometabolous insect. Behav. Ecol. 2018, 29, 1426–1435.
- Steinbeiser, C.M.; Wawrzynowski, C.A.; Ramos, X.; Olson, Z.H. Scavenging and the ecology of fear: Do animal carcasses create islands of risk on the landscape? Can. J. Zool. 2017, 96, 229–236.
- *Tranter, C.; LeFevre, L.; Evison, S.E.F.; Hughes, W.O.H. Threat detection: Contextual recognition and response to parasites by ants. Behav. Ecol. 2015, 26, 396–405.
- Turner, W.C.; Kausrud, K.L.; Krishnappa, Y.S.; Cromsigt, J.P.G.M.; Ganz, H.H.; Mapaure, I.; Cloete, C.C.; Havarua, Z.; Küsters, M.; Getz, W.M.; et al. Fatal attraction: Vegetation responses to nutrient inputs attract herbivores to infectious anthrax carcass sites. Proc. R. Soc. B 2014, 281, 20141785.
- *van Dijk, J.; Andersen, T.; May, R.; Andersen, R.; Andersen, R.; Landa, A. Foraging strategies of wolverines within a predator guild. Can. J. Zool. 2008, 86, 966–975.
- *Wikenros, C.; Sand, H.; Ahlqvist, P.; Liberg, O. Biomass flow and scavengers use of carcasses after re-colonization of an apex predator. PLoS ONE 2013, 8, e77373.
- Wikenros, C.; Ståhlberg, S.; Sand, H. Feeding under high risk of intraguild predation: Vigilance patterns of two medium-sized generalist predators. J. Mammal. 2014, 95, 862–870.
- Willems, E.P.; van Schaik, C.P. The social organization of Homo ergaster: Inferences from anti-predator responses in extant primates. J. Hum. Evol. 2017, 109, 11–21.
References
- Buck, J.C.; Weinstein, S.B.; Young, H.S. Ecological and evolutionary consequences of parasite avoidance. Trends Ecol. Evol. 2018, 33, 619–632. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, S.B.; Buck, J.C.; Young, H.S. A landscape of disgust. Science 2018, 359, 1213–1214. [Google Scholar] [CrossRef]
- Clinchy, M.; Sheriff, M.J.; Zanette, L.Y. Predator-induced stress and the ecology of fear. Funct. Ecol. 2013, 27, 56–65. [Google Scholar] [CrossRef]
- Sarabian, C.; Curtis, V.; McMullan, R. Evolution of pathogen and parasite avoidance behaviours. Phil. Trans. R. Soc. B 2018, 373, 20170256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunningham, C.X.; Johnson, C.N.; Barmuta, L.A.; Hollings, T.; Woehler, E.J.; Jones, M.E. Top carnivore decline has cascading effects on scavengers and carrion persistence. Proc. R. Soc. B 2018, 285, 20181582. [Google Scholar] [CrossRef] [Green Version]
- Doherty, J.-F.; Ruehle, B. An integrated landscape of fear and disgust: The evolution of avoidance behaviors amidst a myriad of natural enemies. Front. Ecol. Evol. 2020, 8, 564343. [Google Scholar] [CrossRef]
- Sebastián-González, E.; Barbosa, J.M.; Pérez-García, J.M.; Morales-Reyes, Z.; Botella, F.; Olea, P.; Mateo-Tomás, P.; Moleón, M.; Hiraldo, F.; Arrondo, E.; et al. Scavenging in the Anthropocene: Human impact drives vertebrate scavenger species richness at a global scale. Glob. Chang. Biol. 2019, 25, 3005–3017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, S.C.; Blaalid, R.; Mayer, M.; Zedrosser, A.; Steyaert, S.M.J.G. Fear the reaper: Ungulate carcasses may generate an ephemeral landscape of fear for rodents. R. Soc. Open Sci. 2020, 7, 191644. [Google Scholar] [CrossRef]
- Atwood, T.C.; Gese, E.M. Coyotes and recolonizing wolves: Social rank mediates risk-conditional behaviour at ungulate carcasses. Anim. Behav. 2008, 75, 753–762. [Google Scholar] [CrossRef]
- Blanco, G.; Cardells, J.; Garijo-Toledo, M.M. Supplementary feeding and endoparasites in threatened avian scavengers: Coprologic evidence from red kites in their wintering stronghold. Environ. Res. 2017, 155, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Turner, W.C.; Kausrud, K.L.; Krishnappa, Y.S.; Cromsigt, J.P.G.M.; Ganz, H.H.; Mapaure, I.; Cloete, C.C.; Havarua, Z.; Küsters, M.; Getz, W.M.; et al. Fatal attraction: Vegetation responses to nutrient inputs attract herbivores to infectious anthrax carcass sites. Proc. R. Soc. B 2014, 281, 20141785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ragir, S.; Rosenberg, M.; Tierno, P. Gut morphology and the avoidance of carrion among chimpanzees, baboons, and early hominids. J. Anthropol. Res. 2000, 56, 477–512. [Google Scholar] [CrossRef]
- Blumenschine, R.J.; Stanistreet, I.G.; Njau, J.K.; Bamford, M.K.; Masao, F.T.; Albert, R.M.; Stollhofen, H.; Andrews, P.; Prassack, K.A.; McHenry, L.J.; et al. Environments and hominin activities across the FLK Peninsula during Zinjanthropus times (1.84 Ma), Olduvai Gorge, Tanzania. J. Hum. Evol. 2012, 63, 364–383. [Google Scholar] [CrossRef] [PubMed]
- Moleón, M.; Sánchez-Zapata, J.A.; Margalida, A.; Carrete, M.; Owen-Smith, N.; Donázar, J.A. Humans and scavengers: The evolution of interactions and ecosystem services. BioScience 2014, 64, 394–403. [Google Scholar] [CrossRef] [Green Version]
- Lev-Yadun, S.; Ne’eman, G.; Shanas, U. A sheep in wolf’s clothing: Do carrion and dung odours of flowers not only attract pollinators but also deter herbivores? BioEssays 2009, 31, 84–88. [Google Scholar] [CrossRef]
- Evans, T.S.; Shi, Z.; Boots, M.; Liu, W.; Olival, K.J.; Xiao, X.; Vandewoude, S.; Brown, H.; Chen, J.L.; Civitello, D.J.; et al. Synergistic China-US ecological research is essential for global emerging infectious disease preparedness. EcoHealth 2020, 17, 160–173. [Google Scholar] [CrossRef] [Green Version]
- Getz, W.M. Biomass transformation webs provide a unified approach to consumer-resource modelling. Ecol. Lett. 2011, 14, 113–124. [Google Scholar] [CrossRef] [Green Version]
- Behringer, D.C.; Karvonen, A.; Bojko, J. Parasite avoidance behaviours in aquatic environments. Philos. Trans. R. Soc. B 2018, 373, 20170202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haddaway, N.R.; Woodcock, P.; Macura, B.; Collins, A. Making literature reviews more reliable through application of lessons from systematic reviews. Conserv. Biol. 2015, 29, 1596–1605. [Google Scholar] [CrossRef]
- Lozano, J.; Olszańska, A.; Morales-Reyes, Z.; Castro, A.A.; Malo, A.F.; Moleón, M.; Sánchez-Zapata, J.A.; Cortés-Avizanda, A.; von Wehrden, H.; Dorresteijn, I.; et al. Human-carnivore relations: A systematic review. Biol. Conserv. 2019, 237, 480–492. [Google Scholar] [CrossRef]
- Lima, S.L.; Dill, L.M. Behavioral decisions made under the risk of predation: A review and prospectus. Can. J. Zool. 1990, 68, 619–640. [Google Scholar] [CrossRef]
- Rohr, J.R.; Swan, A.; Raffel, T.R.; Hudson, P.J. Parasites, info-disruption, and the ecology of fear. Oecologia 2009, 159, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Blumstein, D.T.; Rangchi, T.N.; Briggs, T.; Souza de Andrade, F.; Natterson-Horowitz, B. A systematic review of carrion eater’s adaptations to avoid sickness. J. Wildl. Dis. 2017, 53, 577–581. [Google Scholar] [CrossRef] [PubMed]
- Baruzzi, C.; Mason, D.; Barton, B.; Lashley, M. Effects of increasing carrion biomass on food webs. Food Webs 2018, 16, e00096. [Google Scholar] [CrossRef]
- Buck, J.C. Indirect effects explain the role of parasites in ecosystems. Trends Parasitol. 2019, 35, 835–847. [Google Scholar] [CrossRef]
- Case, T.I.; Stevenson, R.J.; Byrne, R.W.; Hobaiter, C. The animal origins of disgust: Reports of basic disgust in nonhuman great apes. Evol. Behav. Sci. 2020, 14, 231–260. [Google Scholar] [CrossRef] [Green Version]
- Prugh, L.R.; Sivy, K.J. Enemies with benefits: Integrating positive and negative interactions among terrestrial carnivores. Ecol. Lett. 2020, 23, 902–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moleón, M.; Selva, N.; Sánchez-Zapata, J.A. The components and spatiotemporal dimensions of carrion biomass quantification. Trends Ecol. Evol. 2020, 35, 91–92. [Google Scholar] [CrossRef]
- Huang, S.; Bininda-Emonds, O.R.; Stephens, O.R.; Gittleman, J.L.; Altizer, S. Phylogenetically related and ecologically similar carnivores harbor similar parasite assemblages. J. Anim. Ecol. 2014, 83, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Stephens, P.R.; Altizer, S.; Smith, K.F.; Aguirre, A.A.; Brown, J.H.; Budischak, S.A.; Byers, J.E.; Dallas, T.A.; Davies, T.J.; Drake, J.M.; et al. The macroecology of infectious diseases: A new perspective on global-scale drivers of pathogen distributions and impacts. Ecol. Lett. 2016, 19, 1159–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moleón, M.; Sánchez-Zapata, J.A.; Sebastián-González, E.; Owen-Smith, N. Carcass size shapes the structure and functioning of an African scavenging assemblage. Oikos 2015, 124, 1391–1403. [Google Scholar] [CrossRef]
- Ogada, D.L.; Torchin, M.E.; Kinnaird, M.F.; Ezenwa, V.O. Effects of vulture declines on facultative scavengers and potential implications for mammalian disease transmission. Conserv. Biol. 2012, 26, 453–460. [Google Scholar] [CrossRef]
- Morales-Reyes, Z.; Sánchez-Zapata, J.A.; Sebastián-González, E.; Botella, F.; Carrete, M.; Moleón, M. Scavenging efficiency and red fox abundance in Mediterranean mountains with and without vultures. Acta Oecol. 2017, 79, 81–88. [Google Scholar] [CrossRef]
- Hill, J.E.; DeVault, T.L.; Beasley, J.C.; Rhodes, O.E., Jr.; Belant, J.L. Effects of vulture exclusion on carrion consumption by facultative scavengers. Ecol. Evol. 2018, 8, 2518–2526. [Google Scholar] [CrossRef] [PubMed]
- Selva, N.; Jędrzejewska, B.; Jędrzejewski, W.; Wajrak, A. Factors affecting carcass use by a guild of scavengers in European temperate woodland. Can. J. Zool. 2005, 83, 1590–1601. [Google Scholar] [CrossRef]
- Olson, Z.H.; Beasley, J.C.; Rhodes, O.E., Jr. Carcass type affects local scavenger guilds more than habitat connectivity. PLoS ONE 2016, 11, e0147798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moleón, M.; Martínez-Carrasco, C.; Muellerklein, O.C.; Getz, W.M.; Muñoz-Lozano, C.; Sánchez-Zapata, J.A. Carnivore carcasses are avoided by carnivores. J. Anim. Ecol. 2017, 86, 1179–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortés-Avizanda, A.; Selva, N.; Carrete, M.; Donázar, J.A. Effects of carrion resources on herbivore spatial distribution are mediated by facultative scavengers. Basic Appl. Ecol. 2009, 10, 265–272. [Google Scholar] [CrossRef]
- Cortés-Avizanda, A.; Carrete, M.; Serrano, D.; Donázar, J.A. Carcasses increase the probability of predation of ground-nesting birds: A caveat regarding the conservation value of vulture restaurants. Anim. Conserv. 2009, 12, 85–88. [Google Scholar] [CrossRef]
- Steinbeiser, C.M.; Wawrzynowski, C.A.; Ramos, X.; Olson, Z.H. Scavenging and the ecology of fear: Do animal carcasses create islands of risk on the landscape? Can. J. Zool. 2017, 96, 229–236. [Google Scholar] [CrossRef] [Green Version]
- McKillup, S.C.; McKillup, R.V. The decision to feed by a scavenger in relation to the risks of predation and starvation. Oecologia 1994, 97, 41–48. [Google Scholar] [CrossRef]
- Archer, M.S.; Elgar, M.A. Effects of decomposition on carcass attendance in a guild of carrion-breeding flies. Med. Vet. Entomol. 2003, 17, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, J.; Andersen, T.; May, R.; Andersen, R.; Andersen, R.; Landa, A. Foraging strategies of wolverines within a predator guild. Can. J. Zool. 2008, 86, 966–975. [Google Scholar] [CrossRef]
- Fodrie, F.J.; Brodeur, M.C.; Toscano, B.J.; Powers, S.P. Friend or foe: Conflicting demands and conditional risk taking by opportunistic scavengers. J. Exp. Mar. Biol. Ecol. 2012, 422–423, 114–121. [Google Scholar] [CrossRef]
- Schlacher, T.A.; Strydom, S.; Connolly, R.M. Multiple scavengers respond rapidly to pulsed carrion resources at the land-ocean interface. Acta Oecol. 2013, 48, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Wikenros, C.; Ståhlberg, S.; Sand, H. Feeding under high risk of intraguild predation: Vigilance patterns of two medium-sized generalist predators. J. Mammal. 2014, 95, 862–870. [Google Scholar] [CrossRef] [Green Version]
- Allen, M.L.; Elbroch, L.M.; Wilmers, C.C.; Witmer, H.U. The comparative effects of large carnivores on the acquisition of carrion by scavengers. Am. Nat. 2015, 185, 822–833. [Google Scholar] [CrossRef] [Green Version]
- Blumenschine, R.J. Hominid carnivory and foraging strategies, and the socio-economic function of early archaeological sites. Philos. Trans. R. Soc. Lond. B 1991, 334, 211–221. [Google Scholar]
- Monahan, C.M. Tha Hadza carcass transport debate revisited and its archaeological implications. J. Archaeol. Sci. 1998, 25, 405–424. [Google Scholar] [CrossRef]
- Willems, E.P.; van Schaik, C.P. The social organization of Homo ergaster: Inferences from anti-predator responses in extant primates. J. Hum. Evol. 2017, 109, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Lupo, K.D. Experimentally derived extraction rates for marrow: Implications for body part exploitation strategies of Plio-Pleistocene hominid scavengers. J. Archaeol. Sci. 1998, 25, 657–675. [Google Scholar] [CrossRef]
- Foltan, P.; Puza, V. To complete their life cycle, pathogenic nematode-bacteria complexes deter scavengers from feeding on their host cadaver. Behav. Process. 2009, 80, 76–79. [Google Scholar] [CrossRef]
- Jennelle, C.S.; Samuel, M.D.; Nolden, C.A.; Berkley, E.A. Deer carcass decomposition and potential scavenger exposure to chronic wasting disease. J. Wildl. Manag. 2009, 73, 655–662. [Google Scholar] [CrossRef]
- Fialho, V.S.; Rodrigues, V.B.; Elliot, S.L. Nesting strategies and disease risk in necrophagous beetles. Ecol. Evol. 2018, 8, 3296–3310. [Google Scholar] [CrossRef]
- Muñoz-Lozano, C.; Martín-Vega, D.; Martínez-Carrasco, C.; Sánchez-Zapata, J.A.; Morales-Reyes, Z.; Gonzálvez, M.; Moleón, M. Avoidance of carnivore carcasses by vertebrate scavengers enables colonization by a diverse community of carrion insects. PLoS ONE 2019, 14, e0221890. [Google Scholar] [CrossRef] [PubMed]
- Lev-Yadun, S.; Gutman, M. Carrion odor and cattle grazing. Comm. Integr. Biol. 2013, 6, e26111. [Google Scholar] [CrossRef] [PubMed]
- Lepczyk, C.A.; Lohr, C.A.; Duffy, D.C. A review of cat behavior in relation to disease risk and management options. Appl. Anim. Behav. Sci. 2015, 173, 29–39. [Google Scholar] [CrossRef]
- Mateo-Tomás, P.; Olea, P.P.; Moleón, M.; Selva, N.; Sánchez-Zapata, J.A. Both rare and common species support ecosystem services in scavenging communities. Glob. Ecol. Biogeogr. 2017, 26, 1459–1470. [Google Scholar] [CrossRef]
- Rossi, L.; Interisano, M.; Deksne, G.; Pozio, E. The subnivium, a haven for Trichinella larvae in host carcasses. Int. J. Parasitol. 2019, 8, 229–233. [Google Scholar] [CrossRef]
- Margalida, A.; Moleón, M. Toward carrion-free ecosystems? Front. Ecol. Environ. 2016, 14, 183–184. [Google Scholar] [CrossRef] [Green Version]
- Morales-Reyes, Z.; Pérez-García, J.M.; Moleón, M.; Botella, F.; Carrete, M.; Donázar, J.A.; Cortés-Avizanda, A.; Arrondo, E.; Moreno-Opo, R.; Jiménez, J.; et al. Evaluation of the network of protection areas for the feeding of scavengers in Spain: From biodiversity conservation to greenhouse gas emission savings. J. Appl. Ecol. 2017, 54, 1120–1129. [Google Scholar] [CrossRef]
- Rees, J.D.; Webb, J.K.; Crowther, M.S.; Letnic, M. Carrion subsidies provided by fishermen increase predation of beach-nesting bird nests by facultative scavengers. Anim. Conserv. 2015, 18, 44–49. [Google Scholar] [CrossRef] [Green Version]
- DeVault, T.L.; Rhodes, O.E., Jr.; Shivik, J.A. Scavenging by vertebrates: Behavioral, ecological, and evolutionary perspectives on an important energy transfer pathway in terrestrial ecosystems. Oikos 2003, 102, 225–234. [Google Scholar] [CrossRef] [Green Version]
- Moleón, M.; Sánchez-Zapata, J.A.; Selva, N.; Donázar, J.A.; Owen-Smith, N. Inter-specific interactions linking predation and scavenging in terrestrial vertebrate assemblages. Biol. Rev. 2014, 89, 1042–1054. [Google Scholar] [CrossRef]
- Morton, B.; Chan, K. Hunger rapidly overrides the risk of predation in the subtidal scavenger Nassarius siquijorensis (Gastropoda: Nassariidae): An energy budget and a comparison with the intertidal Nassarius festivus in Hong Kong. J. Exp. Mar. Biol. Ecol. 1999, 240, 213–228. [Google Scholar] [CrossRef]
- Pereira, L.M.; Owen-Smith, N.; Moleón, M. Facultative predation and scavenging by mammalian carnivores: Seasonal, regional and intra-guild comparisons. Mammal Rev. 2014, 44, 44–45. [Google Scholar] [CrossRef]
- Hunter, J.S.; Durant, S.M.; Caro, T.M. To flee or not to flee: Predator avoidance by cheetahs at kills. Behav. Ecol. Sociobiol. 2007, 61, 1033–1042. [Google Scholar] [CrossRef]
- Amorós, M.; Gil-Sánchez, J.M.; López-Pastor, B.D.L.N.; Moleón, M. Hyaenas and lions: How the largest African carnivores interact at carcasses. Oikos 2020, 129, 1820–1832. [Google Scholar] [CrossRef]
- Daleo, P.; Alberti, J.; Avaca, M.S.; Narvarte, M.; Martinetto, M.; Iribarne, O. Avoidance of feeding opportunities by the whelk Buccinanops globulosum in the presence of damaged conspecifics. Mar. Biol. 2012, 159, 2359–2365. [Google Scholar] [CrossRef]
- Koprivnikar, J.; Penalva, L. Lesser of two evils? Foraging choices in response to threats of predation and parasitism. PLoS ONE 2015, 10, e0116569. [Google Scholar] [CrossRef] [PubMed]
- Parsons, M.H.; Blumstein, D.T. Familiarity breeds contempt: Kangaroos persistently avoid areas with experimentally deployed dingo scents. PLoS ONE 2010, 5, e10403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barton, P.S.; Cunningham, S.A.; Lindenmayer, D.B.; Manning, A.D. The role of carrion in maintaining biodiversity and ecological processes in terrestrial ecosystems. Oecologia 2013, 171, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Combes, C. Parasitism: The Ecology and Evolution of Intimate Interactions; The University of Chicago Press: Chicago, IL, USA, 2001. [Google Scholar]
- Altizer, S.; Dobson, A.; Hosseini, P.; Hudson, P.; Pascual, M.; Rohani, P. Seasonality and the dynamics of infectious diseases. Ecol. Lett. 2006, 9, 467–484. [Google Scholar] [CrossRef] [Green Version]
- Costard, S.; Wieland, B.; de Glanville, W.; Jori, F.; Rowlands, R.; Vosloo, W.; Roger, F.; Pfeiffer, D.U.; Dixon, L.K. African swine fever: How can global spread be prevented? Philos. Trans. R. Soc. B 2009, 364, 2683–2696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barton, P.S.; Evans, M.J.; Foster, C.N.; Pechal, J.L.; Bump, J.K.; Quaggioto, M.-M.; Benbow, M.E. Towards quantifying carrion biomass in ecosystems. Trends Ecol. Evol. 2019, 34, 950–961. [Google Scholar] [CrossRef] [PubMed]
- Moleón, M.; Sánchez-Zapata, J.A. Non-trophic functions of carcasses: From death to the nest. Front. Ecol. Environ. 2016, 14, 340–341. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moleón, M.; Sánchez-Zapata, J.A. The Role of Carrion in the Landscapes of Fear and Disgust: A Review and Prospects. Diversity 2021, 13, 28. https://doi.org/10.3390/d13010028
Moleón M, Sánchez-Zapata JA. The Role of Carrion in the Landscapes of Fear and Disgust: A Review and Prospects. Diversity. 2021; 13(1):28. https://doi.org/10.3390/d13010028
Chicago/Turabian StyleMoleón, Marcos, and José A. Sánchez-Zapata. 2021. "The Role of Carrion in the Landscapes of Fear and Disgust: A Review and Prospects" Diversity 13, no. 1: 28. https://doi.org/10.3390/d13010028
APA StyleMoleón, M., & Sánchez-Zapata, J. A. (2021). The Role of Carrion in the Landscapes of Fear and Disgust: A Review and Prospects. Diversity, 13(1), 28. https://doi.org/10.3390/d13010028