Occupancy of the American Three-Toed Woodpecker in a Heavily-Managed Boreal Forest of Eastern Canada



Abstract
:1. Introduction
2. Materials and Methods
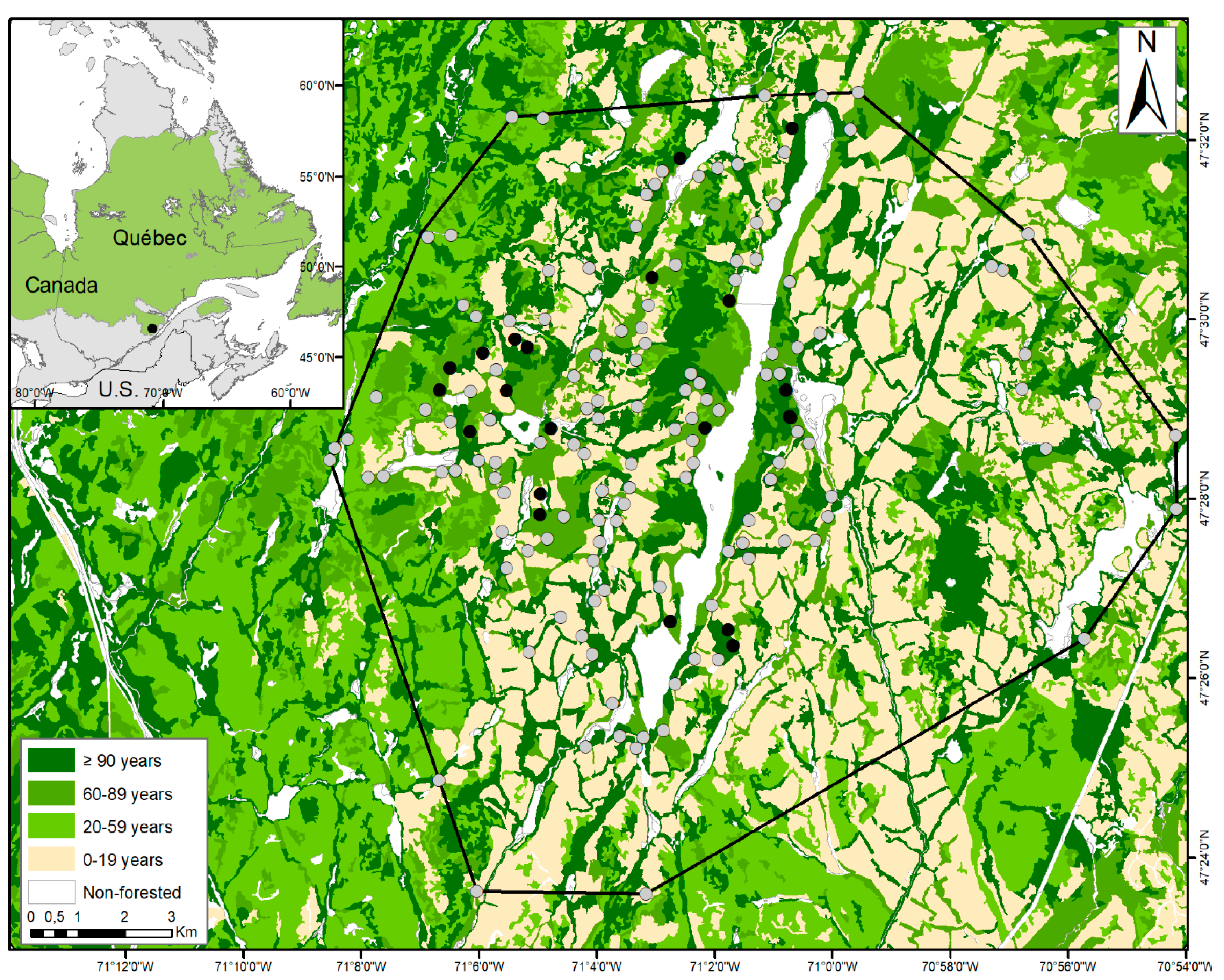
2.1. Study Area
2.2. Woodpecker Surveys
2.3. Habitat Characteristics
2.4. Occupancy Analyses
3. Results
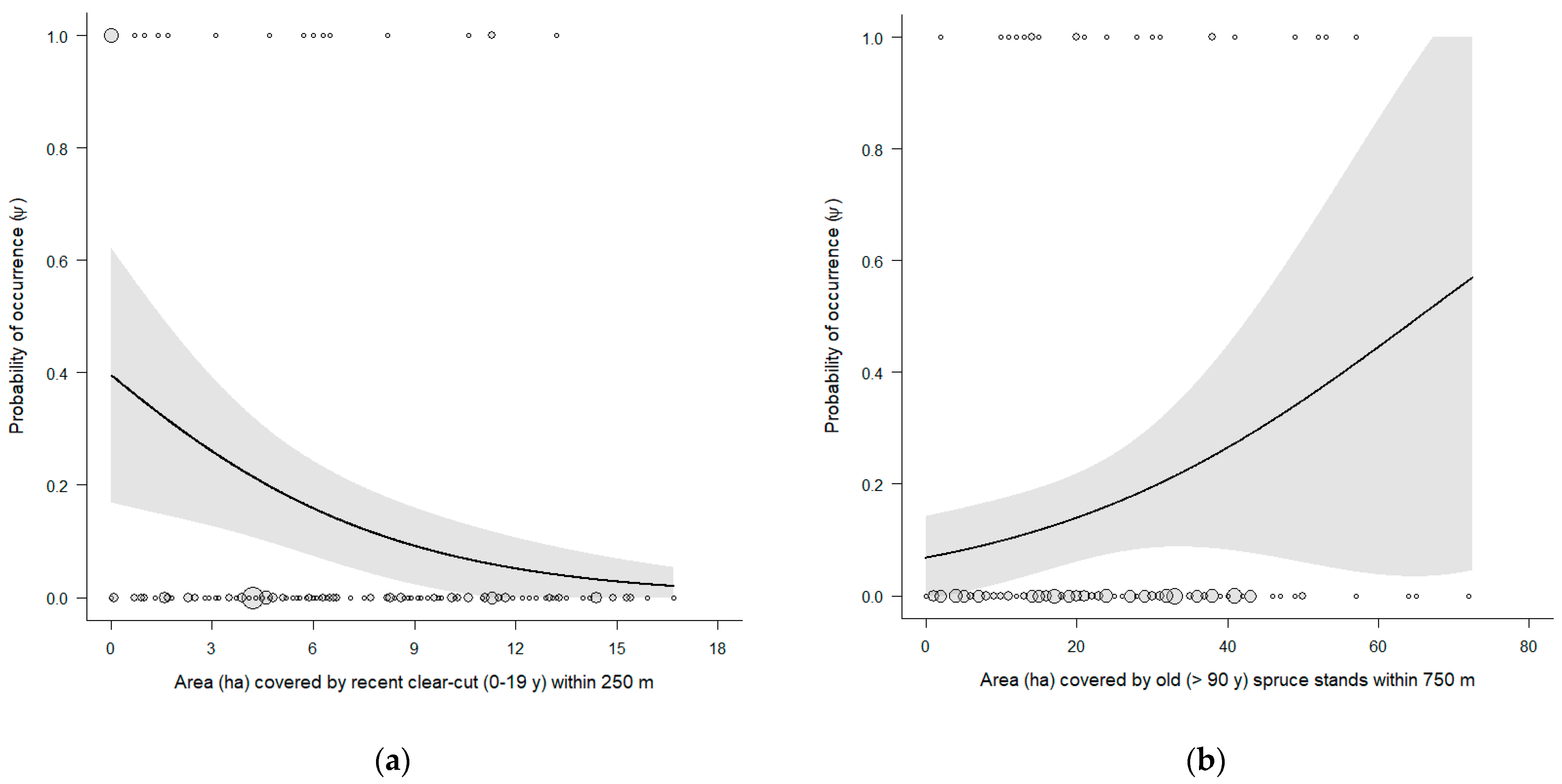
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Candidate Model | k | AICc | ΔAICc | ω | LL |
|---|---|---|---|---|---|
| ~ p(time) | 3 | 185.05 | 0.00 | 0.42 | −89.46 |
| ~ p(year) | 3 | 186.66 | 1.61 | 0.19 | −90.27 |
| ~ p(null) | 2 | 186.82 | 1.77 | 0.17 | −91.38 |
| ~ p(date) | 3 | 187.00 | 1.95 | 0.16 | −90.44 |
| ~ p(wind) | 3 | 188.60 | 3.55 | 0.07 | −91.23 |
| mean_age_250 | clear_cut_250 | old_spruce_250 | mean_age_750 | sd_age_750 | old_spruce_750 | |
|---|---|---|---|---|---|---|
| mean_age_250 | 1 | |||||
| clear_cut_250 | −0.53 | 1 | ||||
| old_spruce_250 | 0.42 | 0.03 | 1 | |||
| mean_age_750 | 0.66 | −0.36 | 0.19 | 1 | ||
| sd_age_750 | 0.19 | 0.31 | 0.32 | 0.28 | 1 | |
| old_spruce_750 | 0.29 | 0.17 | 0.60 | 0.37 | 0.23 | 1 |
References
- Brandt, J.P.; Flannigan, M.D.; Maynard, D.G.; Thompson, I.D.; Volney, W.J.A. An Introduction to Canada’s Boreal Zone: Ecosystem Processes, Health, Sustainability, and Environmental Issues. Environ. Rev. 2013, 21, 207–226. [Google Scholar] [CrossRef]
- Kneeshaw, D.D.; Bergeron, Y. Canopy Gap Characteristics and Tree Replacement in the Southeastern Boreal Forest. Ecology 1998, 79, 783–794. [Google Scholar] [CrossRef]
- Bergeron, Y.; Harvey, B.; Leduc, A.; Gauthier, S. Forest Management Guidelines Based on Natural Disturbance Dynamics: Stand- and Forest-Level Considerations. For. Chron. 1999, 75, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Pasher, J.; Seed, E.; Duffe, J. Development of Boreal Ecosystem Anthropogenic Disturbance Layers for Canada Based on 2008 to 2010 Landsat Imagery. Can. J. Remote Sens. 2013, 39, 42–58. [Google Scholar] [CrossRef]
- Frank, D.; Finckh, M.; Wirth, C. Impacts of Land Use on Habitat Functions of Old-Growth Forests and their Biodiversity. In Old-Growth Forests. Ecological Studies (Analysis and Synthesis); Wirth, C., Gleixner, G., Heimann, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 207, pp. 429–450. ISBN 978-3-540-92705-1. [Google Scholar]
- Grondin, P.; Gauthier, S.; Poirier, V.; Tardif, P.; Boucher, Y.; Bergeron, Y. Have Some Landscapes in the Eastern Canadian Boreal Forest Moved beyond Their Natural Range of Variability? For. Ecosyst. 2018, 5, 30. [Google Scholar] [CrossRef] [Green Version]
- Mosseler, A.; Thompson, I.; Pendrel, B.A. Overview of Old-Growth Forests in Canada from a Science Perspective. Environ. Rev. 2003, 11, S1–S7. [Google Scholar] [CrossRef]
- Stokland, J.; Siitonen, J. Species diversity of saproxylic organisms. In Biodiversity in Dead Wood; Stokland, J., Siitonen, J., Jonsson, B., Eds.; Cambridge University Press: Cambridge, UK, 2012; pp. 248–274. ISBN 978-1-139-02584-3. [Google Scholar]
- Nilsson, S.G. Selecting biodiversity indicators to set conservation targets: Species, structures, or processes? In Setting Conservation Targets for Managed Forest Landscapes; Villard, M.-A., Jonsson, B.G., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 79–108. ISBN 978-0-521-87709-1. [Google Scholar]
- Boudreault, C.; Gauthier, S.; Bergeron, Y. Epiphytic Lichens and Bryophytes on Populus Tremuloides Along a Chronosequence in the Southwestern Boreal Forest of Québec, Canada. Bryologist 2000, 103, 725–738. [Google Scholar] [CrossRef]
- Bergeron, Y.; Fenton, N.J. Boreal Forests of Eastern Canada Revisited: Old Growth, Nonfire Disturbances, Forest Succession, and Biodiversity. Botany 2012, 90, 509–523. [Google Scholar] [CrossRef]
- Martin, M.; Tremblay, J.A.; Ibarzabal, J. An Indicator Species Highlights Continuous Deadwood Supply Is a Key Ecological Attribute of Boreal Old-Growth Forests. Ecosphere. under review.
- Wesołowski, T.; Martin, K. Tree Holes and Hole-Nesting Birds in European and North American Forests. In Ecology and Conservation of Forest Birds; Mikusiński, G., Roberge, J.-M., Fuller, R., Eds.; Cambridge University Press: Cambridge, UK, 2018; pp. 79–134. ISBN 978-1-139-68036-3. [Google Scholar]
- Drapeau, P.; Nappi, A.; Imbeau, L.; Saint-Germain, M. Standing Deadwood for Keystone Bird Species in the Eastern Boreal Forest: Managing for Snag Dynamics. For. Chron. 2009, 85, 227–234. [Google Scholar] [CrossRef] [Green Version]
- Tremblay, J.A.; Savard, J.-P.L.; Ibarzabal, J. Structural Retention Requirements for a Key Ecosystem Engineer in Conifer-Dominated Stands of a Boreal Managed Landscape in Eastern Canada. For. Ecol. Manag. 2015, 357, 220–227. [Google Scholar] [CrossRef]
- Imbeau, L.; Desrochers, A. Area Sensitivity and Edge Avoidance: The Case of the Three-Toed Woodpecker (Picoides Tridactylus) in a Managed Forest. For. Ecol. Manag. 2002, 249–256. [Google Scholar] [CrossRef]
- Nappi, A.; Drapeau, P.; Leduc, A. How Important Is Dead Wood for Woodpeckers Foraging in Eastern North American Boreal Forests? For. Ecol. Manag. 2015, 346, 10–21. [Google Scholar] [CrossRef]
- Cadieux, P.; Drapeau, P. Are Old Boreal Forests a Safe Bet for the Conservation of the Avifauna Associated with Decayed Wood in Eastern Canada? For. Ecol. Manag. 2017, 385, 127–139. [Google Scholar] [CrossRef]
- Imbeau, L.; Savard, J.-P.L.; Gagnon, R. Comparing Bird Assemblages in Successional Black Spruce Stands Originating from Fire and Logging. Can. J. Zool. 1999, 77, 1850–1860. [Google Scholar] [CrossRef]
- Imbeau, L.; Mönkkönen, M.; Desrochers, A. Long-Term Effects of Forestry on Birds of the Eastern Canadian Boreal Forests: A Comparison with Fennoscandia. Conserv. Biol. 2001, 1151–1162. [Google Scholar] [CrossRef]
- Craig, C.; Mazerolle, M.J.; Taylor, P.D.; Tremblay, J.A.; Villard, M.-A. Predictors of Habitat Use and Nesting Success for Two Sympatric Species of Boreal Woodpeckers in an Unburned, Managed Forest Landscape. For. Ecol. Manag. 2019, 438, 134–141. [Google Scholar] [CrossRef]
- Saucier, J.-P.; Robitaille, A.; Grondin, P.; Bergeron, J.-F.; Gosselin, J. Les Régions Écologiques Du Québec Méridional, 4th ed.; Carte à l’échelle de 1/1 250,000; Ministère Des Ressources Naturelles et de La Faune Du Québec: Quebec, QC, Canada, 2011.
- Saucier, J.-P.; Grondin, P.; Robitaille, A.; Gosselin, J.; Morneau, C.; Richard, P.J.H.; Brisson, J.; Sirois, L.; Leduc, A.; Morin, H.; et al. Écologie forestière. In Manuel de Foresterie—Nouvelle Édition Entièrement Revue et Augmentée; Ordre des ingénieurs forestiers du Québec, Ed.; Éditions Mulitmondes: Quebec, QC, Canada, 2009; pp. 165–316. [Google Scholar]
- Boucher, Y.; Grondin, P.; Noël, J.; Hotte, D.; Blouin, J.; Roy, G. Classification Des Écosystèmes et Caractérisation Des Forêts Mûres et Surannées: Le Cas Du Projet Pilote de La Réserve Faunique Des Laurentides; Gouvernement du Québec, Ministère des Ressources naturelles et de la Faune, Direction de la Recherche Forestière: Quebec, QC, Canada, 2008.
- Boucher, Y.; Bouchard, M.; Grondin, P.; Tardif, P. Le Registre Des États de Référence: Intégration Des Connaissances Sur La Structure, La Composition et La Dynamique Des Paysages Forestiers Naturels Du Québec Méridional; Gouvernement du Québec, Ministère des Ressources naturelles et de la Faune, Direction de la recherche forestière: Quebec, QC, Canada, 2011.
- Desrochers, A.; Drolet, B. Le Programme de Surveillance Des Oiseaux Nicheurs de La Forêt Montmorency: Une Nouvelle Source de Tendances Des Populations d’oiseaux Nicheurs Pour La Forêt Boréale Au Québec. Nat. Can. 2017, 141, 61–74. [Google Scholar] [CrossRef] [Green Version]
- Boucher, Y.; Grondin, P. Impact of Logging and Natural Stand-Replacing Disturbances on High-Elevation Boreal Landscape Dynamics (1950–2005) in Eastern Canada. For. Ecol. Manag. 2012, 263, 229–239. [Google Scholar] [CrossRef]
- Brandt, J.P. The Extent of the North American Boreal Zone. Environ. Rev. 2009, 17, 101–161. [Google Scholar] [CrossRef]
- Xeno-Canto. Available online: https://www.xeno-canto.org/ (accessed on 1 May 2016).
- Macaulay Library—The Cornell Lab of Ornithology. Available online: https://www.macaulaylibrary.org/ (accessed on 1 May 2016).
- Weather Archive in Fort Montmorency, METAR. Available online: https://rp5.ru/Weather_archive_in_Fort_Montmorency,_METAR (accessed on 15 August 2020).
- Ministère des Forêts, de la Faune et des Parcs. Norme de Stratification Écoforestière: Quatrième Inventaire Écoforestier du Québec méridional; Secteur des Forêts. Direction des Inventaires Forestiers: Québec, QC, Canada, 2015; ISBN 978-2-550-73857-2.
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Mackenzie, D.I.; Nichols, J.D.; Lachman, G.B.; Droege, S.; Royle, J.A.; Catherine, A. Langtimm Estimating Site Occupancy Rates When Detection Probabilities Are Less than One. Ecology 2002, 2248–2255. [Google Scholar] [CrossRef]
- Fiske, I.; Chandler, R. Unmarked: An R Package for Fitting Hierarchical Models of Wildlife Occurrence and Abundance. J. Stat. Soft. 2011, 43. [Google Scholar] [CrossRef] [Green Version]
- Clement, M.A.; Barrett, K.; Baldwin, R.F. Key Habitat Features Facilitate the Presence of Barred Owls in Developed Landscapes. ACE 2019, 14, art12. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multi-Model Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002; ISBN 0-387-95364-7. [Google Scholar]
- Mazerolle, M.J. AICcmodavg: Model Selection and Multimodel Inference Based on (Q)AIC(c). R Package Version 2.3-0. 2020. Available online: https://repo.bppt.go.id/cran/web/packages/AICcmodavg/vignettes/AICcmodavg.pdf (accessed on 20 May 2020).
- Schieck, J.; Song, S.J. Changes in Bird Communities throughout Succession Following Fire and Harvest in Boreal Forests of Western North America: Literature Review and Meta-Analyses. Can. J. For. Res. 2006, 36, 1299–1318. [Google Scholar] [CrossRef]
- Imbeau, L.; Desrochers, A. Foraging Ecology and Use of Drumming Trees by Three-Toed Woodpeckers. J. Wildl. Manag. 2002, 66, 222. [Google Scholar] [CrossRef]
- Gagné, C.; Imbeau, L.; Drapeau, P. Anthropogenic Edges: Their Influence on the American Three-Toed Woodpecker (Picoides dorsalis) Foraging Behaviour in Managed Boreal Forests of Quebec. For. Ecol. Manag. 2007, 252, 191–200. [Google Scholar] [CrossRef]
- Hagan, J.M.; McKinley, P.S.; Meehan, A.L.; Grove, S.L. Diversity and Abundance of Landbirds in a Northeastern Industrial Forest. J. Wildl. Manag. 1997, 61, 718. [Google Scholar] [CrossRef]
- Vaillancourt, M.-A.; Drapeau, P.; Gauthier, S.; Robert, M. Availability of Standing Trees for Large Cavity-Nesting Birds in the Eastern Boreal Forest of Québec, Canada. For. Ecol. Manag. 2008, 255, 2272–2285. [Google Scholar] [CrossRef]
- Tremblay, J.A.; Leonard, D.L., Jr.; Imbeau, L. American Three-toed Woodpecker (Picoides dorsalis). Version 1.0. In Birds of the World; Rodewald, P.G., Ed.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Environment Canada. Recovery Strategy for the Woodland Caribou, Boreal Population (Rangifer Tarandus Caribou) in Canada [Proposed]. Species at Risk Act Recovery Strategy Series; Environment Canada: Ottawa, ON, Canada, 2011; pp. vi + 55.
- Halme, P.; Allen, K.A.; Auniņš, A.; Bradshaw, R.H.W.; Brūmelis, G.; Čada, V.; Clear, J.L.; Eriksson, A.-M.; Hannon, G.; Hyvärinen, E.; et al. Challenges of Ecological Restoration: Lessons from Forests in Northern Europe. Biol. Conserv. 2013, 167, 248–256. [Google Scholar] [CrossRef]
- Shorohova, E.; Kneeshaw, D.; Kuuluvainen, T.; Gauthier, S. Variability and Dynamics of Old-Growth Forests in the Circumboreal Zone: Implications for Conservation, Restoration and Management. Silva Fenn. 2011, 45. [Google Scholar] [CrossRef] [Green Version]
- Tikkanen, O.-P.; Martikainen, P.; Hyvärinen, E.; Junninen, K.; Kouki, J. Red-Listed Boreal Forest Species of Finland: Associations with Forest Structure, Tree Species, and Decaying Wood. Ann. Zool. Fennici 2006, 43, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Tingstad, L.; Grytnes, J.A.; Felde, V.A.; Juslén, A.; Hyvärinen, E.; Dahlberg, A. The Potential to Use Documentation in National Red Lists to Characterize Red-Listed Forest Species in Fennoscandia and to Guide Conservation. Glob. Ecol. Conserv. 2018, 15, e00410. [Google Scholar] [CrossRef]
- Haavik, A.; Dale, S. Are Reserves Enough? Value of Protected Areas for Boreal Forest Birds in Southeastern Norway. Ann. Zool. Fenn. 2012, 49, 69–80. [Google Scholar] [CrossRef]
- Nappi, A. Sélection d’habitat et Démographie Du Pic à Dos Noir Dans Les Forêts Brûlées de La Forêt Boréale. Ph.D. Thesis, Université du Québec à Montréal, Montréal, QC, Canada, 2009. [Google Scholar]
- Fenton, N.J.; Imbeau, L.; Work, T.; Jacobs, J.; Bescond, H.; Drapeau, P.; Bergeron, Y. Lessons Learned from 12 Years of Ecological Research on Partial Cuts in Black Spruce Forests of Northwestern Québec. For. Chron. 2013, 89, 350–359. [Google Scholar] [CrossRef] [Green Version]
- Tremblay, J.A.; Bélanger, L.; Desponts, M.; Brunet, G. La Restauration Passive Des Sapinières Mixtes de Seconde Venue: Une Alternative Pour La Conservation Des Sapinières Mixtes Anciennes. Can. J. For. Res. 2007, 37, 825–839. [Google Scholar] [CrossRef]
- Desrochers, A. Tendances Ornithologiques du Québec. Available online: https://www.toq.ffgg.ulaval.ca/ (accessed on 13 October 2020).
- Meehan, T.D.; LeBaron, G.S.; Dale, K.; Michel, N.L.; Verutes, G.M.; Langham, G.M. Abundance Trends of Birds Wintering in the USA and Canada, from Audubon Christmas Bird Counts, 1966–2017, Version 2.1; National Audubon Society: New York, NY, USA, 2018. [Google Scholar]



| Habitat Characteristic | Description | Not Detected (n = 163) | Detected (n = 22) | Random (n = 185) |
|---|---|---|---|---|
| clear_cut_250 | Area (ha) covered by recent (0–19 years) clear-cut | 6.58 (4.62) a | 3.91 (4.17) b | 6.60 (5.89) a |
| mean_age_250 | Mean forest age (year) | 70.78 (25.44) a | 83.27 (30.27) a | 64.92 (32.34) b |
| mean_age_750 | 70.07 (15.34) a | 76.27 (20.63) a | 64.33 (19.47) b | |
| young_250 | Area (ha) covered by young closed forest (20–59 years) | 1.29 (2.96) a | 0.86 (1.54) a,b | 3.24 (5.06) b |
| young_750 | 14.71 (24.20) a | 8.34 (10.62) a | 27.03 (35.24) b | |
| mature_250 | Area (ha) covered by mature forest (60–89 years) | 4.87 (4.64) a | 6.23 (5.72) a | 3.11 (3.80) b |
| mature_750 | 38.49 (21.80) a | 48.89 (23.05) a | 28.00 (22.32) b | |
| old_spruce_250 | Area (ha) covered by old forest (≥90 years) dominated by black spruce | 2.92 (3.04) a | 3.85 (3.46) a | 2.26 (3.51) b |
| old_spruce_750 | 21.87 (13.18) a | 26.00 (16.33) a | 19.92 (15.08) a | |
| sd_age_750 | Standard deviation of forest age | 56.71 (8.89) a | 53.08 (9.35) a | 54.62 (9.80) a |
| nf_750 | Area (ha) covered by non-forested lands in the landscape | 21.04 (19.98) a | 26.44 (23.93) a | 12.00 (16.16) b |
| Candidate Model | k | AICc | ΔAICc | ω | LL |
|---|---|---|---|---|---|
| ~ p(time) ~ ψ(clear_cut_250 + old_spruce_750) | 5 | 176.56 | 0.00 | 0.49 | −83.11 |
| ~ p(time) ~ ψ(clear_cut_250) | 4 | 178.46 | 1.90 | 0.19 | −85.12 |
| ~ p(time) ~ ψ(mean_age_250 + sd_age_750) | 5 | 178.99 | 2.43 | 0.15 | −84.33 |
| ~ p(time) ~ ψ(clear_cut_250 + mean_age_750) | 5 | 180.22 | 3.65 | 0.08 | −84.94 |
| ~ p(time) ~ ψ(mean_age_250) | 4 | 182.46 | 5.89 | 0.03 | −87.12 |
| ~ p(time) ~ ψ(old_spruce_250 + sd_age_750) | 5 | 182.76 | 6.20 | 0.02 | −86.21 |
| ~ p(time) ~ ψ(sd_age_750) | 4 | 183.63 | 7.06 | 0.01 | −87.70 |
| ~ p(time) ~ ψ(mean_age_250 + old_spruce_750) | 5 | 183.86 | 7.30 | 0.01 | −86.76 |
| ~ p(time) ~ ψ(null) | 3 | 185.05 | 8.49 | 0.01 | −89.46 |
| ~ p(time) ~ ψ(old_spruce_250) | 4 | 185.25 | 8.68 | 0.01 | −88.51 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lamarre, V.; Tremblay, J.A. Occupancy of the American Three-Toed Woodpecker in a Heavily-Managed Boreal Forest of Eastern Canada. Diversity 2021, 13, 35. https://doi.org/10.3390/d13010035
Lamarre V, Tremblay JA. Occupancy of the American Three-Toed Woodpecker in a Heavily-Managed Boreal Forest of Eastern Canada. Diversity. 2021; 13(1):35. https://doi.org/10.3390/d13010035
Chicago/Turabian StyleLamarre, Vincent, and Junior A. Tremblay. 2021. "Occupancy of the American Three-Toed Woodpecker in a Heavily-Managed Boreal Forest of Eastern Canada" Diversity 13, no. 1: 35. https://doi.org/10.3390/d13010035
APA StyleLamarre, V., & Tremblay, J. A. (2021). Occupancy of the American Three-Toed Woodpecker in a Heavily-Managed Boreal Forest of Eastern Canada. Diversity, 13(1), 35. https://doi.org/10.3390/d13010035